
Multi-Omics Unveils Inflammatory Regulation of Fermented Sini Decoction Dregs in Broilers Infected with Avian Pathogenic Escherichia coli

 ,
,
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Transcriptome Sequencing Datasets
2.2. Differential Expression Genes (DEGs) Analysis
2.3. Weighted Gene Co-Expression Network Analysis (WGCNA)
2.4. Protein–Protein Interaction (PPI) Network
2.5. CIBERSORT Immune Infiltration Analysis
2.6. Gene Enrichment Analysis
2.7. Preparation of Sini Decoction Dregs Fermentation Liquid
2.8. Metabolites Extraction
2.9. LC–MS/MS Analysis
2.10. Molecular Docking
2.11. Construction of Cell Inflammation Model
2.12. Construction of Broiler Prevention Model
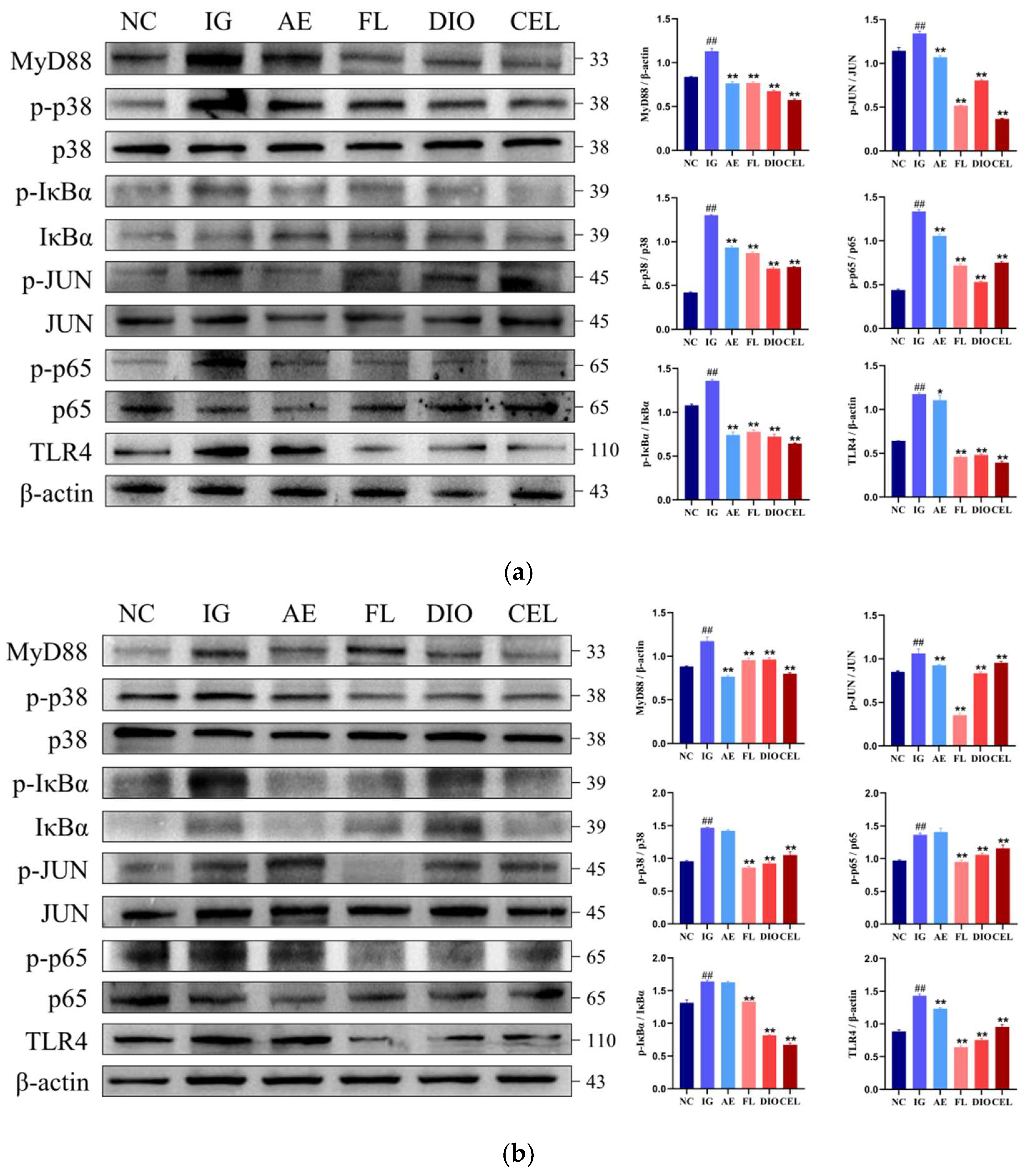
2.13. Total Protein Extraction and Western Blot
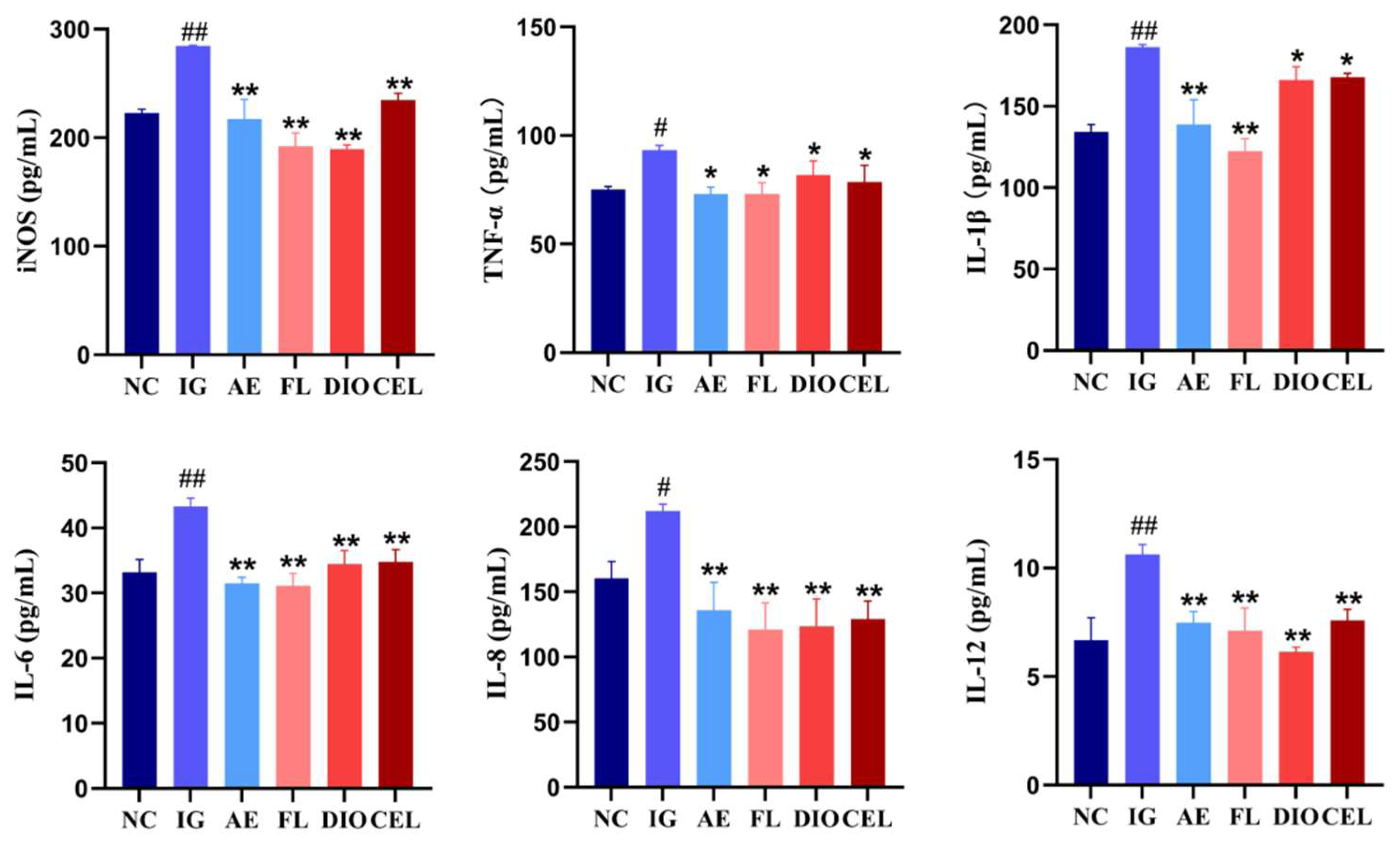
2.14. Enzyme-Linked Immunosorbent Assay (ELISA)
3. Results
3.1. Transcriptome Analysis Results
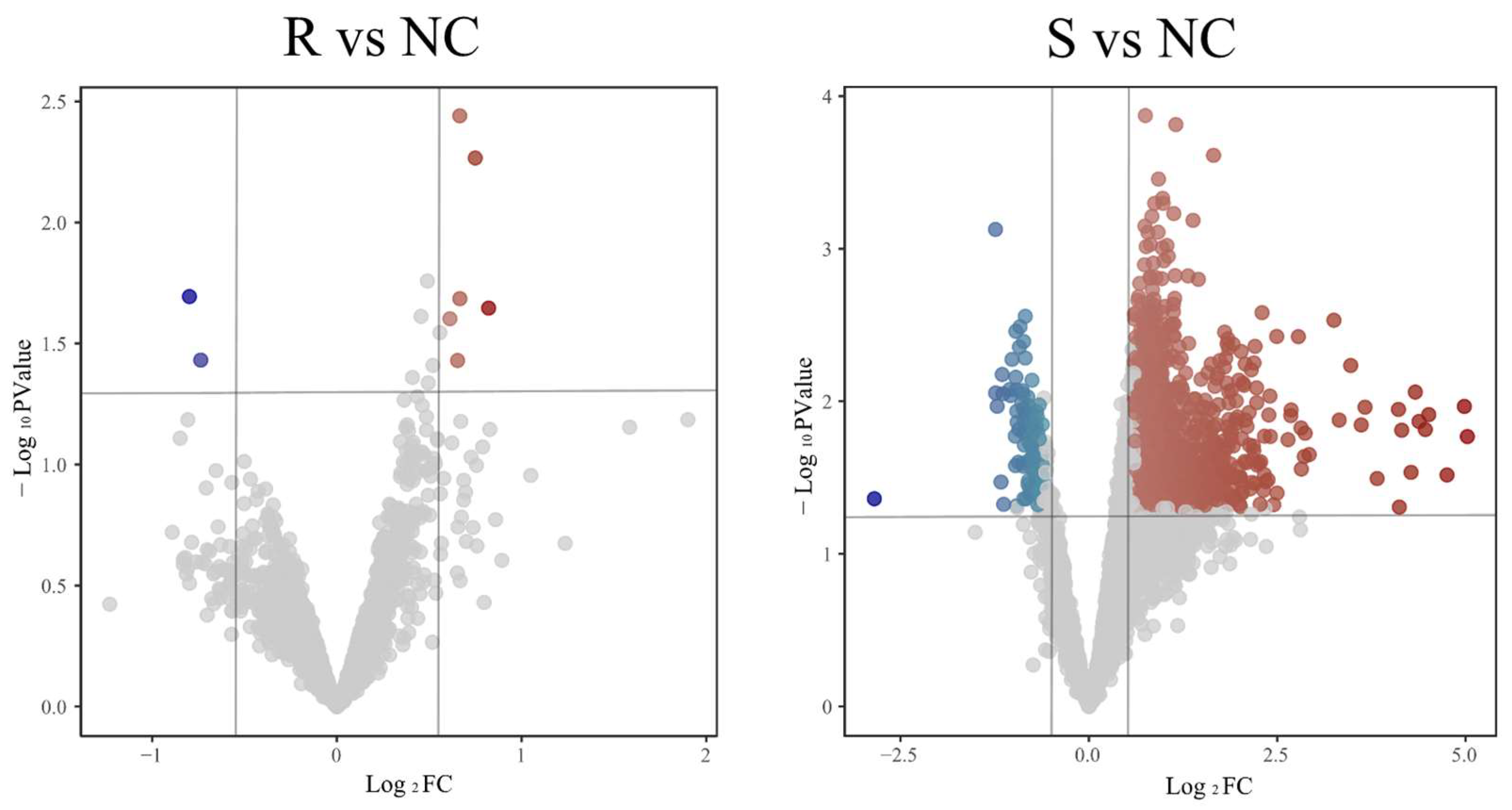
3.1.1. Identification of DEGs
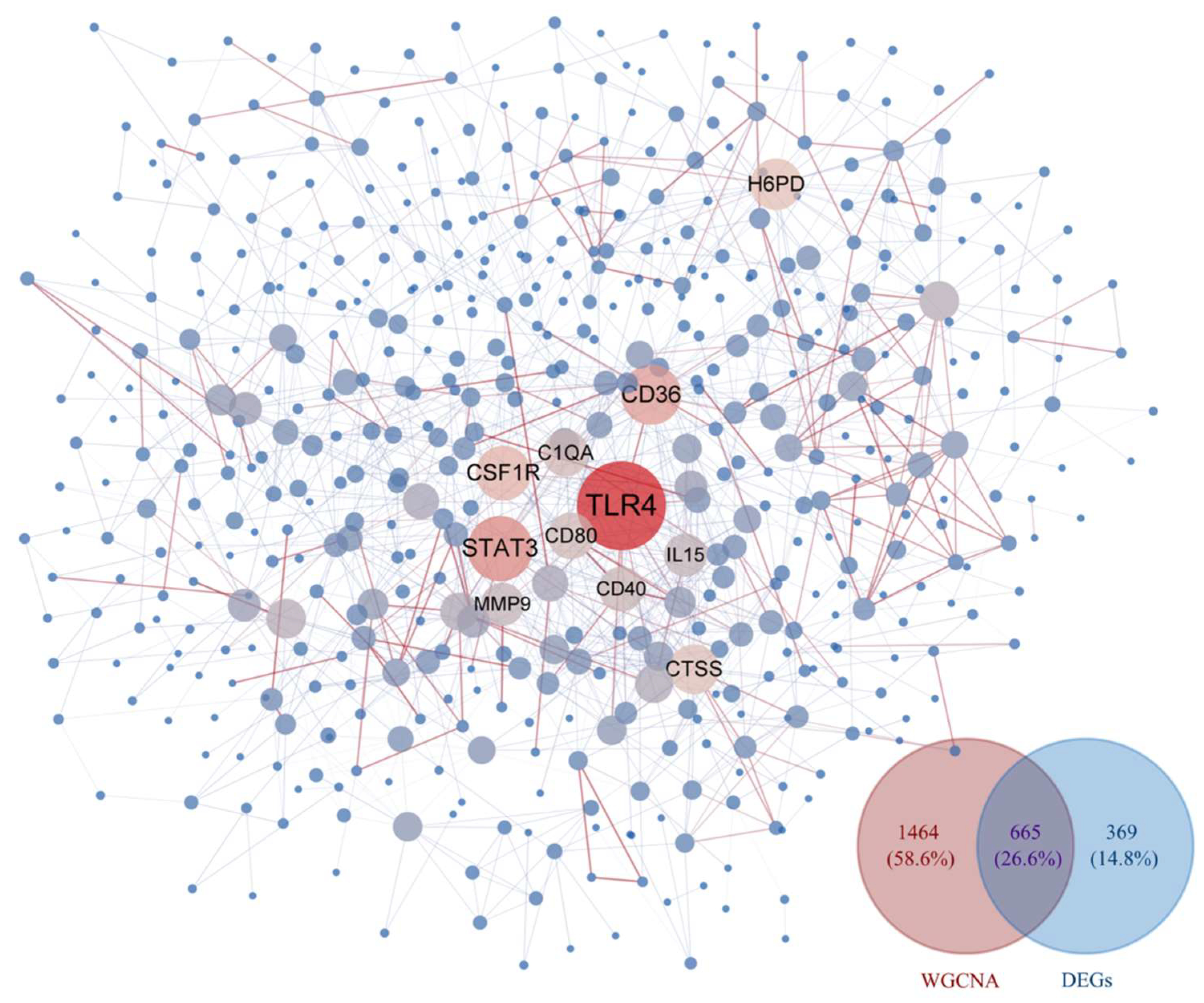
3.1.2. WGCNA and Identification of Key Modules
3.1.3. Construction of PPI Network
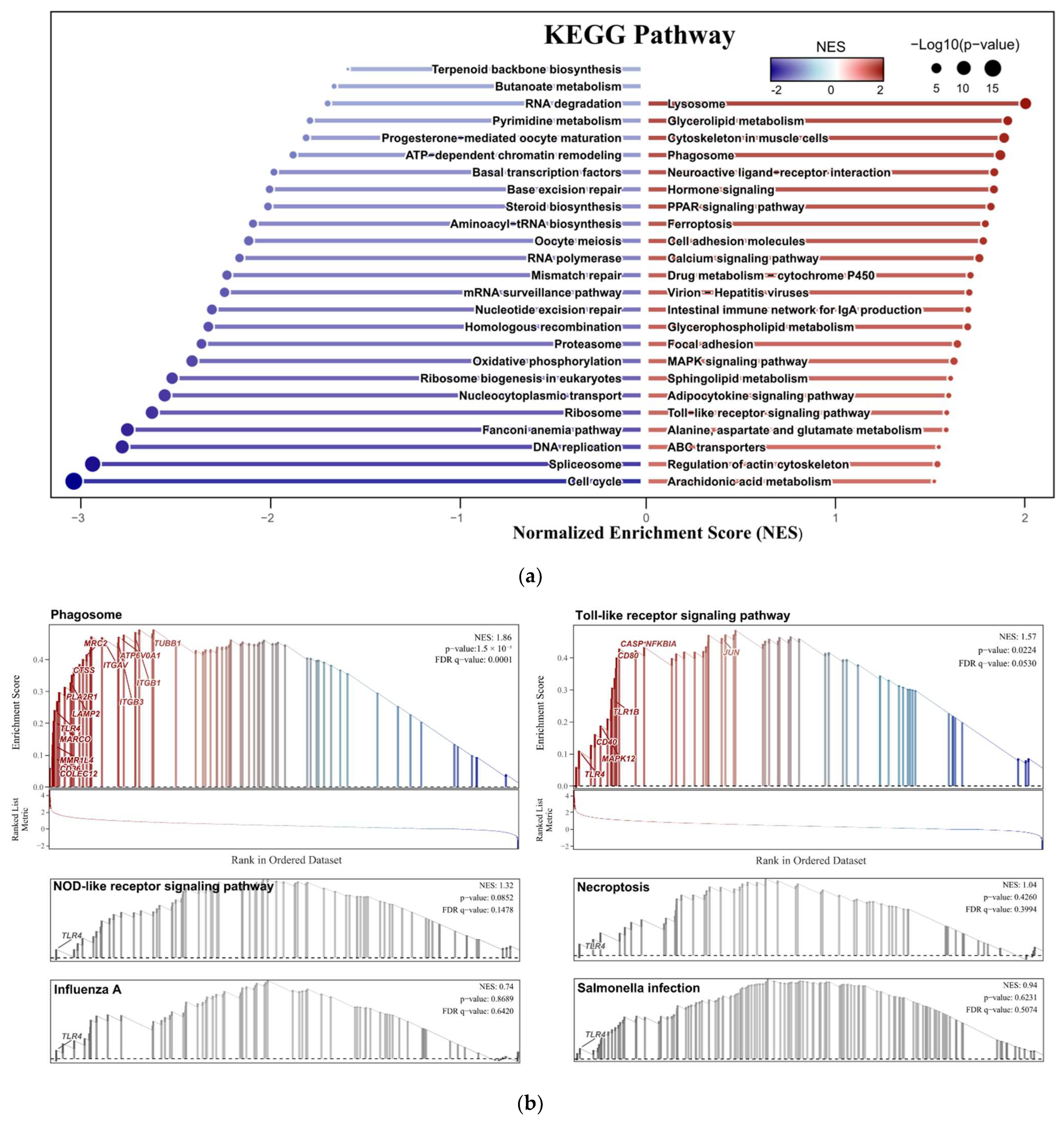
3.1.4. GSEA Results
3.1.5. CIBERSORT Immune Infiltration Results
3.1.6. Components of FLSDD
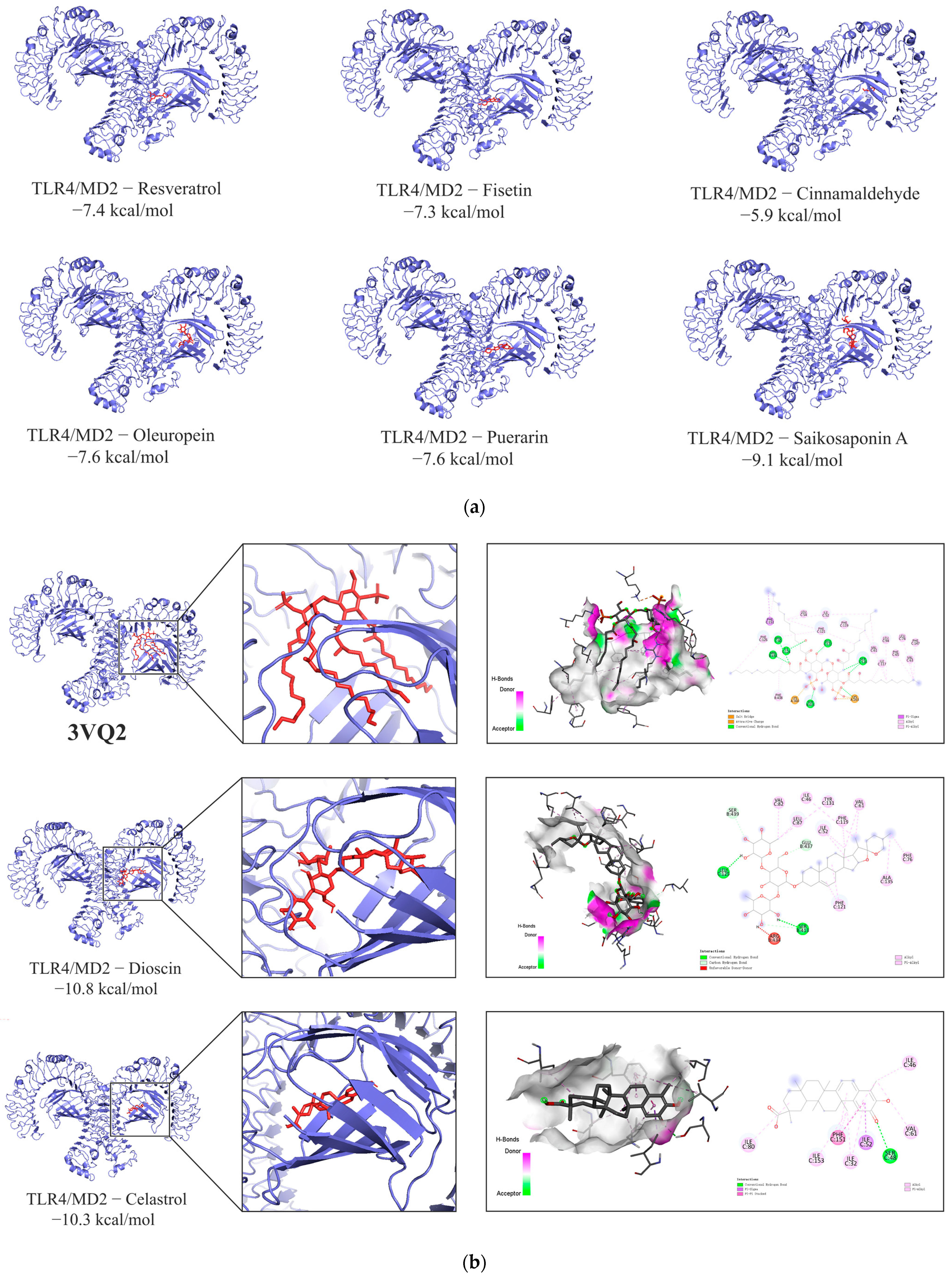
3.1.7. Molecular Docking Results
3.2. Cell and Animal Experiments Results
3.2.1. Western Blot
3.2.2. ELISA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| BCA | Bicinchoninic Acid |
| ECL | Enhanced Chemiluminescence Luminescent Solution |
| PBS | Phosphate-Buffered Saline |
| PMSF | Phenylmethylsulfonyl Fluoride |
| PVDF | Polyvinylidene Difluoride |
| RIPA | Radioimmunoprecipitation Assay |
| SDS-PAGE | Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis |
| TBST | Tris-Buffered Saline with Tween |
References
- Zhao, M.; Chen, Y.; Wang, C.; Xiao, W.; Chen, S.; Zhang, S.; Yang, L.; Li, Y. Systems pharmacology dissection of multi-scale mechanisms of action of Huo-Xiang-Zheng-Qi formula for the treatment of gastrointestinal diseases. Front. Pharmacol. 2019, 9, 1448. [Google Scholar] [CrossRef]
