

The CRISPR/Cas9-Mediated Knockout of VgrG2 in Wild Pathogenic E. coli to Alleviate the Effects on Cell Damage and Autophagy

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cells, Strains and Plasmids
2.2. PCR Detection of the Virulence Gene VgrG2
2.3. Resistance Screening
2.4. Construction of Knockout Vectors
2.5. CRISPR/Cas9 Gene Editing Strategy for E. coli
2.6. Determination of the Growth Curve and Relative Cell Growth Rate
2.7. Cell Infection Experiments
2.8. RT-qPCR Detection
2.9. Cellular Immunofluorescence Detection
2.10. Statistical Analysis
3. Results
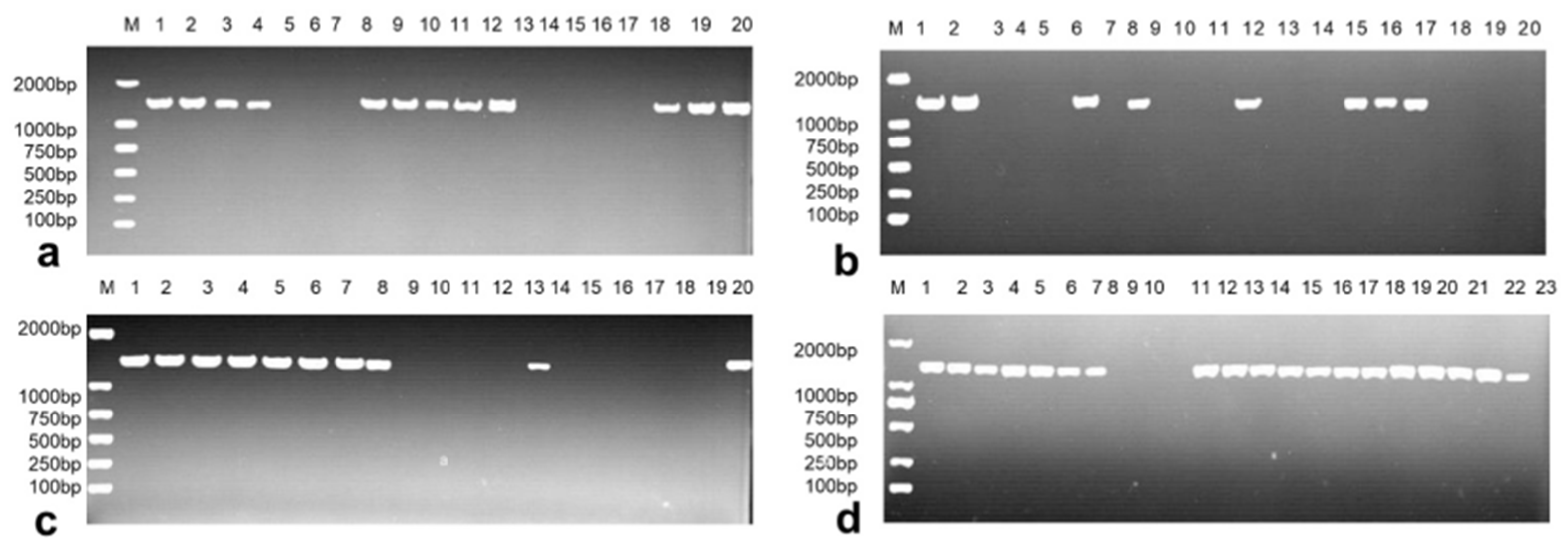
3.1. Identification and Resistance Screening of the Virulence Gene VgrG2 in Pathogenic E. coli
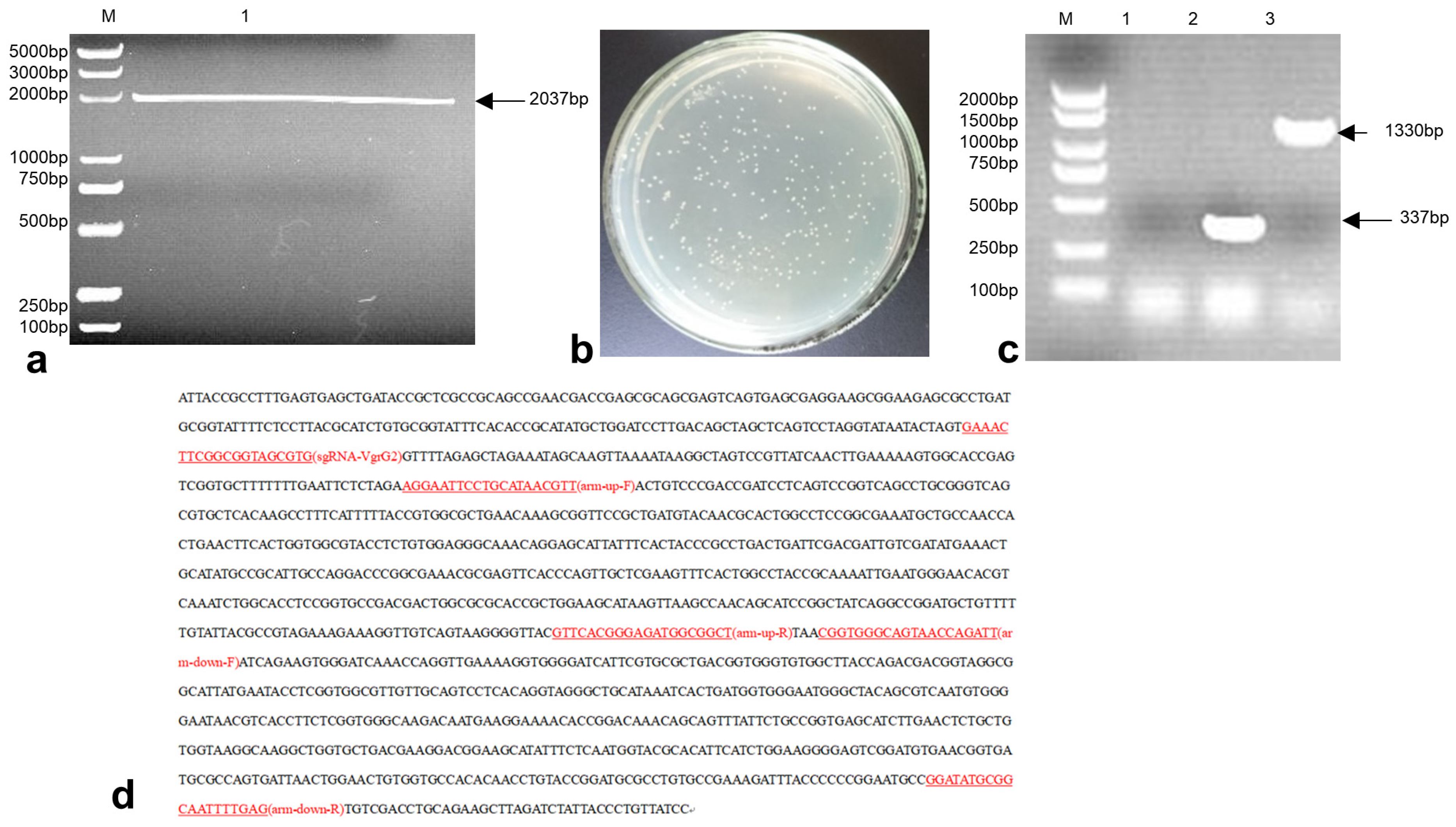
3.2. Construction of the E. coli-VgrG2 Targeting Vector pTargetF-sgRNA-VgrG2-Donor
3.3. Construction of the E. coli VgrG2 Virulence Gene Deletion Strain
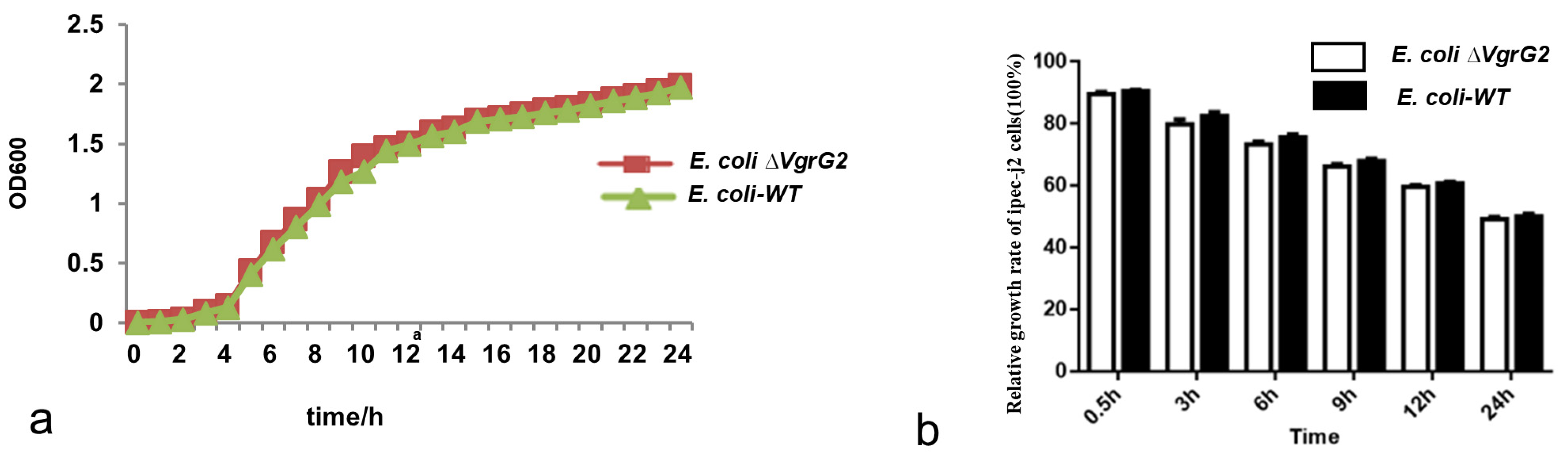
3.4. The Effect of the VgrG2 Virulence Gene Deletion on the E. coli Growth Rate
3.5. The Effect of E. coli-VgrG2 on IPEC-J2 Cell Growth
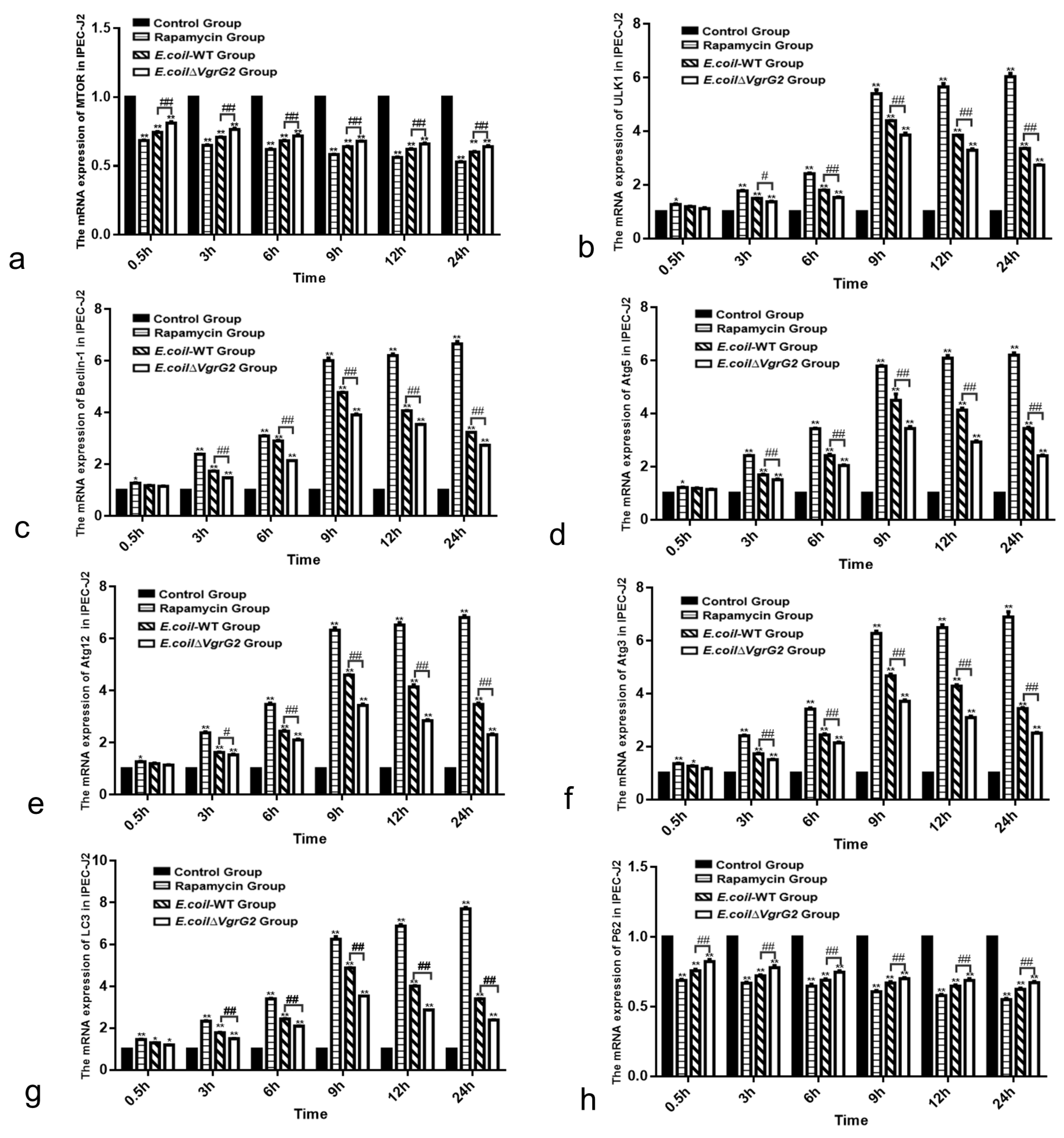
3.6. E. coli VgrG2 Activates the mTOR Signaling Pathway in IPEC-J2 Cells
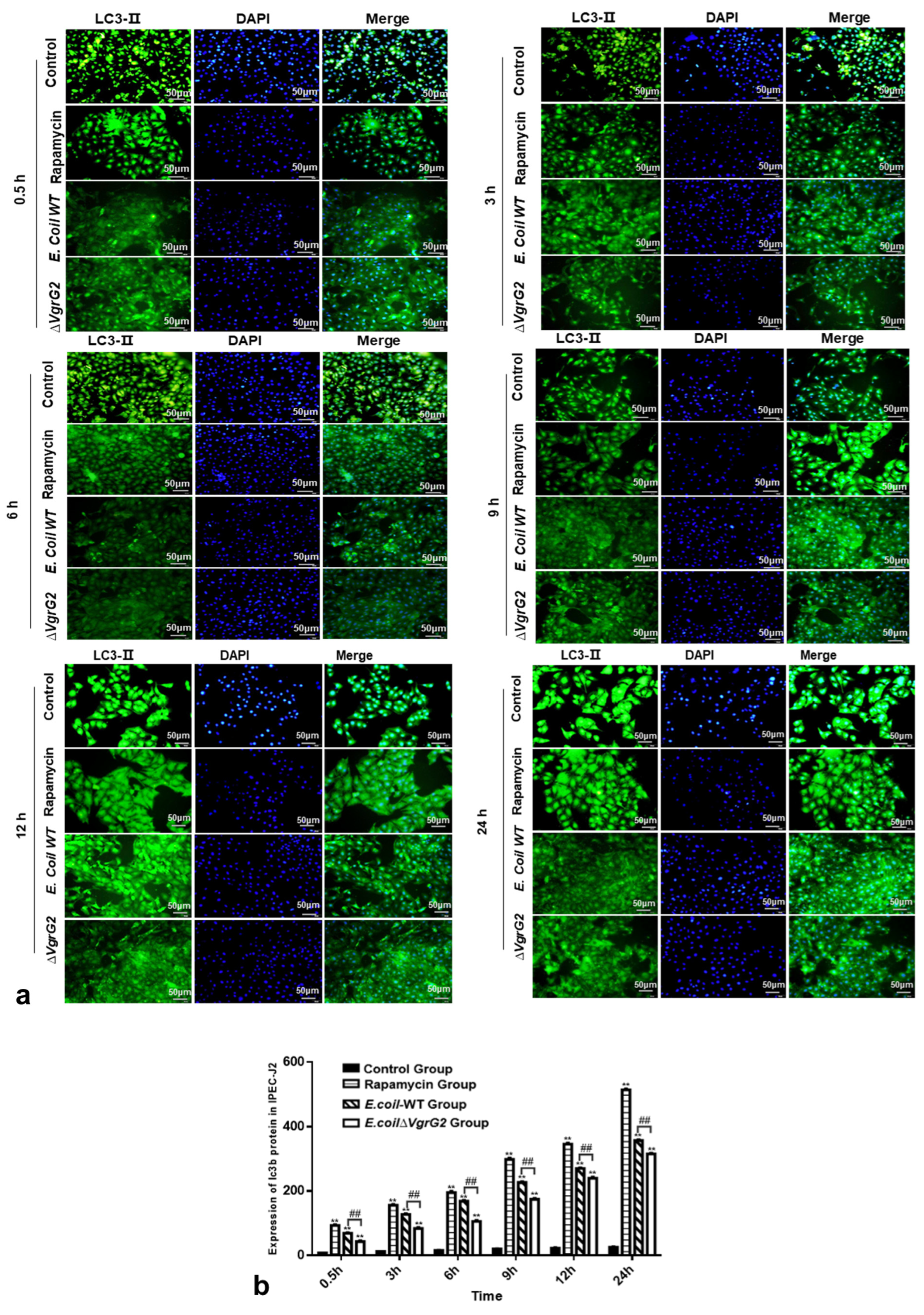
3.7. The Effect of E. coli VgrG2 on the Expression of the Autophagy Marker Protein LC3-II
