1. Introduction
Fats and oils are supplemented in dog diets to increase dietary energy levels in order to meet daily nutrient requirements and palatability. Fats are considered an essential nutrient for dogs [
1]. The rise in the number of pets globally has contributed to substantial growth in this sector, with the global pet food industry estimated to be worth approximately USD 115 billion. Dog and cat food account for approximately 70% of this value [
2]. It has been reported that approximately 290,000 tonnes of animal fat is used each year to produce pet food in the USA, some of which is of human food grade [
3]. With more than 155 million people in 55 countries facing a food crisis worldwide, competition for using ingredients in pet and human food significantly impacts global food security. Therefore, it is necessary to explore the potential of new sources of fats that are not currently used in human food [
4].
The black soldier fly (
Hermetia illucens) is one of the insect species most intensively studied for use in animal nutrition [
5]. Major pet food brands worldwide have introduced formulas containing BSF protein, but there is still little interest in BSF lipids (fat). Recently, two European brands marketed eight pet foods containing BSF fat [
4]. When producing partially or fully defatted larvae meal from black soldier fly larvae (BSFL), a significant amount (30–40%) of fat, a nutritionally valuable by-product, is obtained [
6]. The fat content and fatty acid composition of BSFL are influenced by the feed used for their growth and the process of extracting the fat from larvae [
7,
8,
9]. Approximately 90% of BSFL fat comprises six fatty acids: lauric; palmitic; oleic; linoleic; and, to a lesser extent, myristic and stearic acids. The most abundant is lauric acid, representing about a third of the total fatty acids [
10].
BSFL fat not only fulfils the metabolic requirements of animals but also supports animal health. Lauric acid has a proven antimicrobial effect [
11,
12] and is also associated with many health benefits, such as the prevention of cardiovascular disease, antiviral properties, cancer prevention, and reducing the risk of Alzheimer’s disease and obesity [
13]. Wang et al. [
14] have shown that medium-chain fatty acids may be beneficial in reducing abdominal obesity compared to long-chain saturated or unsaturated fatty acids. Lauric acid offers specific benefits for the cognitive function of dogs: Chronic inflammation can negatively impact brain health in dogs, contributing to cognitive decline. Lauric acid exhibits anti-inflammatory properties, which may help protect neurons and support overall brain health. BSFL contains important levels of proteins and fats to nourish cats and dogs. BSFL also contains bioactive components, such as lauric acid, chitin, and antimicrobial peptides, known for their potential to improve animal health [
15].
Insect fat, with a high lauric acid content, has been shown to positively affect digestive health in fast-growing broiler chickens [
16,
17]. Due to its lauric acid content, BSFL can regulate blood cholesterol levels [
18]. In broiler chickens, BSFL fat significantly reduces serum total cholesterol and HDL compared to coconut oil [
19]. Sypniewski et al. [
20] reported that the serum total cholesterol, HDL, and LDL of young turkeys consuming BSF fat instead of soybean oil are significantly reduced. Furthermore, BSFL fat has been reported to positively influence plasma immune and antioxidant activity, as well as improve gut morphology and barrier integrity in broiler chickens [
21].
The allergenic potential is the potential of a food or ingredient to cause sensitisation and allergic reactions, often in association with an IgE antibody. There is a risk of allergy to edible insects due to different rearing environments or acquisition methods. Many food components trigger IgE and IgG antibodies in patients without skin or gastrointestinal symptoms. Protein is one of the main allergens in dogs [
22]. However, a decreased serum IgG concentration has been recorded with the addition of BSF fat in turkeys and broilers [
23,
24]. The pathogenic significance of IgG antibodies to food antigens in dogs has yet to be clarified; the conventional view is that their detection reflects only previous exposure and tolerance and not specific food-related pathogenesis.
The majority of research on insect fats has focused on consumption by poultry. Studies have shown no significant differences in nutrient digestibility when soybean oil was fully replaced with BSF fat in broiler diets [
7,
25]. Nevertheless, including BSF fat has been reported to enhance ileal crude protein digestibility and ileal digestible energy [
23]. In most studies investigating substituting insect fat in broiler, laying hen, and turkey diets, no significant impact on crude protein and crude fat digestibility has been observed [
24]. Similarly, diets containing BSFL fat do not affect nutrient digestibility, faecal quality, or blood parameters in dogs [
26]. When BSFL fat was used to replace 8% of plant oil in the diets of healthy dogs, serum biochemical parameters remained within normal ranges, with no observed changes in body weight, nutrient digestibility, or short-chain fatty acid (SCFA) concentrations in faeces [
27].
