
The Latest Prevalence, Isolation, and Molecular Characteristics of Feline Herpesvirus Type 1 in Yanji City, China

 , , ,
, , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Samples and Cells
2.2. Isolation and Identification of Viruses
2.3. Transmission Electron Microscopy (TEM)
2.4. Indirect Immunofluorescence Identification
2.5. T-A Cloning and Molecular Epidemiology of FHV-1
2.6. DNA Sequencing and Phylogenetic Analysis
3. Results
3.1. Preliminary Identification of Clinical Samples
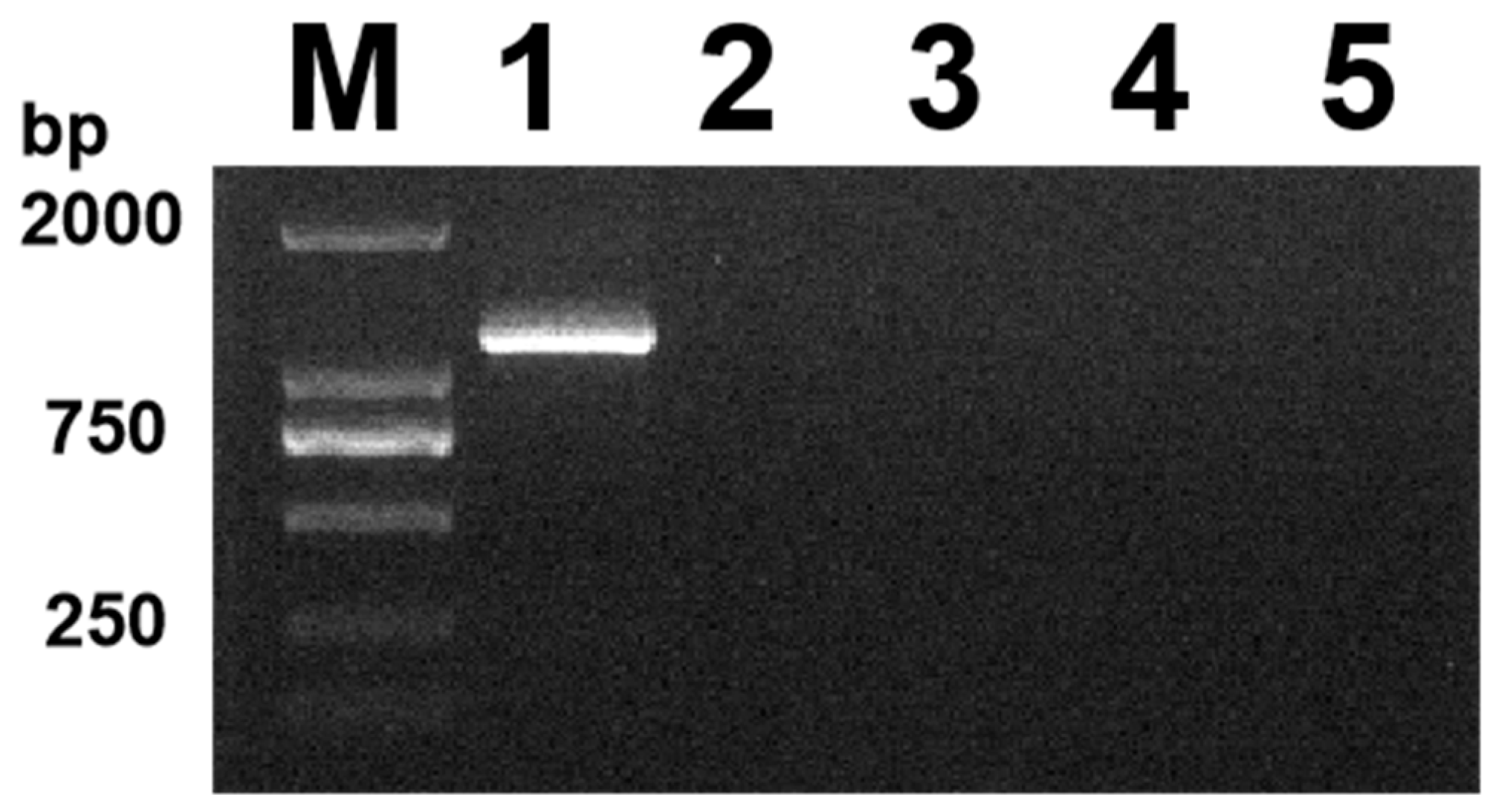
3.2. Data Processing of Positive Samples
3.3. Sequence Analysis of the gD Gene
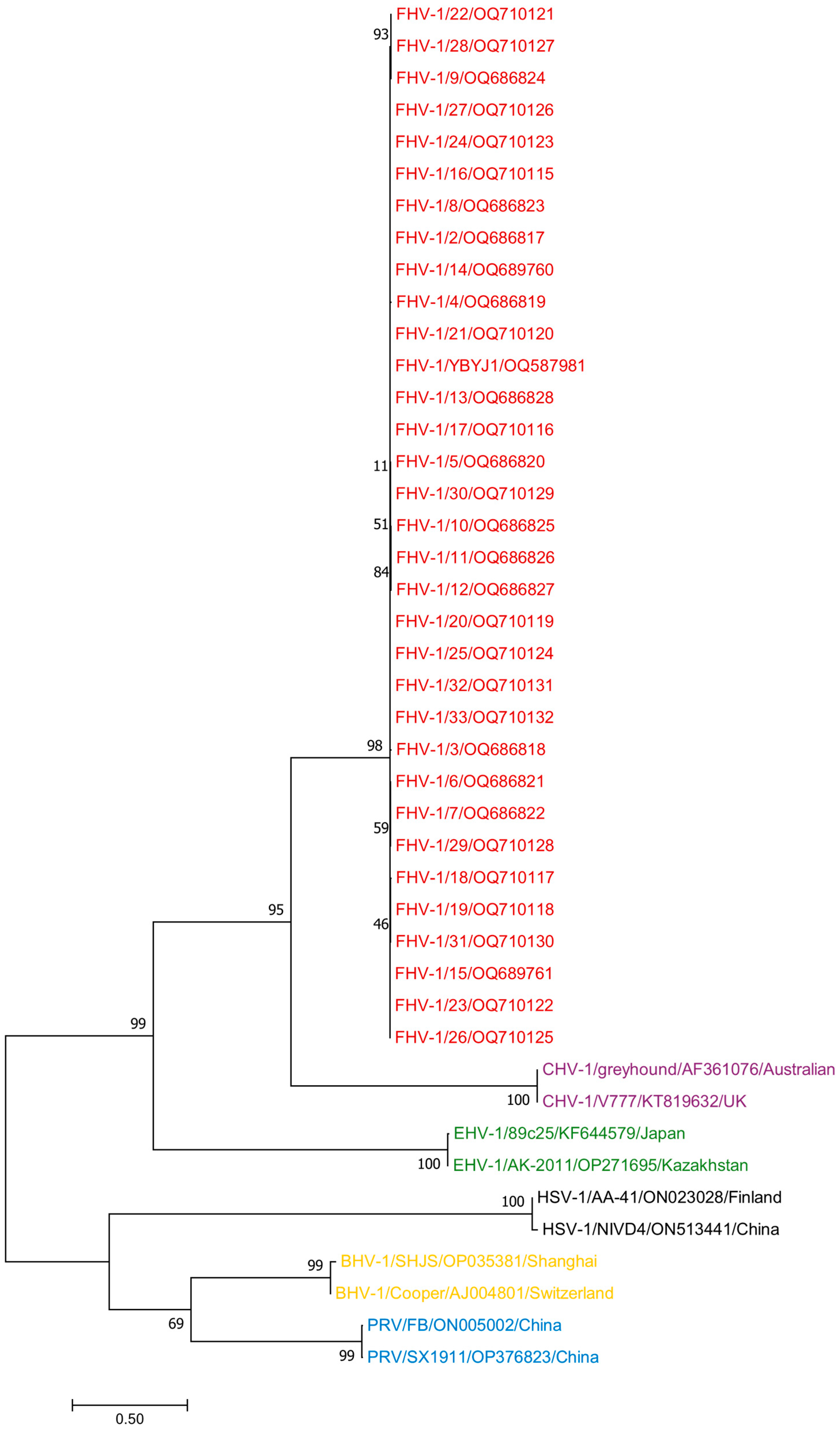
3.4. Phylogenetic Analysis
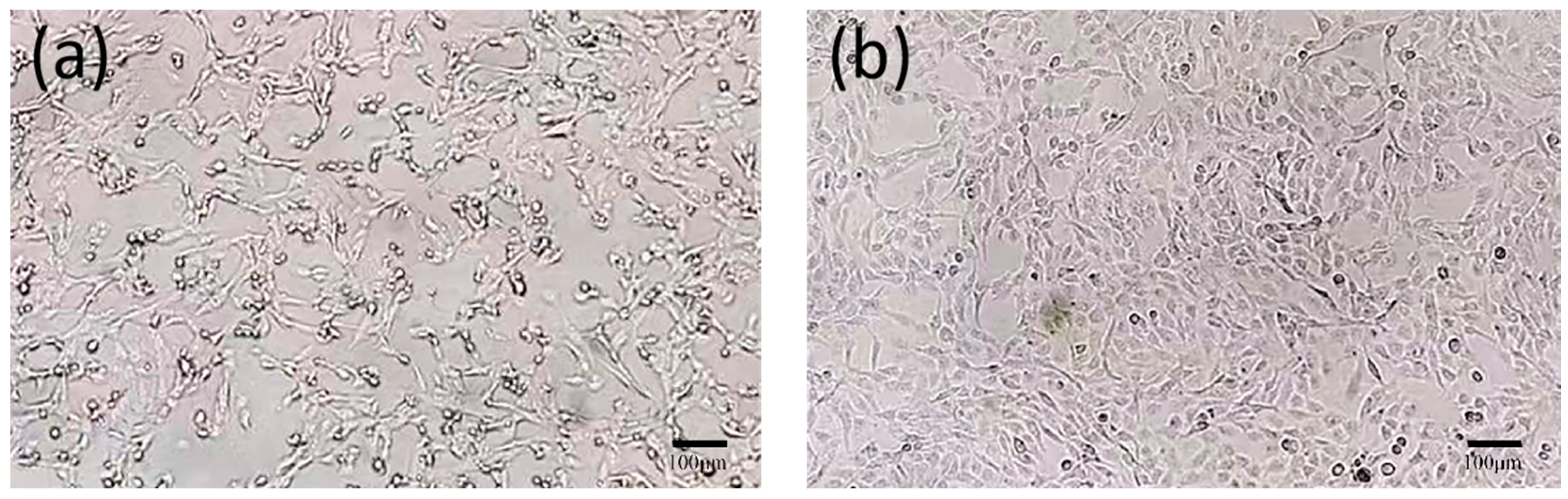
3.5. Virus Isolation
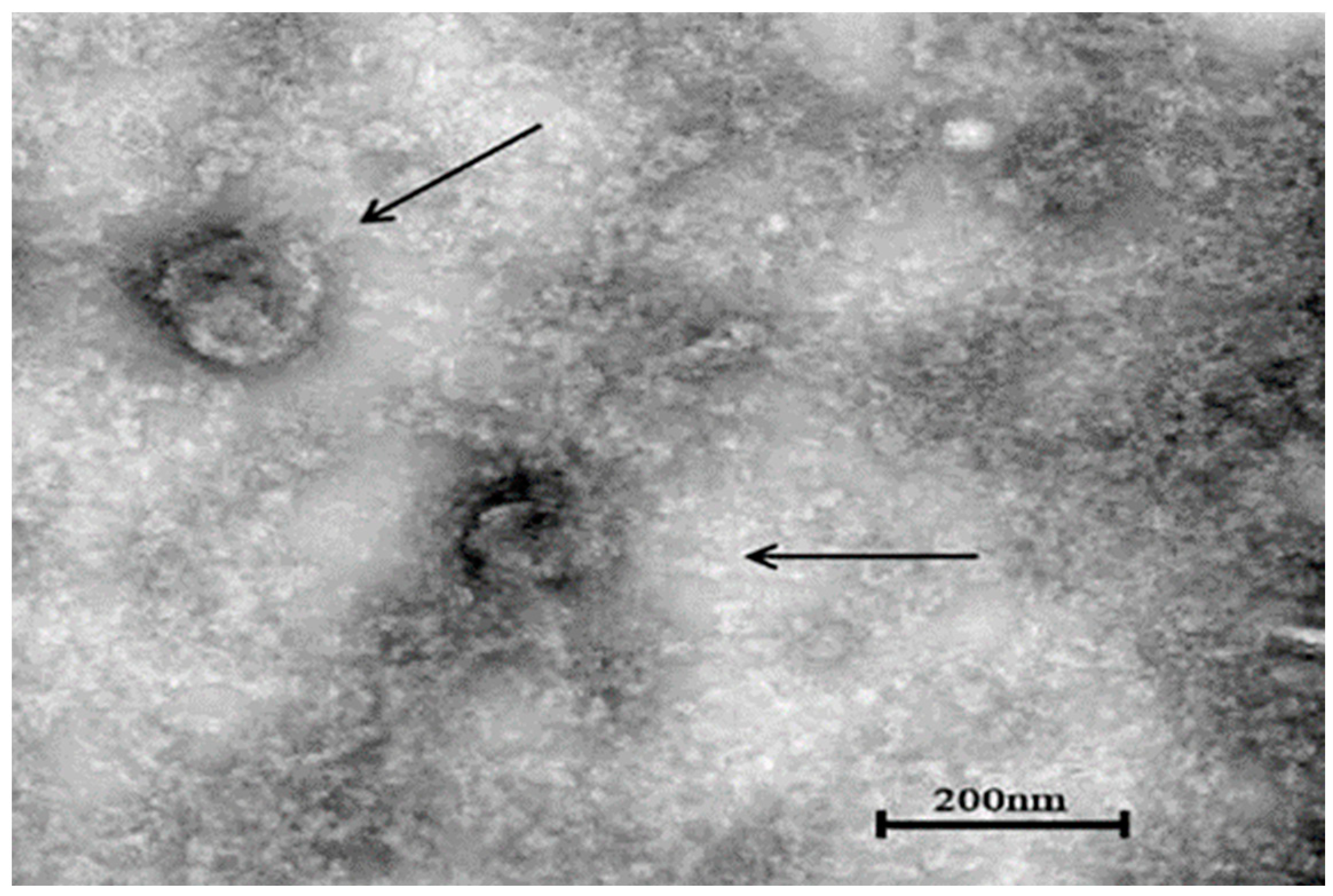
3.6. Transmission Electron Microscopy
3.7. Indirect Immunofluorescence Identification
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gaskell, R.; Dawson, S.; Radford, A.; Thiry, E. Feline herpesvirus. Vet. Res. 2007, 38, 337–354. [Google Scholar] [CrossRef] [PubMed]
- Crandell, R.A.; Maurer, F.D. Isolation of a feline virus associated with intranuclear inclusion bodies. Proc. Soc. Exp. Biol. Med. 1958, 97, 487–490. [Google Scholar] [CrossRef]
- Hartley, C. Aetiology of corneal ulcers assume FHV-1 unless proven otherwise. J. Feline Med. Surg. 2010, 12, 24–35. [Google Scholar] [CrossRef]
- Cohn, L.A. Feline respiratory disease complex. Vet. Clin. N. Am. Small Anim. Pract. 2011, 41, 1273–1289. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.T.; Kelman, M.; Ward, M.P. Surveillance of upper respiratory tract disease in owned cats in Australia, 2009–2012. Prev. Vet. Med. 2013, 112, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Chan, I.; Dowsey, A.; Lait, P.; Tasker, S.; Blackwell, E.; Helps, C.R.; Barker, E.N. Prevalence and risk factors for common respiratory pathogens within a cohort of pet cats in the UK. J. Small Anim. Pract. 