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.Z.; Rehman, A.; Nasir, H.; Asif, A.; Tufail, B.; Usama, M.; Jabbar, B. Advances In Research On Genome Editing Crispr-Cas9 Technology. J. Ayub Med. Coll. Abbottabad JAMC 2019, 31, 108–122. [Google Scholar] [PubMed]
- Redman, M.; King, A.; Watson, C.; King, D. What is CRISPR/Cas9? Arch. Dis. Child. Educ. Pract. Ed. 2016, 101, 213–215. [Google Scholar] [CrossRef] [PubMed]
- El-Mounadi, K.; Morales-Floriano, M.L.; Garcia-Ruiz, H. Principles, Applications, and Biosafety of Plant Genome Editing Using CRISPR-Cas9. Front. Plant Sci. 2020, 11, 56. [Google Scholar] [CrossRef] [PubMed]
- Hsieh-Feng, V.; Yang, Y. Efficient expression of multiple guide RNAs for CRISPR/Cas genome editing. aBIOTECH 2020, 1, 123–134. [Google Scholar] [CrossRef]
- Makarova, K.S.; Grishin, N.V.; Shabalina, S.A.; Wolf, Y.I.; Koonin, E.V. A putative RNA-interference-based immune system in prokaryotes: Computational analysis of the predicted enzymatic machinery, functional analogies with eukaryotic RNAi, and hypothetical mechanisms of action. Biol. Direct 2006, 1, 7. [Google Scholar] [CrossRef]
- Zhang, B.; Wang, H.; Zhao, W.; Shan, C.; Liu, C.; Gao, L.; Zhao, R.; Ao, P.; Xiao, P.; Lv, L.; et al. New insights into the construction of wild-type Saba pig-derived Escherichia coli irp2 gene deletion strains. 