
In Situ Expression of Yak IL-22 in Mammary Glands as a Treatment for Bovine Staphylococcus aureus-Induced Mastitis in Mice

Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bioinformatics Analysis
2.2. Vector Construction
2.3. Cells and Transfection
2.4. Antimicrobial Tests In Vitro
2.5. Animals
2.6. Mastitis Model Test in Mice
2.7. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
2.8. Inflammatory Cytokine Enzyme-Linked Immunosorbent Assay
2.9. Pathological Tests
2.10. Determination of Microbial Diversity
2.11. Statistics and Data Analysis
3. Results
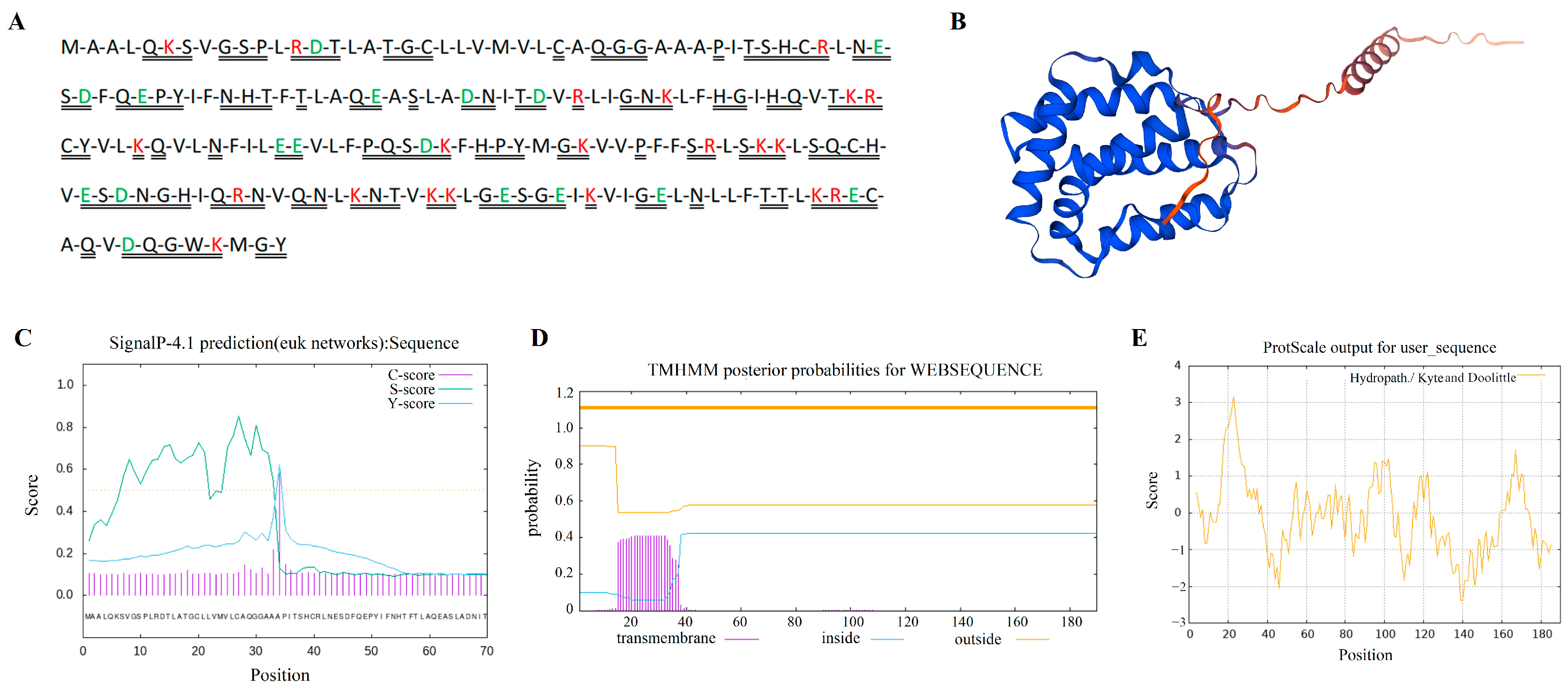
3.1. Bioinformatics Analysis of Yak IL-22 Gene
3.2. The Conversion of Inflammatory Factors In Vivo and In Vitro
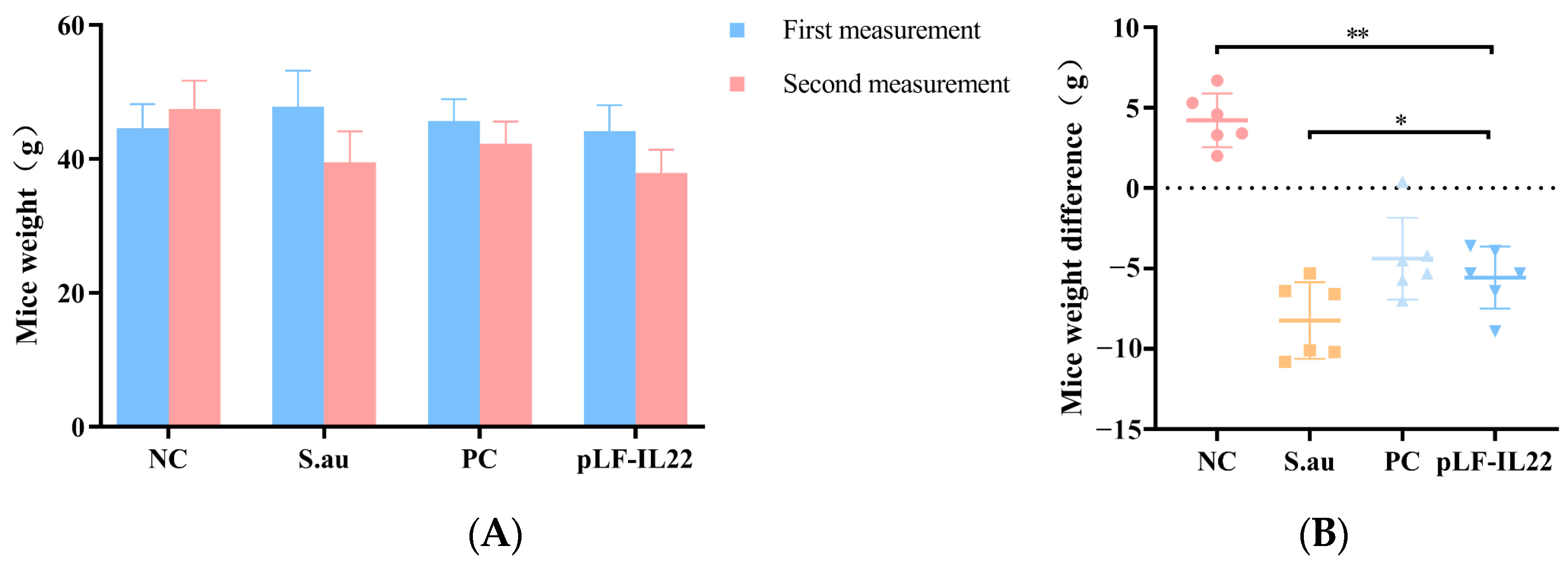
3.3. Body Weight Changes of Mice
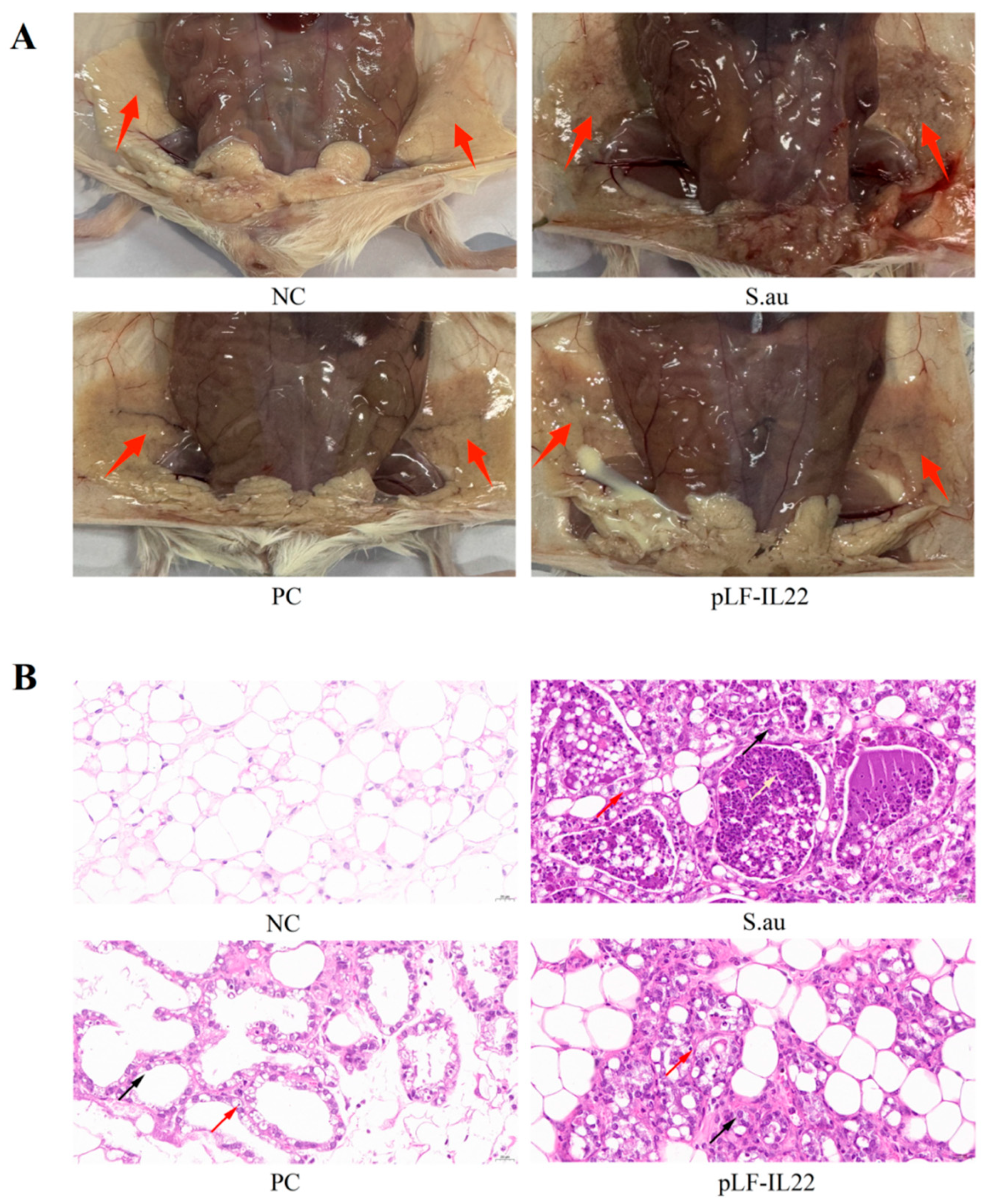
3.4. Histopathological Changes
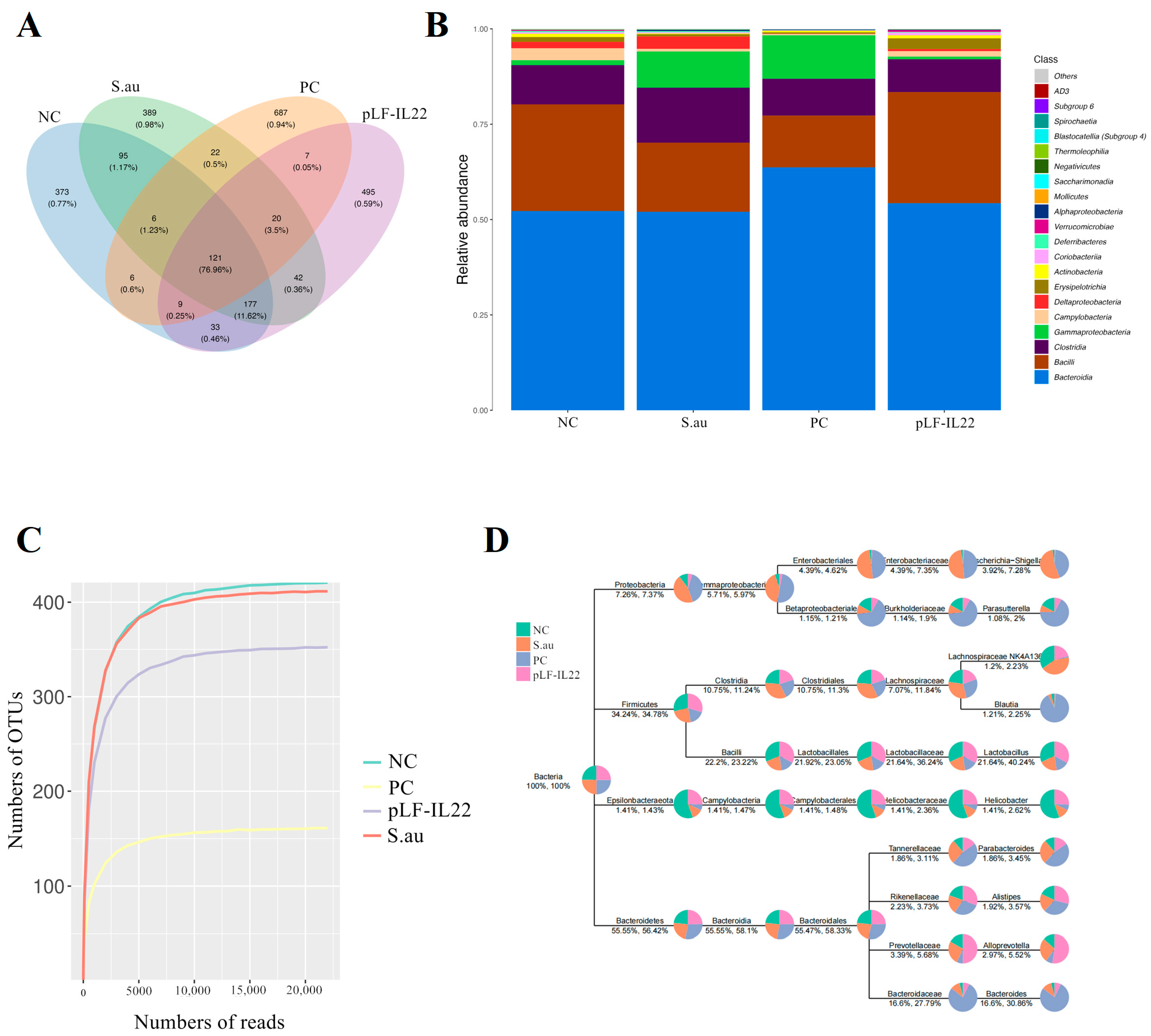
3.5. Effects on Composition of Intestinal Flora
3.6. Differences in Intestinal Flora and Prediction of Flora Function
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, W.; Ma, S.; Lei, L.; He, J.; Li, X.; Tao, J.; Wang, X.; Song, S.; Wang, Y.; Wang, Y.; et al. Prevalence, etiology, and economic impact of clinical mastitis on large dairy farms in China. Vet. Microbiol. 2020, 242, 108570. [Google Scholar] [CrossRef] [PubMed]
- Zaatout, N.; Hezil, D. A meta-analysis of the global prevalence of methicillin-resistant Staphylococcus aureus (MRSA) isolated from clinical and subclinical bovine mastitis. J. Appl. Microbiol. 2022, 132, 140–154. [Google Scholar]
- Sharifi, A.; Mahmoudi, P.; Sobhani, K. The prevalence of adhesion and biofilm genes in Staphylococcus aureus isolates from bovine mastitis: A comprehensive meta-analysis. Vet. Med. Sci. 2024, 10, e31378. [Google Scholar]
