
Intra- and Intertumoral Microglia/Macrophage Infiltration and Their Associated Molecular Signature Is Highly Variable in Canine Oligodendroglioma: A Preliminary Evaluation

 , , , ,
, , , ,
Abstract
Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. Image Acquisition and Analysis
2.2.1. Immunofluorescence and Confocal Microscopy
2.2.2. Microglia Quantification
2.3. Canine Protein Arrays
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
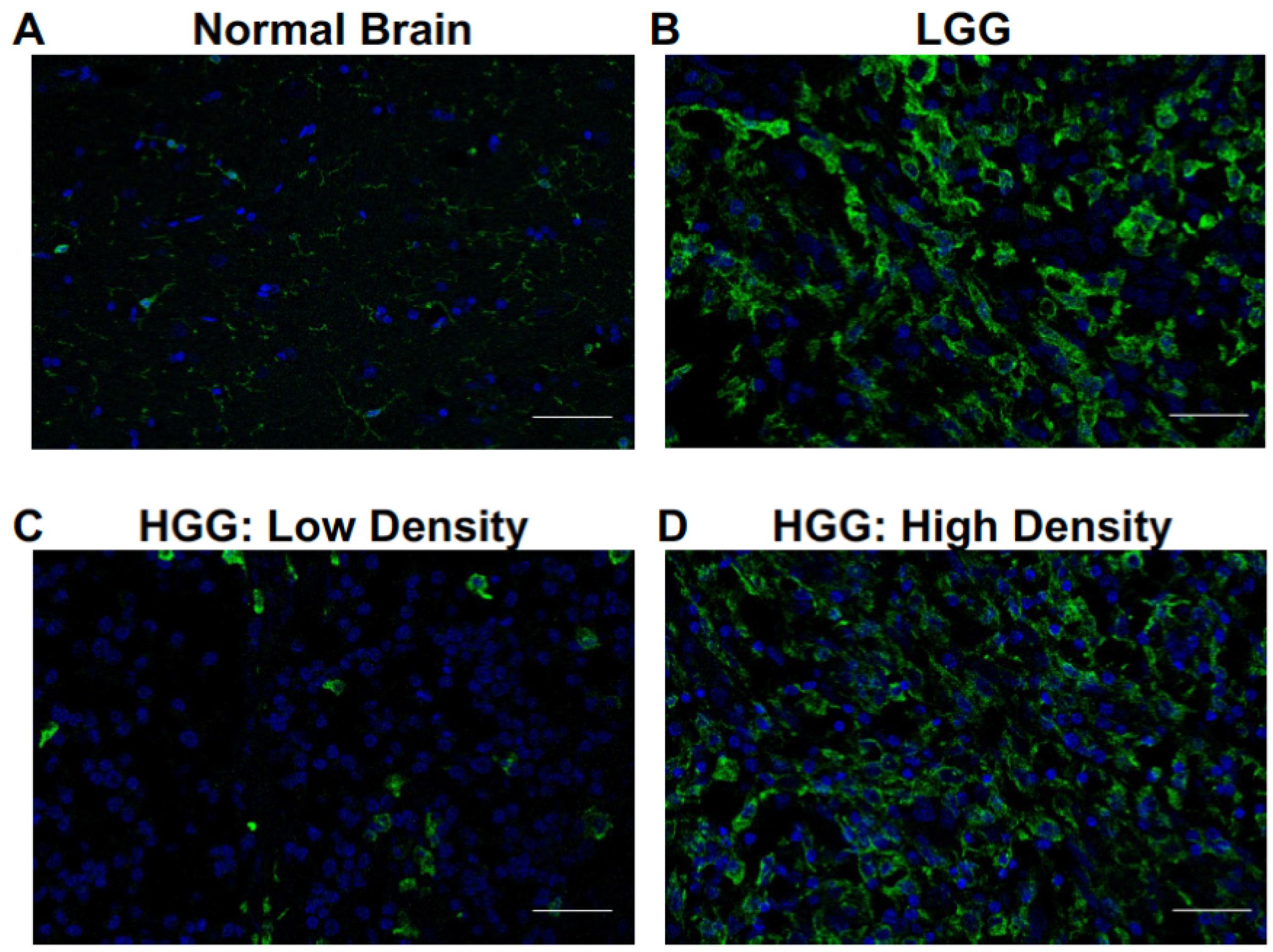
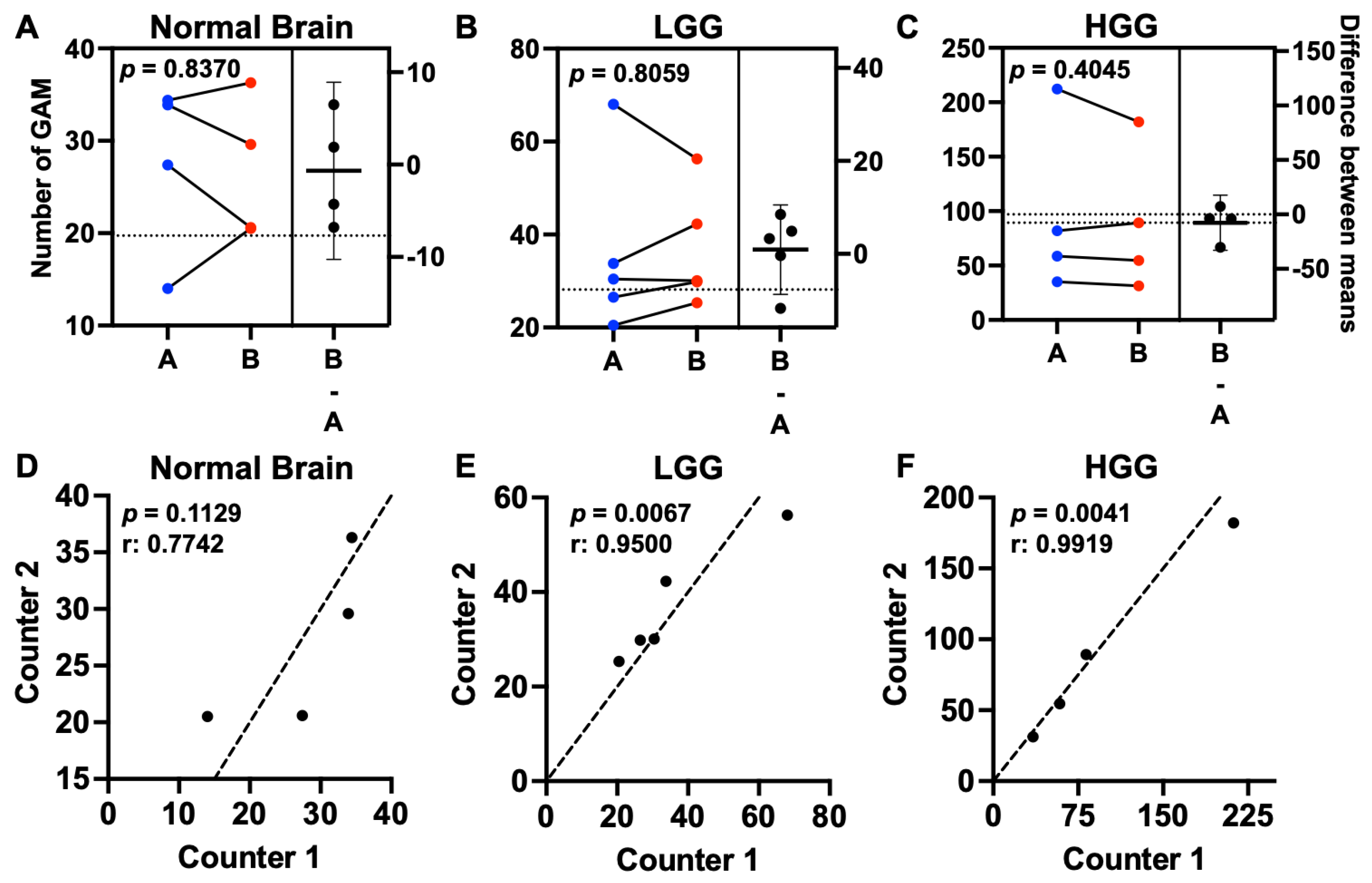
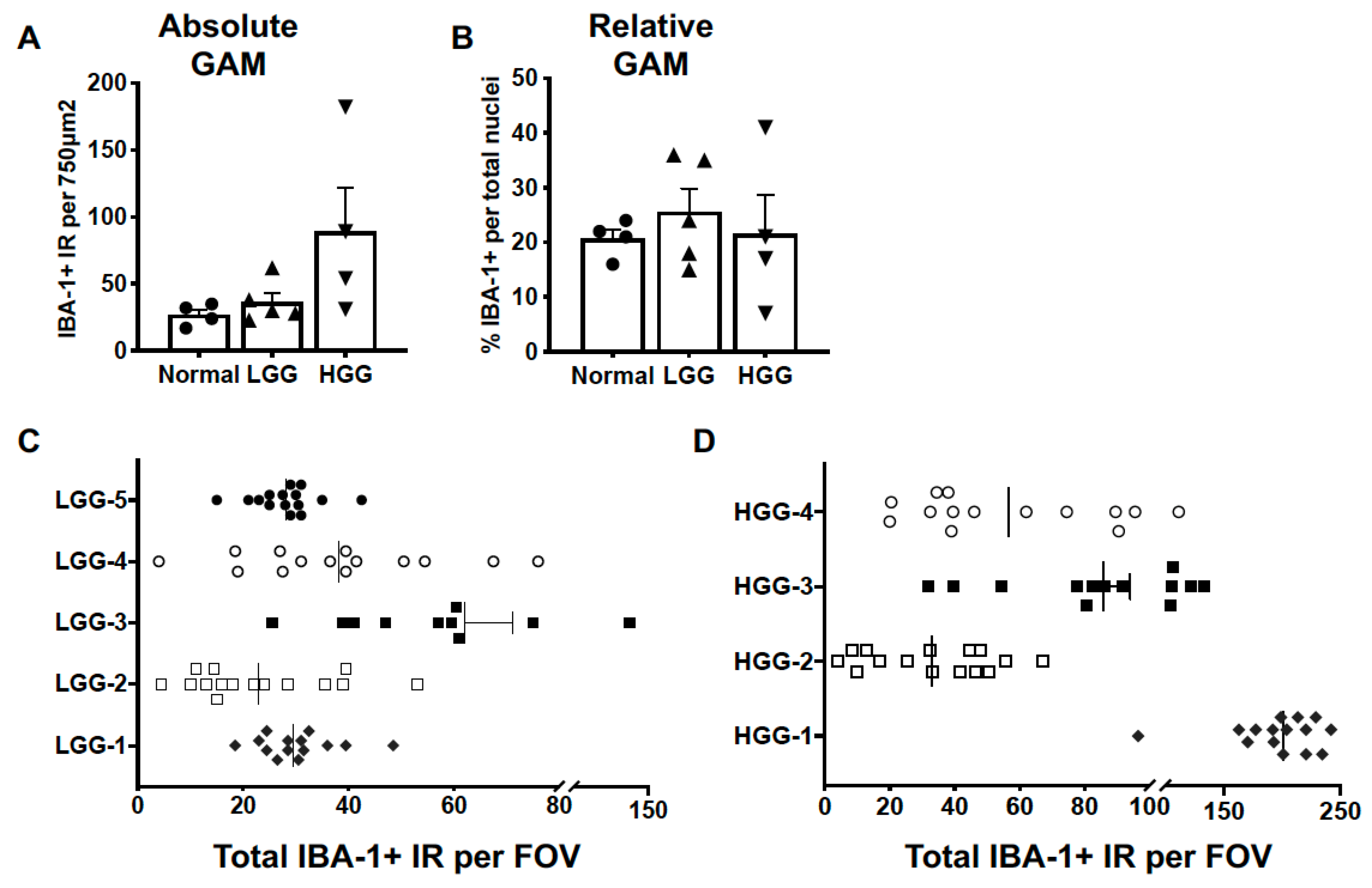
3.1. Microglia and Macrophages Heterogeneously Infiltrate Canine Oligodendrogliomas
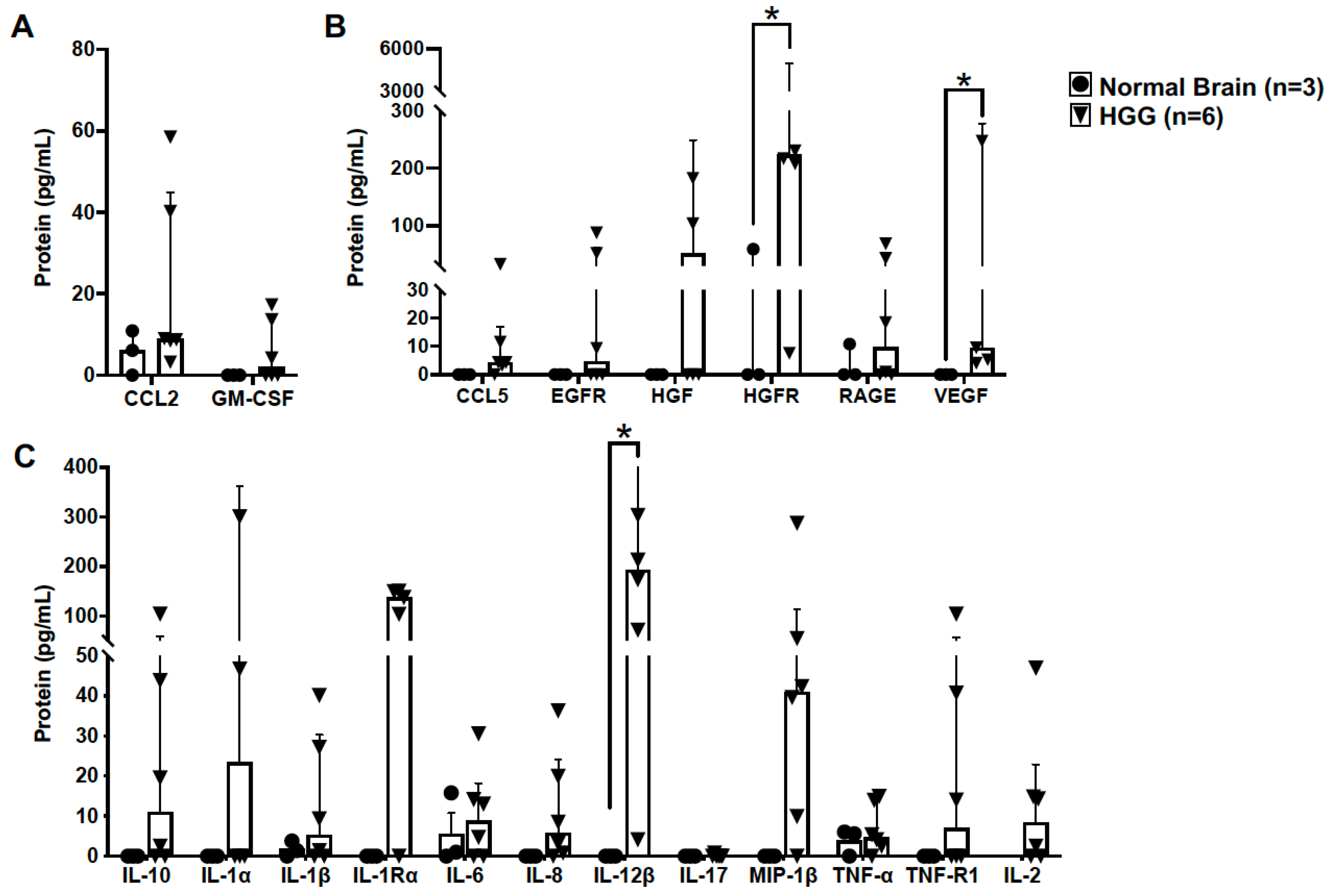
3.2. The Molecular Landscape of Canine High-Grade Oligodendrogliomas Exhibits Increased Expression of Proteins Associated with a Pro-Tumorigenic GAM Signature
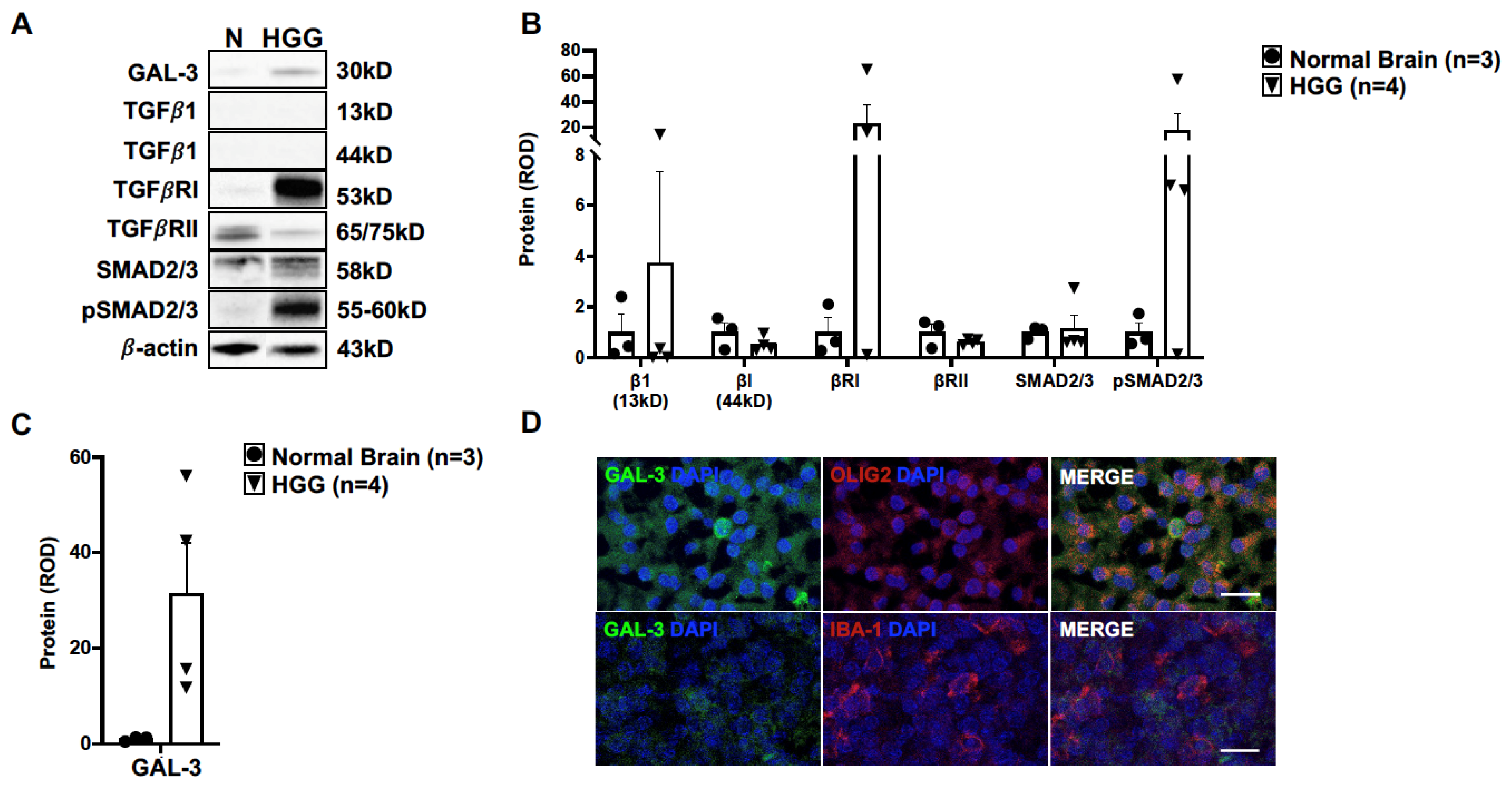
3.3. TGFβ and Galectin-3 May Contribute to Canine Oligodendroglioma Pathogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Song, R.B.; Vite, C.H.; Bradley, C.W.; Cross, J.R. Postmortem Evaluation of 435 Cases of Intracranial Neoplasia in Dogs and Relationship of Neoplasm with Breed, Age, and Body Weight. J. Vet. Intern. Med. 2013, 27, 1143–1152. [Google Scholar] [CrossRef] [PubMed]