3 Biotech 2021, 11, 408. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Ceremuga, M.; Krzowski, L.; Saluk-Bijak, J.; Bijak, M. Various Aspects of a Gene Editing System-CRISPR-Cas9. Int. J. Mol. Sci. 2020, 21, 9604. [Google Scholar] [CrossRef]
- Jiang, Y.; Chen, B.; Duan, C.; Sun, B.; Yang, J.; Yang, S. Multigene editing in the Escherichia coli genome via the CRISPR-Cas9 system. Appl. Environ. Microbiol. 2015, 81, 2506–2514. [Google Scholar] [CrossRef]
- Wang, X.; He, J.; Le, K. Making point mutations in Escherichia coli BL21 genome using the CRISPR-Cas9 system. FEMS Microbiol. Lett. 2018, 365, fny060. [Google Scholar] [CrossRef]
- Mougous, J.D.; Cuff, M.E.; Raunser, S.; Shen, A.; Zhou, M.; Gifford, C.A.; Goodman, A.L.; Joachimiak, G.; Ordoñez, C.L.; Lory, S.; et al. A virulence locus of Pseudomonas aeruginosa encodes a protein secretion apparatus. Science 2006, 312, 1526–1530. [Google Scholar] [CrossRef] [PubMed]
- Monjarás Feria, J.; Valvano, M.A. An Overview of Anti-Eukaryotic T6SS Effectors. Front. Cell. Infect. Microbiol. 2020, 10, 584751. [Google Scholar] [CrossRef] [PubMed]
- Wettstadt, S.; Filloux, A. Manipulating the type VI secretion system spike to shuttle passenger proteins. PLoS ONE 2020, 15, e0228941. [Google Scholar] [CrossRef] [PubMed]