- Dahl, M.O.; De Vries, A.; Maunsell, F.P.; Galvao, K.N.; Risco, C.A.; Hernandez, J.A. Epidemiologic and economic analyses of pregnancy loss attributable to mastitis in primiparous Holstein cows. J. Dairy Sci. 2018, 101, 10142–10150. [Google Scholar] [PubMed]
- Zheng, Z.; Zheng, Y.; Liang, X.; Xue, G.; Wu, H. Sanguinarine Enhances the Integrity of the Blood-Milk Barrier and Inhibits Oxidative Stress in Lipopolysaccharide-Stimulated Mastitis. Cells 2022, 11, 3658. [Google Scholar]
- Das, M.S.; Bandopadhyay, S.; Jadhao, S.; Shome, R.; Shome, B.R. Genetic characterization and comparative genomics of a multi drug resistant (MDR) Escherichia coli SCM-21 isolated from a subclinical case of bovine mastitis. Comp. Immunol. Microbiol. Infect. Dis. 2022, 85, 101799. [Google Scholar]
- Ramachandra, B.; Manjunath, H.; Prabha, R.; Krishna, R. Residual effect of common antibiotics used for mastitis treatment at the farm level on the growth of Bacillus subtilis and Streptococcus thermophilus. Asian J. Dairy Food Res. 2012, 31, 95–98. [Google Scholar]
- Chen, Y.; Yang, J.; Huang, Z.; Yin, B.; Umar, T.; Yang, C.; Zhang, X.; Jing, H.; Guo, S.; Guo, M.; et al. Vitexin Mitigates Staphylococcus aureus-Induced Mastitis via Regulation of ROS/ER Stress/NF-kappaB/MAPK Pathway. Oxidative Med. Cell. Longev. 2022, 2022, 7977433. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.H.; Aggarwal, S.; Ho, W.H.; Foster, J.; Zhang, Z.; Stinson, J.; Wood, W.I.; Goddard, A.D.; Gurney, A.L. Interleukin (IL)-22, a novel human cytokine that signals through the interferon receptor-related proteins CRF2-4 and IL-22R. J. Biol. Chem. 2000, 275, 31335–31339. [Google Scholar]
- Starkey, M.R.; Plank, M.W.; Casolari, P.; Papi, A.; Pavlidis, S.; Guo, Y.; Cameron, G.J.; Haw, T.J.; Tam, A.; Obiedat, M.; et al. IL-22 and its receptors are increased in human and experimental COPD and contribute to pathogenesis. Eur. Respir. J. 2019, 54, 1800174. [Google Scholar] [CrossRef]