- Truve, K.; Dickinson, P.; Xiong, A.; York, D.; Jayashankar, K.; Pielberg, G.; Koltookian, M.; Muren, E.; Fuxelius, H.H.; Weishaupt, H.; et al. Utilizing the Dog Genome in the Search for Novel Candidate Genes Involved in Glioma Development-Genome Wide Association Mapping followed by Targeted Massive Parallel Sequencing Identifies a Strongly Associated Locus. PLoS Genet. 2016, 12, e1006000. [Google Scholar] [CrossRef] [PubMed]
- Mariani, C.L.; Schubert, T.A.; House, R.A.; Wong, M.A.; Hopkins, A.L.; Barnes Heller, H.L.; Milner, R.J.; Lester, N.V.; Lurie, D.M.; Rajon, D.A.; et al. Frameless stereotactic radiosurgery for the treatment of primary intracranial tumours in dogs. Vet. Comp. Oncol. 2015, 13, 409–423. [Google Scholar] [CrossRef] [PubMed]
- Brearley, M.J.; Jeffery, N.D.; Phillips, S.M.; Dennis, R. Hypofractionated radiation therapy of brain masses in dogs: A retrospective analysis of survival of 83 cases (1991–1996). J. Vet. Intern. Med. Am. Coll. Vet. Intern. Med. 1999, 13, 408–412. [Google Scholar]