- Yao, Y.; Zhang, X.; Wang, Z.; Zheng, C.; Li, P.; Huang, C.; Tao, W.; Xiao, W.; Wang, Y.; Huang, L. Deciphering the combination principles of Traditional Chinese Medicine from a systems pharmacology perspective based on Ma-huang Decoction. J. Ethnopharmacol. 2013, 150, 619–638. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Ding, C.; Liu, X.; Zhao, Y.; Ding, Q.; Sun, S.; Zhang, J.; Yang, J.; Liu, W.; Li, W. Research progress on extraction, isolation, structural analysis and biological activity of polysaccharides from panax genus. Molecules 2023, 28, 3733. [Google Scholar] [CrossRef]
- Yuan, D.; Cui, Y.; Kou, Y.; Guo, X.; Chen, B. Environmental risk and management of herbal-extraction residues induced by the composition and metal binding properties of DOM. Front. Environ. Sci. 2022, 10, 856308. [Google Scholar] [CrossRef]
- Tao, W.; Jin, J.; Zheng, Y.; Li, S. Current advances of resource utilization of herbal extraction residues in China. Waste Biomass Valorization 2021, 12, 5853–5868. [Google Scholar] [CrossRef]
- Cui, Y.; Li, J.; Deng, D.; Lu, H.; Tian, Z.; Liu, Z.; Ma, X. Solid-state fermentation by Aspergillus niger and Trichoderma koningii improves the quality of tea dregs for use as feed additives. PLoS ONE 2021, 16, e0260045. [Google Scholar] [CrossRef]
- Apostolakos, I.; Laconi, A.; Mughini-Gras, L.; Yapicier, Ö.Ş.; Piccirillo, A. Occurrence of colibacillosis in broilers and its relationship with avian pathogenic Escherichia coli (APEC) population structure and molecular characteristics. Front. Vet. Sci. 2021, 8, 737720. [Google Scholar] [CrossRef] [PubMed]
- Feng, A.; Akter, S.; Leigh, S.A.; Wang, H.; Pharr, G.T.; Evans, J.; Branton, S.L.; Landinez, M.P.; Pace, L.; Wan, X.-F. Genomic diversity, pathogenicity and antimicrobial resistance of Escherichia coli isolated from poultry in the southern United States. BMC Microbiol. 2023, 23, 15. [Google Scholar] [CrossRef]
- Ibrahim, R.A.; Cryer, T.L.; Lafi, S.Q.; Basha, E.-A.; Good, L.; Tarazi, Y.H. Identification of Escherichia coli from broiler chickens in Jordan, their antimicrobial resistance, gene characterization and the associated risk factors. BMC Vet. Res. 2019, 15, 159. [Google Scholar] [CrossRef]