- De Maayer, P.; Venter, S.N.; Kamber, T.; Duffy, B.; Coutinho, T.A.; Smits, T.H. Comparative genomics of the Type VI secretion systems of Pantoea and Erwinia species reveals the presence of putative effector islands that may be translocated by the VgrG and Hcp proteins. BMC Genom. 2011, 12, 576. [Google Scholar] [CrossRef] [PubMed]
- Dutta, P.; Jijumon, A.S.; Mazumder, M.; Dileep, D.; Mukhopadhyay, A.K.; Gourinath, S.; Maiti, S. Presence of actin binding motif in VgrG-1 toxin of Vibrio cholerae reveals the molecular mechanism of actin cross-linking. Int. J. Biol. Macromol. 2019, 133, 775–785. [Google Scholar] [CrossRef]
- Ma, J.; Sun, M.; Pan, Z.; Lu, C.; Yao, H. Diverse toxic effectors are harbored by vgrG islands for interbacterial antagonism in type VI secretion system. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 1635–1643. [Google Scholar] [CrossRef]
- Xu, T.; Liu, R.; Lu, X.; Wu, X.; Heneberg, P.; Mao, Y.; Jiang, Q.; Loor, J.; Yang, Z. Lycium barbarum polysaccharides alleviate LPS-induced inflammatory responses through PPARγ/MAPK/NF-κB pathway in bovine mammary epithelial cells. J. Anim. Sci. 2022, 100, skab345. [Google Scholar] [CrossRef]
- Braun, V.; Hantke, K. Lipoproteins: Structure, Function, Biosynthesis. Subcell. Biochem. 2019, 92, 39–77. [Google Scholar]
- Xu, L.; Brink, M. mTOR, cardiomyocytes and inflammation in cardiac hypertrophy. Biochim. Biophys. Acta 2016, 1863, 1894–1903. [Google Scholar] [CrossRef]