- MacLellan, J.D.; Arnold, S.A.; Dave, A.C.; Hunt, M.A.; Pluhar, G.E. Association of magnetic resonance imaging-based preoperative tumor volume with postsurgical survival time in dogs with primary intracranial glioma. J. Am. Vet. Med. Assoc. 2018, 252, 98–102. [Google Scholar] [CrossRef]
- Amin, S.B.; Anderson, K.J.; Boudreau, C.E.; Martinez-Ledesma, E.; Kocakavuk, E.; Johnson, K.C.; Barthel, F.P.; Varn, F.S.; Kassab, C.; Ling, X.; et al. Comparative Molecular Life History of Spontaneous Canine and Human Gliomas. Cancer Cell 2020, 37, 243–257.e7. [Google Scholar] [CrossRef]
- Wu, A.; Wei, J.; Kong, L.Y.; Wang, Y.; Priebe, W.; Qiao, W.; Sawaya, R.; Heimberger, A.B. Glioma cancer stem cells induce immunosuppressive macrophages/microglia. Neuro-Oncol. 2010, 12, 1113–1125. [Google Scholar] [CrossRef]
- Krane, G.A.; O’Dea, C.A.; Malarkey, D.E.; Miller, A.D.; Miller, C.R.; Tokarz, D.A.; Jensen, H.L.; Janardhan, K.S.; Shockley, K.R.; Flagler, N.; et al. Immunohistochemical evaluation of immune cell infiltration in canine gliomas. Vet. Pathol. 2021, 58, 952–963. [Google Scholar] [CrossRef]