- Geng, T.; He, F.; Su, S.; Sun, K.; Zhao, L.; Zhao, Y.; Bao, N.; Pan, L.; Sun, H. Probiotics Lactobacillus rhamnosus GG ATCC53103 and Lactobacillus plantarum JL01 induce cytokine alterations by the production of TCDA, DHA, and succinic and palmitic acids, and enhance immunity of weaned piglets. Res. Vet. Sci. 2021, 137, 56–67. [Google Scholar] [CrossRef]
- Weill, F.S.; Cela, E.M.; Paz, M.L.; Ferrari, A.; Leoni, J.; Maglio, D.H.G. Lipoteichoic acid from Lactobacillus rhamnosus GG as an oral photoprotective agent against UV-induced carcinogenesis. Br. J. Nutr. 2013, 109, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.; Kyoung, H.; Park, K.I.; Kim, Y.; Lee, J.I.; Ha, S.; Nam, J.; Kim, K.; Kang, Y.; Park, Y. PSXIII-7 Effect of Dietary Supplementation of Lactobacillus Rhamnosus on Blood Profiles, Intestinal Morphology, Immune Responses, and Gene Expression of Tight Junction Proteins and Cytokines of Weaned Pigs. J. Anim. Sci. 2023, 101 (Suppl. S3), 574–575. [Google Scholar] [CrossRef]
- Sun, H.; Liu, P.; Nolan, L.K.; Lamont, S.J. Thymus transcriptome reveals novel pathways in response to avian pathogenic Escherichia coli infection. Poult. Sci. 2016, 95, 2803–2814. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, X.; Lv, N.; Wang, L.; Gan, J.; Jiang, X.; Wang, Y. Therapeutic role and potential mechanism of resveratrol in atherosclerosis: TLR4/NF-κB/HIF-1α. Mediat. Inflamm. 2023, 2023, 1097706. [Google Scholar] [CrossRef]
- Okan, A.; Doğanyiğit, Z.; Yilmaz, S.; Uçar, S.; Arikan Söylemez, E.S.; Attar, R. Evaluation of the protective role of resveratrol against sepsis caused by LPS via TLR4/NF-κB/TNF-α signaling pathways: Experimental study. Cell Biochem. Funct. 2023, 41, 423–433. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Huang, L.; Sun, K.; Li, J.; Han, S.; Gao, X.; Wang, Q.-Q.; Yang, S.; Sun, W.; Gao, H. Oleuropein alleviates myocardial ischemia–reperfusion injury by suppressing oxidative stress and excessive autophagy via TLR4/MAPK signaling pathway. Chin. Med. 2024, 19, 59. [Google Scholar] [CrossRef]
- Yakut, S.; Gelen, V.; Kara, H.; Özkanlar, S.; Yeşildağ, A. Silver Nanoparticles Loaded With Oleuropein Alleviates LPS-Induced Acute Lung Injury by Modulating the TLR4/P2X7 Receptor-Mediated Inflammation and Apoptosis in Rats. Environ. Toxicol. 2024, 39, 4960–4973. [Google Scholar] [CrossRef]