- Yu, D.; Yang, G.; Mo, J.; Zhang, M.; Xia, H.; Gan, Z.; Lu, Y. Identification and functional characterization of interleukin-22 (IL-22) in orange-spotted grouper (Epinephelus coioides). Fish Shellfish. Immunol. 2024, 150, 109598. [Google Scholar] [CrossRef] [PubMed]
- Sajiir, H.; Wong, K.Y.; Muller, A.; Keshvari, S.; Burr, L.; Aiello, E.; Mezza, T.; Giaccari, A.; Sebastiani, G.; Dotta, F.; et al. Pancreatic beta-cell IL-22 receptor deficiency induces age-dependent dysregulation of insulin biosynthesis and systemic glucose homeostasis. Nat. Commun. 2024, 15, 4527. [Google Scholar] [CrossRef]
- Toth, K.A.; Schmitt, E.G.; Kolicheski, A.; Greenberg, Z.J.; Levendosky, E.; Saucier, N.; Trammel, K.; Oikonomou, V.; Lionakis, M.S.; Klechevsky, E.; et al. A human STAT3 gain-of-function variant drives local Th17 dysregulation and skin inflammation in mice. J. Exp. Med. 2024, 221, e20232091. [Google Scholar] [CrossRef] [PubMed]
- Pravoverov, K.; Fatima, I.; Barman, S.; Jühling, F.; Primeaux, M.; Baumert, T.F.; Singh, A.B.; Dhawan, P. IL-22 regulates MASTL expression in intestinal epithelial cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2024, 327, G123–G139. [Google Scholar] [CrossRef]
- Zhang, T.; Seeger, P.; Simsek, Y.; Sabihi, M.; Lücke, J.; Zazara, D.E.; Shiri, A.M.; Kempski, J.; Blankenburg, T.; Zhao, L.; et al. IL-22 promotes liver regeneration after portal vein ligation. Heliyon 2024, 10, e27578. [Google Scholar] [CrossRef] [PubMed]
- Breugelmans, T.; Arras, W.; Osterlinck, B.; Jauregui-Amezaga, A.; Somers, M.; Cuypers, B.; Laukens, K.; De Man, J.G.; De Schepper, H.U.; De Winter, B.Y.; et al. IL-22-Activated MUC13 Impacts on Colonic Barrier Function through JAK1/STAT3, SNAI1/ZEB1 and ROCK2/MAPK Signaling. Cells 2023, 12, 1224. [Google Scholar] [CrossRef]