- Sloma, E.A.; Creneti, C.T.; Erb, H.N.; Miller, A.D. Characterization of Inflammatory Changes Associated with Canine Oligodendroglioma. J. Comp. Pathol. 2015, 153, 92–100. [Google Scholar] [CrossRef]
- Flores-Toro, J.A.; Luo, D.; Gopinath, A.; Sarkisian, M.R.; Campbell, J.J.; Charo, I.F.; Singh, R.; Schall, T.J.; Datta, M.; Jain, R.K.; et al. CCR2 inhibition reduces tumor myeloid cells and unmasks a checkpoint inhibitor effect to slow progression of resistant murine gliomas. Proc. Natl. Acad. Sci. USA 2020, 117, 1129–1138. [Google Scholar] [CrossRef]
- Platten, M.; Kretz, A.; Naumann, U.; Aulwurm, S.; Egashira, K.; Isenmann, S.; Weller, M. Monocyte chemoattractant protein-1 increases microglial infiltration and aggressiveness of gliomas. Ann. Neurol. 2003, 54, 388–392. [Google Scholar] [CrossRef]
- Sorensen, M.D.; Dahlrot, R.H.; Boldt, H.B.; Hansen, S.; Kristensen, B.W. Tumour-associated microglia/macrophages predict poor prognosis in high-grade gliomas and correlate with an aggressive tumour subtype. Neuropathol. Appl. Neurobiol. 2018, 44, 185–206. [Google Scholar] [CrossRef]