- Chen, P.; Zhou, J.; Ruan, A.; Zeng, L.; Liu, J.; Wang, Q. Cinnamic Aldehyde, the main monomer component of Cinnamon, exhibits anti-inflammatory property in OA synovial fibroblasts via TLR4/MyD88 pathway. J. Cell. Mol. Med. 2022, 26, 913–924. [Google Scholar] [CrossRef]
- Han, J.; Shi, G.; Li, W.; Xie, Y.; Li, F.; Jiang, D. Preventive effect of Dioscin against monosodium urate-mediated gouty arthritis through inhibiting inflammasome NLRP3 and TLR4/NF-κB signaling pathway activation: An in vivo and in vitro study. J. Nat. Med. 2021, 75, 37–47. [Google Scholar] [CrossRef]
- Jin, S.; Guan, T.; Wang, S.; Hu, M.; Liu, X.; Huang, S.; Liu, Y. Dioscin alleviates cisplatin-induced mucositis in rats by modulating gut microbiota, enhancing intestinal barrier function and attenuating TLR4/NF-κB signaling cascade. Int. J. Mol. Sci. 2022, 23, 4431. [Google Scholar] [CrossRef]
- Huang, X.; Shen, H.; Liu, Y.; Qiu, S.; Guo, Y. Fisetin attenuates periodontitis through FGFR1/TLR4/NLRP3 inflammasome pathway. Int. Immunopharmacol. 2021, 95, 107505. [Google Scholar] [CrossRef] [PubMed]
- Jian, J.; Wang, D.; Xiong, Y.; Wang, J.; Zheng, Q.; Jiang, Z.; Zhong, J.; Yang, S.; Wang, L. Puerarin alleviated oxidative stress and ferroptosis during renal fibrosis induced by ischemia/reperfusion injury via TLR4/Nox4 pathway in rats. Acta Cir. Bras. 2023, 38, e382523. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-L.; Fan, L.; Yue, B.-H.; Lou, Z. Saikosaponin A mitigates the progression of Parkinson’s disease via attenuating microglial neuroinflammation through TLR4/MyD88/NF-κB pathway. Eur. Rev. Med. Pharmacol. Sci. 2023, 27, 6956–6971. [Google Scholar]
- Jin, G.-j.; Peng, X.; Chen, Z.-G.; Wang, Y.-l.; Liao, W.-J. Celastrol attenuates chronic constrictive injury-induced neuropathic pain and inhibits the TLR4/NF-κB signaling pathway in the spinal cord. J. Nat. Med. 2022, 76, 268–275. [Google Scholar] [CrossRef]
- Gerasimova, A.; Novikova, O.; Savicheva, A. Avian colibacillosis—Current aspects. Vet. Sci. Today 2023, 2023, 284. [Google Scholar] [CrossRef]
- Ahmed, A.A.; Salem, H.M.; Hamoud, M.M.; Amer, M.M. Avian Colibacillosis, Multidrug Resistance, Antibiotic Alternatives: An Updated Review. Egypt. J. Vet. Sci. 2025, 56, 1–21. [Google Scholar] [CrossRef]