2023, 64, 552–560. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, R.M.; Povey, R.C. Feline viral rhinotracheitis: Sites of virus replication and persistence in acutely and persistently infected cats. Res. Vet. Sci. 1979, 27, 167–174. [Google Scholar] [CrossRef]
- Cavalheiro, J.B.; Echeverria, J.T.; Ramos, C.A.N.; Babo-Terra, V.J. Frequency of feline herpesvirus 1 (FHV-1) in domestic cats from Campo Grande, MS, Brazil. An. Acad. Bras. Cienc. 2023, 95 (Suppl. 2), e20221010. [Google Scholar] [CrossRef]
- Dinnage, J.D.; Scarlett, J.M.; Richards, J.R. Descriptive epidemiology of feline upper respiratory tract disease in an animal shelter. J. Feline Med. Surg. 2009, 11, 816–825. [Google Scholar] [CrossRef]
- Rota, P.A.; Maes, R.K.; Ruyechan, W.T. Physical characterization of the genome of feline herpesvirus-1. Virology 1986, 154, 168–179. [Google Scholar] [CrossRef]
- Grail, A.; Harbour, D.A.; Chia, W. Restriction endonuclease mapping of the genome of feline herpesvirus type 1. Arch. Virol. 1991, 116, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Maes, R.; Kiupel, M.; Nauwynck, H.; Soboll Hussey, G. Characterization of feline herpesvirus-1 deletion mutants in tissue explant cultures. Virus Res. 2020, 284, 197981. [Google Scholar] [CrossRef]