- Sajiir, H.; Keshvari, S.; Wong, K.Y.; Borg, D.J.; Steyn, F.J.; Fercher, C.; Taylor, K.; Taylor, B.; Barnard, R.T.; Müller, A.; et al. Liver and pancreatic-targeted interleukin-22 as a therapeutic for metabolic dysfunction-associated steatohepatitis. Nat. Commun. 2024, 15, 4528. [Google Scholar] [CrossRef]
- Zhu, Q.; Korenfeld, D.; Suarezfueyo, A.; Graham, S.; Jin, L.; Punit, S.; Duffy, R.; Puri, M.; Caruso, A.; Hu, C.; et al. Epithelial dysfunction is prevented by IL-22 treatment in a Citrobacter rodentium-induced colitis model that shares similarities with inflammatory bowel disease. Mucosal. Immunol. 2022, 15, 1338–1349. [Google Scholar] [CrossRef]
- Yang, R.; Chen, J.; Qu, X.; Liu, H.; Wang, X.; Tan, C.; Chen, H.; Wang, X. Interleukin-22 Contributes to Blood-Brain Barrier Disruption via STAT3/VEGFA Activation in Escherichia coli Meningitis. ACS Infect. Dis. 2024, 10, 988–999. [Google Scholar] [CrossRef]
- Schukken, Y.H.; Gunther, J.; Fitzpatrick, J.; Fontaine, M.; Goetze, L.; Holst, O.; Leigh, J.; Petzl, W.; Schuberth, H.-J.; Sipka, A.; et al. Host-response patterns of intramammary infections in dairy cows. Vet. Immunol. Immunopathol. 2011, 144, 270–289. [Google Scholar]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Xie, L.; Liu, S.; Dai, A.; Chi, X.; Zhang, D. Non-targeted metabolomics and microbial analyses of the impact of oat antimicrobial peptides on rats with dextran sulfate sodium-induced enteritis. Front. Nutr. 2022, 9, 1095483. [Google Scholar]
- Middleton, J.R.; Saeman, A.; Fox, L.K.; Lombard, J.; Hogan, J.S.; Smith, K.L. The National Mastitis Council: A Global Organization for Mastitis Control and Milk Quality, 50 Years and Beyond. J. Mammary Gland. Biol. Neoplasia 2014, 19, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, A.; Kamel, M. Bovine mastitis prevention and control in the post-antibiotic era. Trop. Anim. Health Prod. 2021, 53, 236. [Google Scholar]