- Bhattarai, R.K.; Basnet, H.B.; Dhakal, I.P.; Devkota, B. Antimicrobial resistance of avian pathogenic Escherichia coli isolated from broiler, layer, and breeder chickens. Vet. World 2024, 17, 480. [Google Scholar] [CrossRef]
- Lu, Q.; Li, C. Comprehensive utilization of Chinese medicine residues for industry and environment protection: Turning waste into treasure. J. Clean. Prod. 2021, 279, 123856. [Google Scholar] [CrossRef]
- Yang, H.-Y.; Han, L.; Lin, Y.-Q.; Li, T.; Wei, Y.; Zhao, L.-H.; Tong, X.-L. Probiotic fermentation of herbal medicine: Progress, challenges, and opportunities. Am. J. Chin. Med. 2023, 51, 1105–1126. [Google Scholar] [CrossRef]
- Hao, S.; Shi, W.; Chen, L.; Kong, T.; Wang, B.; Chen, S.; Guo, X. CATH-2-derived antimicrobial peptide inhibits multidrug-resistant Escherichia coli infection in chickens. Front. Cell. Infect. Microbiol. 2024, 14, 1390934. [Google Scholar] [CrossRef]
- Gauthier, A.E.; Rotjan, R.D.; Kagan, J.C. Lipopolysaccharide detection by the innate immune system may be an uncommon defence strategy used in nature. Open Biol. 2022, 12, 220146. [Google Scholar] [CrossRef]
- Baranova, I.N.; Bocharov, A.V.; Vishnyakova, T.G.; Chen, Z.; Ke, Y.; Birukova, A.A.; Yuen, P.S.; Tsuji, T.; Star, R.A.; Birukov, K.G. Class B Scavenger Receptor CD36 as a Potential Therapeutic Target in Inflammation Induced by Danger-Associated Molecular Patterns. Cells 2024, 13, 1992. [Google Scholar] [CrossRef]
- Chen, G.-Y.; Xu, X.-D.; Feng, J.-X.; Zhang, J.; Li, X.-P. A single C1q A chain promotes leukocytes phagocytosis/chemotaxis and mediates bacterial clearance in Japanese flounder Paralichthys olivaceus. Aquaculture 2024, 587, 740840. [Google Scholar] [CrossRef]
- Smirnova, T.; Spertini, C.; Spertini, O. CSF1R inhibition combined with GM-CSF reprograms macrophages and disrupts protumoral interplays with AML cells. Cancers 2021, 13, 5289. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X.-Y. Transcription factors associated with IL-15 cytokine signaling during NK cell development. Front. Immunol. 2021, 12, 610789. [Google Scholar] [CrossRef] [PubMed]
- Mazzoccoli, L.; Liu, B. Dendritic cells in shaping anti-tumor T cell response. Cancers 2024, 16, 2211. [Google Scholar] [CrossRef]