- Surviladze, Z.; Sterk, R.T.; DeHaro, S.A.; Ozbun, M.A. Cellular entry of human papillomavirus type 16 involves activation of the phosphatidylinositol 3-kinase/Akt/mTOR pathway and inhibition of autophagy. J. Virol. 2013, 87, 2508–2517. [Google Scholar] [CrossRef]
- Chang, H.; Li, X.; Cai, Q.; Li, C.; Tian, L.; Chen, J.; Xing, X.; Gan, Y.; Ouyang, W.; Yang, Z. The PI3K/Akt/mTOR pathway is involved in CVB3-induced autophagy of HeLa cells. Int. J. Mol. Med. 2017, 40, 182–192. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Fang, L.; Zhang, Y.; Sheng, H.; Fang, W. VgrG2 of type VI secretion system 2 of Vibrio parahaemolyticus induces autophagy in macrophages. Front. Microbiol. 2015, 6, 168. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Guan, K.L. mTOR: A pharmacologic target for autophagy regulation. J. Clin. Investig. 2015, 125, 25–32. [Google Scholar] [CrossRef]
- Tanida, I.; Ueno, T.; Kominami, E. LC3 and Autophagy. Methods Mol. Biol. 2008, 445, 77–88. [Google Scholar] [PubMed]
- Li, Q.; Sun, B.; Chen, J.; Zhang, Y.; Jiang, Y.; Yang, S. A modified pCas/pTargetF system for CRISPR-Cas9-assisted genome editing in Escherichia coli. Acta Biochim. Biophys. Sin. 2021, 53, 620–627. [Google Scholar] [CrossRef]