Over the past two decades, advances in human and companion animal immunology have facilitated a deeper understanding of the relationship between host nutrition, immunomodulation, and their impact on overall health. Given the adverse effects associated with high dietary levels of saturated fatty acids, such as palmitic and stearic acids, it is crucial to establish the optimal inclusion levels of insects as a dietary source of fatty acids. BSF is a highly investigated insect due to its strong fecundity, high conversion rate, high nutrition, low cost, and easy management, and it is most commonly used in pet food [
28]. Despite the recognised nutritional potential of BSFL fat, there remains a lack of comprehensive scientific data regarding its effects on blood chemistry, immune function, nutrient digestibility, and the palatability of the food to dogs. Consequently, this study evaluates the impact of replacing half or all of the poultry fat in extruded dog food with BSFL fat on nutrient digestibility, faecal metabolites, faecal quality, selected biochemical and immunological blood parameters, food preference by dogs, and oxidative stability of the food.
3. Results
The dry matter, organic matter, crude protein, and fat digestibility levels determined by the indicator method are given in
Table 3. Dry matter and organic matter digestibility percentages were lowest in BSF6 (
p < 0.001). The crude protein digestibility of BSF6 was lower than the control (
p < 0.001). The highest digestibility percentage of the ether extraction was determined in the control food (
p = 0.002).
Biochemical parameters measured in blood samples collected from all animals on the final day (30th day) of the digestibility trial, before the morning feeding, are presented in
Table 4, and blood IgE and IgG levels are shown in
Table 5. No significant differences were observed in serum glucose, triglyceride, cholesterol, blood urea nitrogen (BUN), total protein, creatinine, aspartate aminotransferase (AST), or alanine aminotransferase (ALT) levels between the groups. Similarly, including poultry fat or BSFL fat in the diets had no significant effect on serum IgE or IgG concentrations (
p > 0.05).
The faecal scores of the faeces of all animals on the last five days of the digestion trial were visually scored by three researchers. The pH, dry matter, ammonia nitrogen, and SCFA of fresh faecal samples collected on the last three days of the digestion trial are given in
Table 6. The faecal acetic acid level was higher in the control-group dogs (
p = 0.005). Although the amount of isovaleric acid was lower in the BSF3-group dogs, the lowest valeric acid levels were measured in control-group dog faeces (
p = 0.037).
The preference test results are presented in
Table 7. The BSF6 food was preferred at the rate of 55.13%. This percentage was significantly higher than the preference rate of the BSF6 food (44.88%) (
p = 0.035).
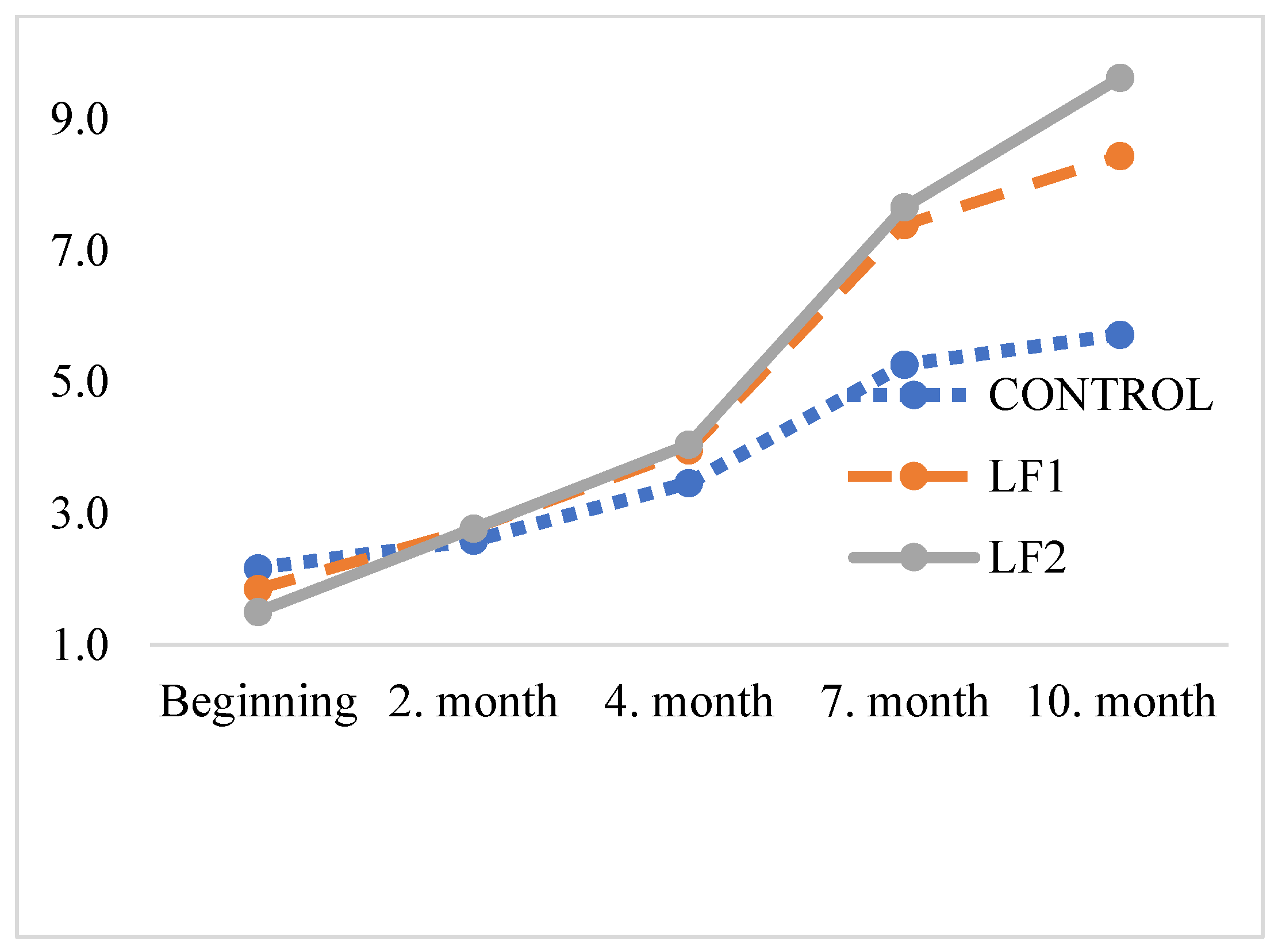
The results of MDA levels of 0, 2, 4, 7, and 10 months after storage of the foods are presented in