- Zeng, Y.; Gao, M.; Lin, D.; Du, G.; Cai, Y. Prognostic and immunological roles of MMP-9 in Pan-cancer. BioMed Res. Int. 2022, 2022, 2592962. [Google Scholar] [CrossRef]
- Zhao, K.; Sun, Y.; Zhong, S.; Luo, J.-L. The multifaceted roles of cathepsins in immune and inflammatory responses: Implications for cancer therapy, autoimmune diseases, and infectious diseases. Biomark. Res. 2024, 12, 165. [Google Scholar] [CrossRef]
- De Angelis, M.; Amatore, D.; Checconi, P.; Zevini, A.; Fraternale, A.; Magnani, M.; Hiscott, J.; De Chiara, G.; Palamara, A.T.; Nencioni, L. Influenza virus down-modulates G6PD expression and activity to induce oxidative stress and promote its replication. Front. Cell. Infect. Microbiol. 2022, 11, 804976. [Google Scholar] [CrossRef]
- Li, D.; Wu, M. Pattern recognition receptors in health and diseases. Signal Transduct. Target. Ther. 2021, 6, 291. [Google Scholar]
- Wicherska-Pawłowska, K.; Wróbel, T.; Rybka, J. Toll-like receptors (TLRs), NOD-like receptors (NLRs), and RIG-I-like receptors (RLRs) in innate immunity. TLRs, NLRs, and RLRs ligands as immunotherapeutic agents for hematopoietic diseases. Int. J. Mol. Sci. 2021, 22, 13397. [Google Scholar] [CrossRef]
- Li, X.; Xu, M.; Shen, J.; Li, Y.; Lin, S.; Zhu, M.; Pang, Q.; Tan, X.; Tang, J. Sorafenib inhibits LPS-induced inflammation by regulating Lyn-MAPK-NF-kB/AP-1 pathway and TLR4 expression. Cell Death Discov. 2022, 8, 281. [Google Scholar] [CrossRef]
- Sawoo, R.; Dey, R.; Ghosh, R.; Bishayi, B. TLR4 and TNFR1 blockade dampen M1 macrophage activation and shifts them towards an M2 phenotype. Immunol. Res. 2021, 69, 334–351. [Google Scholar] [CrossRef]
- Yan, L.; Wang, J.; Cai, X.; Liou, Y.C.; Shen, H.M.; Hao, J.; Huang, C.; Luo, G.; He, W. Macrophage plasticity: Signaling pathways, tissue repair, and regeneration. MedComm 2024, 5, e658. [Google Scholar] [CrossRef]
- Oblak, A.; Jerala, R. The molecular mechanism of species-specific recognition of lipopolysaccharides by the MD-2/TLR4 receptor complex. Mol. Immunol. 2015, 63, 134–142. [Google Scholar] [CrossRef]