- Li, X.; He, S.; Ma, B. Autophagy and autophagy-related proteins in cancer. Mol. Cancer 2020, 19, 12. [Google Scholar] [CrossRef]
- Denton, D.; Kumar, S. Autophagy-dependent cell death. Cell Death Differ. 2019, 26, 605–616. [Google Scholar] [CrossRef]
- Green, D.R.; Levine, B. To be or not to be? How selective autophagy and cell death govern cell fate. Cell 2014, 157, 65–75. [Google Scholar] [CrossRef]
- Liu, Y.; Levine, B. Autosis and autophagic cell death: The dark side of autophagy. Cell Death Differ. 2015, 22, 367–376. [Google Scholar] [CrossRef]
- Renna, F.J.; Herrera Lopez, M.; Manifava, M.; Ktistakis, N.T.; Vaccaro, M.I. Evaluating Autophagy Levels in Two Different Pancreatic Cell Models Using LC3 Immunofluorescence. J. Vis. Exp. JoVE 2023. [CrossRef]
- Kabeya, Y.; Mizushima, N.; Ueno, T.; Yamamoto, A.; Kirisako, T.; Noda, T.; Kominami, E.; Ohsumi, Y.; Yoshimori, T. LC3, a mammalian homologue of yeast Apg8p, is localized in autophagosome membranes after processing. EMBO J. 2000, 19, 5720–5728. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, Y.; Zhou, Y.; Du, H.; Zhang, C.; Zhao, Z.; Chen, Y.; Zhou, Z.; Mei, J.; Wu, W.; et al. High fat diet-induced obesity leads to depressive and anxiety-like behaviors in mice via AMPK/mTOR-mediated autophagy. Exp. Neurol. 2022, 348, 113949. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pan, T.