- Herting, C.J.; Chen, Z.; Maximov, V.; Duffy, A.; Szulzewsky, F.; Shayakhmetov, D.M.; Hambardzumyan, D. Tumour-associated macrophage-derived interleukin-1 mediates glioblastoma-associated cerebral oedema. Brain J. Neurol. 2019, 142, 3834–3851. [Google Scholar] [CrossRef]
- Blank, A.; Kremenetskaia, I.; Urbantat, R.M.; Acker, G.; Turkowski, K.; Radke, J.; Schneider, U.C.; Vajkoczy, P.; Brandenburg, S. Microglia/macrophages express alternative proangiogenic factors depending on granulocyte content in human glioblastoma. J. Pathol. 2021, 253, 160–173. [Google Scholar] [CrossRef] [PubMed]
- Toedebusch, R.; Grodzki, A.C.; Dickinson, P.J.; Woolard, K.; Vinson, N.; Sturges, B.; Snyder, J.; Li, C.-F.; Nagasaka, O.; Consales, B.; et al. Glioma-associated microglia/macrophages augment tumorigenicity in canine astrocytoma, a naturally occurring model of human glioma. Neuro-Oncol. Adv. 2021, 3, vdab062. [Google Scholar] [CrossRef] [PubMed]
- Kjellman, C.; Olofsson, S.P.; Hansson, O.; Von Schantz, T.; Lindvall, M.; Nilsson, I.; Salford, L.G.; Sjogren, H.O.; Widegren, B. Expression of TGF-beta isoforms, TGF-beta receptors, and SMAD molecules at different