- Hopp, M.T.; Holze, J.; Lauber, F.; Holtkamp, L.; Rathod, D.C.; Miteva, M.A.; Prestes, E.B.; Geyer, M.; Manoury, B.; Merle, N.S. Insights into the molecular basis and mechanism of heme-triggered TLR4 signalling: The role of heme-binding motifs in TLR4 and MD2. Immunology 2024, 171, 181–197. [Google Scholar] [CrossRef] [PubMed]
- Bandopadhyay, S.; Anand, U.; Gadekar, V.S.; Jha, N.K.; Gupta, P.K.; Behl, T.; Kumar, M.; Radha; Shekhawat, M.S.; Dey, A. Dioscin: A review on pharmacological properties and therapeutic values. BioFactors 2022, 48, 22–55. [Google Scholar] [PubMed]
- Wang, C.; Dai, S.; Zhao, X.; Zhang, Y.; Gong, L.; Fu, K.; Ma, C.; Peng, C.; Li, Y. Celastrol as an emerging anticancer agent: Current status, challenges and therapeutic strategies. Biomed. Pharmacother. 2023, 163, 114882. [Google Scholar] [CrossRef]
- Opo, F.A.D.M.; Rahman, M.M.; Ahammad, F.; Ahmed, I.; Bhuiyan, M.A.; Asiri, A.M. Structure based pharmacophore modeling, virtual screening, molecular docking and ADMET approaches for identification of natural anti-cancer agents targeting XIAP protein. Sci. Rep. 2021, 11, 4049. [Google Scholar]
- Bolmanis, E.; Dubencovs, K.; Suleiko, A.; Vanags, J. Model predictive control—A stand out among competitors for fed-batch fermentation improvement. Fermentation 2023, 9, 206. [Google Scholar] [CrossRef]
- Yao, Z.; Yu, J.; Tang, Z.; Liu, H.; Ruan, K.; Song, Z.; Liu, Y.; Yan, K.; Liu, Y.; Tang, Y. Multi-evaluating strategy for Siji-kangbingdu mixture: Chemical profiling, fingerprint characterization, and quantitative analysis. Molecules 2019, 24, 3545. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, P.K.; Banerjee, S.; Kar, A. Molecular combination networks in medicinal plants: Understanding synergy by network pharmacology in Indian traditional medicine. Phytochem. Rev. 2021, 20, 693–703. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ID | Group | Concentration | Challenge Dose |
|---|---|---|---|
| 1 | NC | — | — |
| 2 | AE | 50 mg/mL | 1 × 105/well |
| 3 | FLSDD | 50 mg/mL | 1 × 105/well |
| 4 | CEL | 1 μg/mL | 1 × 105/well |
| 5 | DIO | 150 μg/mL | 1 × 105/well |
| 7 | MG | — | 1 × 105/well |
| ID | Group | Treat | Dosage | Challenge Dose |
|---|---|---|---|---|
| 1 | NC | — | — | — |
| 2 | AE | Oral | 100 mg/mL, 0.5 mL/day/bird | 1 × 109 CFU |