- Tumanov, A.V.; Koroleva, E.P.; Guo, X.; Wang, Y.; Kruglov, A.; Nedospasov, S.; Fu, Y.-X. Lymphotoxin controls the IL-22 protection pathway in gut innate lymphoid cells during mucosal pathogen challenge. Cell Host Microbe 2011, 10, 44–53. [Google Scholar] [CrossRef]
- Dumoutier, L.; Van Roost, E.; Colau, D.; Renauld, J.C. Human interleukin-10-related T cell-derived inducible factor: Molecular cloning and functional characterization as an hepatocyte-stimulating factor. Proc. Natl. Acad. Sci. USA 2000, 97, 10144–10149. [Google Scholar]
- Keir, M.; Yi, Y.; Lu, T.; Ghilardi, N. The role of IL-22 in intestinal health and disease. J. Exp. Med. 2020, 217, e20192195. [Google Scholar] [CrossRef]
- OUYANG, W. Distinct roles of IL-22 in human psoriasis and inflammatory bowel disease. Cytokine Growth Factor Rev. 2010, 21, 435–441. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, L.; Tan, L.; Zhang, C.; Li, X.; Wang, P.; Gao, L.; Zhao, C. Interleukin-22 Inhibits Apoptosis of Gingival Epithelial Cells Through TGF-beta Signaling Pathway During Periodontitis. Inflammation 2023, 46, 1871–1886. [Google Scholar]
- Agonia, I.; Couras, J.; Cunha, A.; Andrade, A.J.; Macedo, J.; Sousa-Pinto, B. IL-17, IL-21 and IL-22 polymorphisms in rheumatoid arthritis: A systematic review and meta-analysis. Cytokine 2020, 125, 154813. [Google Scholar]
- Rainard, P.; Cunha, P.; Martins, R.P.; Gilbert, F.B.; Germon, P.; Foucras, G. Type 3 immunity: A perspective for the defense of the mammary gland against infections. Vet. Res. 2020, 51, 129. [Google Scholar] [CrossRef]
- Xiao, Z.; Liu, L.; Jin, Y.; Pei, X.; Sun, W.; Wang, M. Clostridium tyrobutyricum Protects against LPS-Induced Colonic Inflammation via IL-22 Signaling in Mice. Nutrients 2021, 13, 215. [Google Scholar]
- Shao, L.; Xiong, X.; Zhang, Y.; Miao, H.; Ren, Y.; Tang, X.; Song, J.; Wang, C. IL-22 ameliorates LPS-induced acute liver injury by autophagy activation through ATF4-ATG7 signaling. Cell Death Dis. 2020, 11, 970. [Google Scholar] [CrossRef] [PubMed]