- Maeda, K.; Horimoto, T.; Mikami, T. Properties and functions of feline herpesvirus type 1 glycoproteins. J. Vet. Med. Sci. 1998, 60, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Spatz, S.J.; Rota, P.A.; Maes, R.K. Identification of the feline herpesvirus type 1 (FHV-1) genes encoding glycoproteins G, D, I and E: Expression of FHV-1 glycoprotein D in vaccinia and raccoon poxviruses. J. Gen. Virol. 1994, 75, 1235–1244. [Google Scholar] [CrossRef]
- Gaskell, R.M.; Povey, R.C. Re-excretion of feline viral rhinotracheitis virus following corticosteroid treatment. Vet. Rec. 1973, 93, 204–205. [Google Scholar] [CrossRef]
- Möstl, K.; Egberink, H.; Addie, D.; Frymus, T.; Boucraut-Baralon, C.; Truyen, U.; Hartmann, K.; Lutz, H.; Gruffydd-Jones, T.; Radford, A.D.; et al. Prevention of infectious diseases in cat shelters: ABCD guidelines. J. Feline Med. Surg. 2013, 15, 546–554. [Google Scholar] [CrossRef]
- Orr, C.M.; Gaskell, C.J.; Gaskell, R.M. Interaction of a combined feline viral rhinotracheitis-feline calicivirus vaccine and the FVR carrier state. Vet. Rec. 1978, 103, 200–202. [Google Scholar] [CrossRef] [PubMed]
- Di Martino, B.; Di Francesco, C.E.; Meridiani, I.; Marsilio, F. Etiological investigation of multiple respiratory infections in cats. New Microbiol. 2007, 30, 455–461. [Google Scholar]
- McManus, C.M.; Levy, J.K.; Andersen, L.A.; McGorray, S.P.; Leutenegger, C.M.; Gray, L.K.; Hilligas, J.; Tucker, S.J. Prevalence of upper respiratory pathogens in four management models for unowned cats in the Southeast United States. Vet. J. 2014, 201, 196–201. [Google Scholar] [CrossRef]
- Helps, C.R.; Lait, P.; Damhuis, A.; Björnehammar, U.; Bolta, D.; Brovida, C.; Chabanne, L.; Egberink, H.; Ferrand, G.; Fontbonne, A.; et al. Factors associated with upper respiratory tract disease caused by feline herpesvirus, feline calicivirus, Chlamydophila felis and Bordetella bronchiseptica in cats: Experience from 218 European catteries. Vet. Rec. 2005, 156, 669–673. [Google Scholar] [CrossRef]