-L.; Cha, J.-L.; Wang, H.; Zhang, J.-S.; Xiao, J.-L.; Shen, J.; Zhou, M.; Li, Y.; Ma, J.-Z.; Zhao, K.-Y.; et al. The CRISPR/Cas9-Mediated Knockout of VgrG2 in Wild Pathogenic E. coli to Alleviate the Effects on Cell Damage and Autophagy. Vet. Sci. 2025, 12, 249. https://doi.org/10.3390/vetsci12030249
Pan T-L, Cha J-L, Wang H, Zhang J-S, Xiao J-L, Shen J, Zhou M, Li Y, Ma J-Z, Zhao K-Y, et al. The CRISPR/Cas9-Mediated Knockout of VgrG2 in Wild Pathogenic E. coli to Alleviate the Effects on Cell Damage and Autophagy. Veterinary Sciences. 2025; 12(3):249. https://doi.org/10.3390/vetsci12030249
Chicago/Turabian StylePan, Tian-Ling, Jin-Long Cha, Hao Wang, Jing-Song Zhang, Jin-Long Xiao, Jue Shen, Meng Zhou, Yue Li, Jin-Zhi Ma, Kai-Yuan Zhao, and et al. 2025. "The CRISPR/Cas9-Mediated Knockout of VgrG2 in Wild Pathogenic E. coli to Alleviate the Effects on Cell Damage and Autophagy" Veterinary Sciences 12, no. 3: 249. https://doi.org/10.3390/vetsci12030249
APA StylePan, T.-L., Cha, J.-L., Wang, H., Zhang, J.-S., Xiao, J.-L., Shen, J., Zhou, M., Li, Y., Ma, J.-Z., Zhao, K.-Y., Zhang, Y.-K., Xiao, P., & Gao, H. (2025). The CRISPR/Cas9-Mediated Knockout of VgrG2 in Wild Pathogenic E. coli to Alleviate the Effects on Cell Damage and Autophagy. Veterinary Sciences, 12(3), 249. https://doi.org/10.3390/vetsci12030249