Table 8, and corresponding curves are shown in
Figure 2. Similar oxidation values were observed in dog foods containing poultry fat and BSFL fat throughout the 10-month storage period.
4. Discussion
Although an equal amount of fat was added to all three foods, chemical analyses showed that the ether extract was lower in the foods where BSFL was used. In the chemical analysis of BSFL, 99.79% dry matter and 94.22% ether extract were measured. After the fat was obtained, the protein was separated along with the sticky substances in the purification process. A previous study measured 92% ether extract in BSFL [
41].
Martins et al. [
42] reported that replacing linseed oil with BSFL fat at 3% and 6% in the diets of young rabbits negatively affected the digestibility of dry matter, organic matter, and crude fat, as observed in this study. However, another study showed that replacing soybean oil with BSFL fat did not impact nutrient digestibility [
43]. This discrepancy is likely due to the lower levels of BSFL fat (1.5%) in the study by Gasco et al. [
43]. In contrast to these findings, Freel et al. [
30] reported no differences in the digestibility of dry matter and crude protein between diets containing poultry fat and those containing BSFL fat for dogs. Freel et al. [
27] also observed higher digestibility coefficients for dry matter (90–91%), crude protein (89–91%), and ether extraction (96.0–96.8%) of dry foods compared to the results of this study (74.5–81.2%, 73.9–79.9%, and 86.3–93.9%, respectively). Similarly, Jian et al. [
28] found that the inclusion of BSFL fat in dog diets reduced the dry matter digestibility (from 82.2% to 78.9%) and ether extract digestibility (from 95.5% to 93.9%), although these reductions were not statistically significant. Kierończyk et al. [
7] reported that replacing soybean oil with BSFL fat did not affect ether extract and crude protein digestibility in broiler chickens.
In contrast, another study by Kierończyk et al. [
23] demonstrated increased crude protein digestibility in broilers fed diets containing BSFL fat. These variations in digestibility results can likely be attributed to differences in the nutritional composition and fatty acid profiles of the BSFL fat, which are influenced by the environmental conditions in which the larvae were reared. The reduced nutrient digestibility in diets containing larvae fat in this study may be due to the inhibitory effect of lauric acid on gut microbiota [
44]. Insect fat, which is normally rich in medium-chain fatty acids, is expected to be absorbed more than poultry fat, which contains more long-chain fatty acids. However, the triacylglycerol composition of BSFL fat and the distribution of lauric acid in triacylglycerols may have adversely affected digestibility. In addition, it has been reported that the high melting point of saturated fatty acids, as in BSFL fat, makes them less soluble in bile, reducing the viscosity of the intestinal contents and their digestibility in the intestine [
45].