- Spada, E.; Vitale, F.; Bruno, F.; Castelli, G.; Reale, S.; Perego, R.; Baggiani, L.; Proverbio, D. A pre- and during Pandemic Survey of Sars-Cov-2 Infection in Stray Colony and Shelter Cats from a High Endemic Area of Northern Italy. Viruses 2021, 13, 618. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Ju, H.; Li, X.; Shen, H.; Ge, F.; Yang, X.; Zhao, H.; Wu, X.; Zhu, X.; Wang, X.; et al. Epidemiological Surveillance of Respiratory Diseases in Urban Stray Cats in Shanghai. Animals 2024, 14, 1562. [Google Scholar] [CrossRef] [PubMed]
- Thiry, E.; Addie, D.; Belák, S.; Boucraut-Baralon, C.; Egberink, H.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lloret, A.; et al. Feline herpesvirus infection. ABCD guidelines on prevention and management. J. Feline Med. Surg. 2009, 11, 547–555. [Google Scholar] [CrossRef] [PubMed]
- Chvala-Mannsberger, S.; Bagó, Z.; Weissenböck, H. Occurrence, morphological characterization and antigen localization of felid herpesvirus-induced pneumonia in cats: A retrospective study (2000–2006). J. Comp. Pathol. 2009, 141, 163–169. [Google Scholar] [CrossRef]
- Hargis, A.M.; Ginn, P.E. Feline herpesvirus 1-associated facial and nasal dermatitis and stomatitis in domestic cats. Vet. Clin. N. Am. Small Anim. Pract. 1999, 29, 1281–1290. [Google Scholar] [CrossRef]
- Nakanishi, H.; Furuya, M.; Soma, T.; Hayashiuchi, Y.; Yoshiuchi, R.; Matsubayashi, M.; Tani, H.; Sasai, K. Prevalence of microorganisms associated with feline gingivostomatitis. J. Feline Med. Surg. 2019, 21, 103–108. [Google Scholar] [CrossRef]