- Muhl, H.; Bachmann, M. IL-18/IL-18BP and IL-22/IL-22BP: Two interrelated couples with therapeutic potential. Cell Signal 2019, 63, 109388. [Google Scholar] [PubMed]
- Zenewicz, L.A. IL-22 Binding Protein (IL-22BP) in the Regulation of IL-22 Biology. Front. Immunol. 2021, 12, 766586. [Google Scholar] [CrossRef] [PubMed]
- Raza, S.H.A.; Liang, C.; Guohua, W.; Pant, S.D.; Mohammedsaleh, Z.M.; Shater, A.F.; Alotaibi, M.A.; Khan, R.; Schreurs, N.; Cheng, G.; et al. Screening and Identification of Muscle-Specific Candidate Genes via Mouse Microarray Data Analysis. Front Vet. Sci. 2021, 8, 794628. [Google Scholar] [CrossRef] [PubMed]
- Ganieva, U.; Schneiderman, S.; Bu, P.; Beaman, K.; Dambaeva, S. IL-22 regulates endometrial regeneration by enhancing tight junctions and orchestrating extracellular matrix. Front. Immunol. 2022, 13, 955576. [Google Scholar]
- Magne, F.; Gotteland, M.; Gauthier, L.; Zazueta, A.; Pesoa, S.; Navarrete, P.; Balamurugan, R. The Firmicutes/Bacteroidetes Ratio: A Relevant Marker of Gut Dysbiosis in Obese Patients. Nutrients 2020, 12, 1474. [Google Scholar] [CrossRef]
- Bron, S.; Bolhuis, A.; Tjalsma, H.; Holsappel, S.; Venema, G.; Van Dijl, J.M. Protein secretion and possible roles for multiple signal peptidases for precursor processing in bacilli. J. Biotechnol. 1998, 64, 3–13. [Google Scholar]
- Jezewska, F.J.; Seroczynska, K.; Banaszczyk, J.; Woźniak, D.; Żylicz-Stachula, A.; Skowron, P.M. The promises and risks of probiotic Bacillus species. Acta Biochim. Pol. 2018, 65, 509–519. [Google Scholar]
- Gibson, G.R.; Roberfroid, M.B. Dietary modulation of the human colonic microbiota: Introducing the concept of prebiotics. J. Nutr. 1995, 125, 1401–1412. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Liu, Y.; Wang, Y.; Chen, X.; Wang, C.; Chen, X.; Yuan, X.; Liu, L.; Yang, J.; Zhou, X. Prevotellaceae produces butyrate to alleviate PD-1/PD-L1 inhibitor-related cardiotoxicity via PPARalpha-CYP4X1 axis in colonic macrophages. J. Exp. Clin. Cancer Res. 2022, 41, 1. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.K.; Freedman, S.N.; Murra, A.C.; Zarei, K.; Sompallae, R.; Gibson-Corley, K.N.; Karandikar, N.J.; Murray, J.A.; Mangalam, A.K. Prevotella histicola, A Human Gut Commensal, Is as Potent as COPAXONE(R) in an Animal Model of Multiple Sclerosis. Front. Immunol. 2019, 10, 462. [Google Scholar] [CrossRef] [PubMed]
- Shahi, S.K.; Jensen, S.N.; Murra, A.C.; Tang, N.; Guo, H.; Gibson-Corley, K.N.; Zhang, J.; Karandikar, N.J.; Murray, J.A.; Mangalam, A.K. Human Commensal Prevotella histicola Ameliorates Disease as Effectively as Interferon-Beta in the Experimental Autoimmune Encephalomyelitis. Front. Immunol. 2020, 11, 578648. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, M.; Zhang, P.; Fan, S.; Huang, J.; Yu, S.; Zhang, C.; Li, H. Fucoidan and galactooligosaccharides ameliorate high-fat diet-induced dyslipidemia in rats by modulating the gut microbiota and bile acid metabolism. Nutrition 2019, 65, 50–59. [Google Scholar] [CrossRef]
- Taylor, J.A.; Sichel, S.R.; Salama, N.R. Bent Bacteria: A Comparison of Cell Shape Mechanisms in Proteobacteria. Annu. Rev. Microbiol. 2019, 73, 457–480. [Google Scholar] [CrossRef]
- Rozwandowicz, M.; Brouwer, M.S.M.; Fischer, J.; A Wagenaar, J.; Gonzalez-Zorn, B.; Guerra, B.; Mevius, D.J.; Hordijk, J. Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 2018, 73, 1121–1137. [Google Scholar] [CrossRef]
- Zhao, J.; Bai, M.; Ning, X.; Qin, Y.; Wang, Y.; Yu, Z.; Dong, R.; Zhang, Y.; Sun, S. Expansion of Escherichia-Shigella in Gut Is Associated with the Onset and Response to Immunosuppressive Therapy of IgA Nephropathy. J. Am. Soc. Nephrol. 2022, 33, 2276–2292. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Size | Primer Sequence (5′-3′) |
|---|---|---|