The biochemical results obtained in this study are consistent with the studies in dogs, rabbits, fish, and broilers [
23,
25,
28,
43,
46]. Because there is no standard for the diet of BSFLs and their fatty acid composition may vary considerably, BSFL fat used in different studies may affect cholesterol levels or different parameters differently in animals.
Acetate is the major SCFA in the colon and has been shown to increase cholesterol synthesis after absorption. However, propionate has been shown to inhibit cholesterol synthesis. Therefore, substrates that can reduce the acetate:propionate ratio may reduce the risk of cardiovascular disease. As long as the faecal quality is not affected, it is preferable to produce more SCFAs [
39]. Aside from its effects on health, the low intestinal acetic acid level in dogs consuming BSF fat-based food is an unfavourable condition. Acetic acid regulates the pH balance in the intestine and aids digestion. Low levels can negatively affect digestion and impair nutrient absorption [
47]. This could be another reason for the reduced nutrient digestibility of the BSF3 and BSF6 dry foods.
The decreased acetate level may be due to lauric acid in BSFL fat suppressing acetic acid-producing bacteria by changing the intestinal microbiota [
48].
Clostridiaceae, which break down indigestible complex carbohydrates to produce SCFAs, may have been negatively affected by the presence of lauric acid [
49]. Consistent with this study, Kierończyk et al. [
50] and Jian et al. [
28] also reported propionate and butyrate levels unaffected by BSFL fat administration. Valeric acid was reported to increase in the faecal content of broiler chickens fed larvae fat [
51]. A previous study highlighted numerous protective and beneficial effects associated with the increase in or administration of valeric acid, including its potential role in mitigating allergies, intestinal dysbiosis, colitis, enteritis, experimental encephalitis, and eczema [
49].
Ammonia in faeces is a putrefactive by-product generated through the fermentation of undigested protein. In the present study, faecal ammonia concentrations increased for dogs fed diets containing BSFL fat as a replacement for whole-poultry oil, which aligns with the reduced protein digestibility of these diets. Faecal pH is strongly correlated with SCFAs and is a reliable indicator of SCFA production. Within the intestine, microbial fermentation of substrates produces SCFAs, lowering luminal pH and creating an acidic environment that inhibits the proliferation of pathogenic bacteria. A reduction in butyrate levels typically increases pH. Faecal pH is also regarded as a marker of faecal health [
52].
This study hypothesised that the faecal pH would be lower in the groups fed BSFL fat due to the observed lower total SCFA concentrations. However, no decrease in faecal pH was detected in these groups, likely due to elevated faecal ammonia levels. Despite these differences, faecal pH remained within normal physiological limits. Similarly, Jian et al. [
28] reported no significant changes in faecal pH in dogs fed diets containing BSFL fat as a replacement for poultry fat, which agrees with the findings of the present study. In this study, the crude BSFL fat used in the diets was noted to have an unpleasant odour compared to poultry fat. Previous research has indicated that various residues and components in larval fat can influence its flavour [
11]. Understandably, dogs prefer poultry fat, which they are accustomed to, over the unfamiliar larval fat. Additionally, the higher linoleic acid content in poultry fat compared to larval fat may also influence the preference of dogs [
1]. Although Freel et al. [
27] have reported that dogs readily accepted BSFL fat, their study measured the consumption of larval fat-containing diets without directly comparing them to a control group. Furthermore, the larval fat used in their study contained 38.4% lauric acid and 17.5% linoleic acid, whereas the BSFL fat in the present study contained 42.84% lauric acid and 9.90% linoleic acid. In a study by Schiavone et al. [
17], no significant differences were observed in the preference of broiler chickens between soybean oil and BSFL fat, further highlighting the variability in fat preferences across species.
Currently, there is no established limit for MDA levels in dog foods. In this study, the MDA levels of the poultry oil and BSFL fat were not analysed prior to their incorporation into the diets. However, measurements conducted immediately after food production gave an initial MDA value of 1.84 mg/kg, suggesting that the MDA levels in the oils could be substantially higher. Given the variability in measurement methods and the wide range of reported values in the literature, determining whether the diets were oxidised remains challenging. For instance, Larouche et al. [
52] reported an MDA concentration of 2.8 mg/kg in BSFL dried at 60 °C for 30 min prior to degreasing. Similarly, Zhen et al. [
53] observed MDA levels exceeding 3.6 mg/kg in dried larvae using various methods prior to degreasing and storage for 1 month. In the current work, malondialdehyde levels increased from 2.17 to 5.72 mg/kg (2.6 times) in the control group and from 1.50 to 9.63 mg/kg (6.4 times) in the BSF6 group after 10 months.


 ,
, 


{kind=link}
{kind=link}