- McGregor, G.F.; Sheehan, K.; Simko, E. Pneumonia and gastritis in a cat caused by feline herpesvirus-1. Can. Vet. J. = La Rev. Vet. Can. 2016, 57, 147–150. [Google Scholar]
- Gaskell, R.; Willoughby, K. Herpesviruses of carnivores. Vet. Microbiol. 1999, 69, 73–88. [Google Scholar] [CrossRef]
- Bergmann, M.; Speck, S.; Rieger, A.; Truyen, U.; Hartmann, K. Antibody response to feline herpesvirus-1 vaccination in healthy adult cats. J. Feline Med. Surg. 2020, 22, 329–338. [Google Scholar] [CrossRef]
- Walter, J.; Foley, P.; Yason, C.; Vanderstichel, R.; Muckle, A. Prevalence of feline herpesvirus-1, feline calicivirus, Chlamydia felis, and Bordetella bronchiseptica in a population of shelter cats on Prince Edward Island. Can. J. Vet. Res. = Rev. Can. Rech. Vet. 2020, 84, 181–188. [Google Scholar]
- Fernandez, M.; Manzanilla, E.G.; Lloret, A.; León, M.; Thibault, J.C. Prevalence of feline herpesvirus-1, feline calicivirus, Chlamydophila felis and Mycoplasma felis DNA and associated risk factors in cats in Spain with upper respiratory tract disease, conjunctivitis and/or gingivostomatitis. J. Feline Med. Surg. 2017, 19, 461–469. [Google Scholar] [CrossRef] [PubMed]
- Amoroso, M.G.; Serra, F.; Miletti, G.; Cardillo, L.; de Martinis, C.; Marati, L.; Alfano, F.; Ferrara, G.; Pagnini, U.; De Carlo, E.; et al. A Retrospective Study of Viral Molecular Prevalences in Cats in Southern Italy (Campania Region). Viruses 2022, 14, 2583. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Cheng, Y.; Fang, Z.; Zhongqi, Q.; Weidong, Y.; Yilmaz, A.; Yilmaz, H.; Umar, S. First report of molecular epidemiology and phylogenetic characteristics of feline herpesvirus (FHV-1) from naturally infected cats in Kunshan, China. Virol. J. 2024, 21, 115. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Liu, Y.; Qian, P.; Cao, Y.; Wang, J.; Sun, C.; Huang, B.; Cui, N.; Huo, N.; Wu, H.; et al. Molecular and serological investigation of cat viral infectious diseases in China from 2016 to 2019. Transbound. Emerg. Dis. 2020, 67, 2329–2335. [Google Scholar] [CrossRef] [PubMed]
- Murrell, B.; Moola, S.; Mabona, A.; Weighill, T.; Sheward, D.; Kosakovsky Pond, S.L.; Scheffler, K. FUBAR: A fast, unconstrained bayesian approximation for inferring selection. Mol. Biol. Evol. 2013, 30, 1196–1205. [Google Scholar] [CrossRef]
- Witte, C.L.; Lamberski, N.; Rideout, B.A.; Vaida, F.; Citino, S.B.; Barrie, M.T.; Haefele, H.J.; Junge, R.E.; Murray, S.; Hungerford, L.L. Epidemiology of clinical feline herpesvirus infection in zoo-housed cheetahs (Acinonyx jubatus). J. Am. Vet. Med. Assoc. 2017, 251, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, D.; Barrs, V.R.; Kelman, M.; Ward, M.P. Feline upper respiratory tract infection and disease in Australia. J. Feline Med. Surg. 2019, 21, 973–978. [Google Scholar] [CrossRef] [PubMed]
- Tran, V.; Kelman, M.; Ward, M.; Westman, M. Risk of Feline Immunodeficiency Virus (FIV) Infection in Pet Cats in Australia is Higher in Areas of Lower Socioeconomic Status. Animals 2019, 9, 592. [Google Scholar] [CrossRef] [PubMed]
- Holst, B.S.; Berndtsson, L.T.; Englund, L. Isolation of feline herpesvirus-1 and feline calicivirus from healthy cats in Swedish breeding catteries. J. Feline Med. Surg. 2005, 7, 325–331. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Breed | Age (Month) | Clinical Symptoms | Vaccine History | GenBank |
|---|---|---|---|---|---|
| 1 | Maine | 4 | Snot, sneeze | - | OQ587981 |
| 2 | American Shorthair | 10 | Eye conjunctivitis | ○ | OQ686817 |