| GAPDH-F-Cell | 208 bp | TGTTGTGGATCTGACCTGCC |
| GAPDH-R-Cell | AAGTCGCAGGAGACAACCTG | |
| GAPDH-F-Mice | 130 bp | AGGTCGGTGTGAACGGATTTG |
| GAPDH-R-Mice | GGGGTCGTTGATGGCAACA | |
| TNF-α-F-Cell | 126 bp | GTTCTCCCCATGACACCACCTG |
| TNF-α-R-Cell | GGGAGAAGAGAGTCAGACAGGC | |
| IL-6-F-Cell | 134 bp | ACAAGCGCCTTCACTCCATT |
| IL-6-R-Cell | AGAAGTAGTCTGCCTGGGGT | |
| IL-1β-F-Cell | 140 bp | ATGGCAACCGTACCTGAACC |
| IL-1β-R-Cell | CCATCTCCCATGGAACCGAG | |
| TNF-α-F-Mice | 126 bp | CCACCACGCTCTTCTGTCTA |
| TNF-α-R-Mice | CCACTTGGTGGTTTGTGAGTG | |
| IL-6-F-Mice | 138 bp | GGTCTTCTGGAGTACCATAGC |
| IL-6-R-Mice | GTGACTCCAGCTTATCTCTTGGT | |
| IL-1β-F-Mice | 136 bp | TGCCACCTTTTGACAGTGATG |
| IL-1β-R-Mice | ATGTGCTGCTGCGAGATTTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Riqing, D.; Ma, L.; Jiang, M.; Zhuoma, C.; Li, X.; Liu, Y. In Situ Expression of Yak IL-22 in Mammary Glands as a Treatment for Bovine Staphylococcus aureus-Induced Mastitis in Mice. Vet. Sci. 2024, 11, 515. https://doi.org/10.3390/vetsci11100515
Wang Z, Riqing D, Ma L, Jiang M, Zhuoma C, Li X, Liu Y. In Situ Expression of Yak IL-22 in Mammary Glands as a Treatment for Bovine Staphylococcus aureus-Induced Mastitis in Mice. Veterinary Sciences. 2024; 11(10):515. https://doi.org/10.3390/vetsci11100515
Chicago/Turabian StyleWang, Zening, Daojie Riqing, Liangliang Ma, Mingfeng Jiang, Ciren Zhuoma, Xiaowei Li, and Yili Liu. 2024. "In Situ Expression of Yak IL-22 in Mammary Glands as a Treatment for Bovine Staphylococcus aureus-Induced Mastitis in Mice" Veterinary Sciences 11, no. 10: 515. https://doi.org/10.3390/vetsci11100515
APA StyleWang, Z., Riqing, D., Ma, L., Jiang, M., Zhuoma, C., Li, X., & Liu, Y. (2024). In Situ Expression of Yak IL-22 in Mammary Glands as a Treatment for Bovine Staphylococcus aureus-Induced Mastitis in Mice. Veterinary Sciences, 11(10), 515. https://doi.org/10.3390/vetsci11100515