| 3 | FLSDD | Oral | 100 mg/mL, 0.5 mL/day/bird | 1 × 109 CFU |
| 4 | CEL | Oral | 0.1 mg/kg/day/bird | 1 × 109 CFU |
| 5 | DIO | Oral | 10 mg/kg/day/bird | 1 × 109 CFU |
| 7 | MG | — | — | 1 × 109 CFU |
| ID | Compounds Name | CAS | Structures | References |
|---|---|---|---|---|
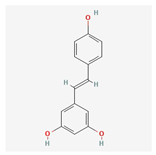 | ||||
| 1 | Resveratrol | 501-36-0 | [14,15] | |
 | ||||
| 2 | Oleuropein | 32619-42-4 | [16,17] | |
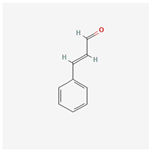 | ||||
| 3 | Cinnamaldehyde | 104-55-2 | [18] | |
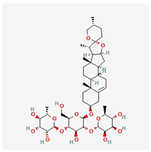 | ||||
| 4 | Dioscin | 19057-60-4 | [19,20] | |
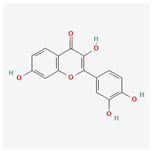 | ||||
| 5 | Fisetin | 528-48-3 | [21] | |
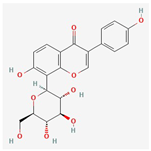 | ||||
| 6 | Puerarin | 3681-99-0 | [22] | |
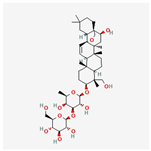 | ||||
| 7 | Saikosaponin A | 20736-09-8 | [23] | |
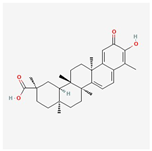 | ||||
| 8 | Celastrol | 34157-83-0 | [24] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mo, S.; Fang, X.; Xiao, W.; Huang, B.; Li, C.; Yang, H.; Wu, Y.; Wang, Y.; Ma, H. Multi-Omics Unveils Inflammatory Regulation of Fermented Sini Decoction Dregs in Broilers Infected with Avian Pathogenic Escherichia coli. Vet. Sci. 2025, 12, 479. https://doi.org/10.3390/vetsci12050479
Mo S, Fang X, Xiao W, Huang B, Li C, Yang H, Wu Y, Wang Y, Ma H. Multi-Omics Unveils Inflammatory Regulation of Fermented Sini Decoction Dregs in Broilers Infected with Avian Pathogenic Escherichia coli. Veterinary Sciences. 2025; 12(5):479. https://doi.org/10.3390/vetsci12050479
Chicago/Turabian StyleMo, Shuanghao, Xin Fang, Wenxi Xiao, Bowen Huang, Chunsheng Li, Hui Yang, Yilin Wu, Yiming Wang, and Hongxia Ma. 2025. "Multi-Omics Unveils Inflammatory Regulation of Fermented Sini Decoction Dregs in Broilers Infected with Avian Pathogenic Escherichia coli" Veterinary Sciences 12, no. 5: 479. https://doi.org/10.3390/vetsci12050479
APA StyleMo, S., Fang, X., Xiao, W., Huang, B., Li, C., Yang, H., Wu, Y., Wang, Y., & Ma, H. (2025). Multi-Omics Unveils Inflammatory Regulation of Fermented Sini Decoction Dregs in Broilers Infected with Avian Pathogenic Escherichia coli. Veterinary Sciences, 12(5), 479. https://doi.org/10.3390/vetsci12050479