| 3 | British short-haired cat | 6 | Conjunctivitis, sneezing | - | OQ686818 |
| 4 | Orange cat | 3 | Dispirited, fever | - | OQ686819 |
| 5 | British short-haired cat | 8 | Fever, eye secretions | - | OQ686820 |
| 6 | Ragdoll | 2 | Eye and nose secretions | - | OQ686821 |
| 7 | Ragdoll | 2 | Conjunctivitis, sneezing | - | OQ686822 |
| 8 | British short-haired cat | 10 | Sneezing | + | OQ686823 |
| 9 | Chinese Li Hua | 2 | Conjunctivitis, ocular secretions | - | OQ686824 |
| 10 | Chinese Li Hua | 5 | Cough, conjunctivitis | - | OQ686825 |
| 11 | Orange cat | 2 | Cough, fever | ○ | OQ686826 |
| 12 | American Shorthair | 7 | Conjunctivitis | - | OQ686827 |
| 13 | Pastoral cat | 9 | Dispirited, sneezing | + | OQ686828 |
| 14 | Siamese | 12 | Conjunctivitis, ocular secretions | ○ | OQ689760 |
| 15 | British short-haired cat | 5 | Eye and nose secretions | - | OQ689761 |
| 16 | Garfield | 5 | Conjunctivitis | - | OQ710115 |
| 17 | Maine | 6 | Conjunctivitis, sneezing | ○ | OQ710116 |
| 18 | British short-haired cat | 8 | Sneezing | - | OQ710117 |
| 19 | British short-haired cat | 3 | Eye rot bone | + | OQ710118 |
| 20 | Pastoral cat | 3 | Conjunctivitis, ocular secretions | - | OQ710119 |
| 21 | American Shorthair | 4 | Fever, eye and nose secretions | - | OQ710120 |
| 22 | American Shorthair | 7 | Eye and nose secretions, sneezing | - | OQ710121 |
| 23 | Maine | 6 | Conjunctivitis, sneezing | ○ | OQ710122 |
| 24 | Orange cat | 8 | Sneezing | + | OQ710123 |
| 25 | Garfield | 12 | Conjunctivitis, ocular secretions | ○ | OQ710124 |
| 26 | British short-haired cat | 2 | Eye and nose secretions, Dispirited | - | OQ710125 |
| 27 | American Shorthair | 5 | Fever, sneezing and coughing | - | OQ710126 |
| 28 | Sphynx | 2 | Fever | - | OQ710127 |
| 29 | Chinese Li Hua | 2 | Fever, sneezing | - | OQ710128 |
| 30 | Ragdoll | 3 | Conjunctivitis | ○ | OQ710129 |
| 31 | Maine | 5 | Fever, eye and nose secretions | - | OQ710130 |
| 32 | British short-haired cat | 7 | Conjunctivitis, ocular secretions | ○ | OQ710131 |
| 33 | American Shorthair | 4 | Fever, eye discharge | - | OQ710132 |
| Prevalence rate = (number of positive samples/total number of subjects) × 100% = 32.2% | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, M.; Mu, B.; Ma, H.; Xue, H.; Song, Y.; Zhu, K.; Hao, J.; Liu, D.; Li, W.; Zhang, Y.; et al. The Latest Prevalence, Isolation, and Molecular Characteristics of Feline Herpesvirus Type 1 in Yanji City, China. Vet. Sci. 2024, 11, 417. https://doi.org/10.3390/vetsci11090417
Yang M, Mu B, Ma H, Xue H, Song Y, Zhu K, Hao J, Liu D, Li W, Zhang Y, et al. The Latest Prevalence, Isolation, and Molecular Characteristics of Feline Herpesvirus Type 1 in Yanji City, China. Veterinary Sciences. 2024; 11(9):417. https://doi.org/10.3390/vetsci11090417
Chicago/Turabian StyleYang, Meng, Biying Mu, Haoyuan Ma, Haowen Xue, Yanhao Song, Kunru Zhu, Jingrui Hao, Dan Liu, Weijian Li, Yaning Zhang, and et al. 2024. "The Latest Prevalence, Isolation, and Molecular Characteristics of Feline Herpesvirus Type 1 in Yanji City, China" Veterinary Sciences 11, no. 9: 417. https://doi.org/10.3390/vetsci11090417
APA StyleYang, M., Mu, B., Ma, H., Xue, H., Song, Y., Zhu, K., Hao, J., Liu, D., Li, W., Zhang, Y., & Gao, X. (2024). The Latest Prevalence, Isolation, and Molecular Characteristics of Feline Herpesvirus Type 1 in Yanji City, China. Veterinary Sciences, 11(9), 417. https://doi.org/10.3390/vetsci11090417