stages of human glioma. Int. J. Cancer 2000, 89, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Bodmer, S.; Strommer, K.; Frei, K.; Siepl, C.; de Tribolet, N.; Heid, I.; Fontana, A. Immunosuppression and transforming growth factor-beta in glioblastoma. Preferential production of transforming growth factor-beta 2. J. Immunol. 1989, 143, 3222–3229. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.Z.; Xu, S.L.; Xin, Y.H.; Yu, S.C.; Ping, Y.F.; Chen, L.; Xiao, H.L.; Wang, B.; Yi, L.; Wang, Q.L.; et al. Tumor-associated microglia/macrophages enhance the invasion of glioma stem-like cells via TGF-β1 signaling pathway. J. Immunol. 2012, 189, 444–453. [Google Scholar] [CrossRef]
- Wesolowska, A.; Kwiatkowska, A.; Slomnicki, L.; Dembinski, M.; Master, A.; Sliwa, M.; Franciszkiewicz, K.; Chouaib, S.; Kaminska, B. Microglia-derived TGF-beta as an important regulator of glioblastoma invasion—An inhibition of TGF-beta-dependent effects by shRNA against human TGF-beta type II receptor. Oncogene 2008, 27, 918–930. [Google Scholar] [CrossRef]
- Toedebusch, R.G.; Lucchesi, C.A.; Debebe, E.T.; Wittenburg, L.A.; Chen, X.; Toedebusch, C.M. Microglia-Derived Olfactomedin-like 3 Promotes Pro-Tumorigenic Microglial Function and Malignant Features of Glioma Cells. Int. J. Mol. Sci. 2021, 22, 13052. [Google Scholar] [CrossRef]
- Koehler, J.W.; Miller, A.D.; Miller, C.R.; Porter, B.; Aldape, K.; Beck, J.; Brat, D.; Cornax, I.; Corps, K.; Frank, C.; et al. A Revised Diagnostic Classification of Canine Glioma: Towards Validation of the Canine Glioma Patient as a Naturally Occurring Preclinical Model for Human Glioma. J. Neuropathol. Exp. Neurol. 2018, 77, 1039–1054. [Google Scholar] [CrossRef] [PubMed]
- Louis, D.N.; Perry, A.; Wesseling, P.; Brat, D.J.; Cree, I.A.; Figarella-Branger, D.; Hawkins, C.; Ng, H.K.; Pfister, S.M.; Reifenberger, G.; et al. The 2021 WHO Classification of Tumors of the Central Nervous System: A summary. Neuro-Oncol. 2021, 23, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Lum-Naihe, K.; Toedebusch, R.; Mahmood, A.; Bajwa, J.; Carmack, T.; Kumar, S.A.; Ardhanari, S.; DeMarco, V.G.; Emter, C.A.; Pulakat, L. Cardiovascular disease progression in female Zucker Diabetic Fatty rats occurs via unique mechanisms compared to males. Sci. Rep. 2017, 7, 17823. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, A.E.; Garcia-Bueno, B.; Leza, J.C.; Madrigal, J.L. CCL2/MCP-1 modulation of microglial activation and proliferation. J. Neuroinflamm. 2011, 8, 77. [Google Scholar] [CrossRef]
- Chang, A.L.; Miska, J.; Wainwright, D.A.; Dey, M.; Rivetta, C.V.; Yu, D.; Kanojia, D.; Pituch, K.C.; Qiao, J.; Pytel, P.; et al. CCL2 Produced by the Glioma Microenvironment Is Essential for the Recruitment of Regulatory T Cells and Myeloid-Derived Suppressor Cells. Cancer Res. 2016, 76, 5671–5682. [Google Scholar] [CrossRef] [PubMed]
- Moransard, M.; Sawitzky, M.; Fontana, A.; Suter, T. Expression of the HGF receptor c-met by macrophages in experimental autoimmune encephalomyelitis. Glia 2010, 58, 559–571. [Google Scholar] [CrossRef]
- Petterson, S.A.; Dahlrot, R.H.; Hermansen, S.K.; KA Munthe, S.; Gundesen, M.T.; Wohlleben, H.; Rasmussen, T.; Beier, C.P.; Hansen, S.; Kristensen, B.W. High levels of c-Met is associated with poor prognosis in glioblastoma. J. Neuro-Oncol. 2015, 122, 517–527. [Google Scholar] [CrossRef]
- Tian, Y.; Yuan, W.; Li, J.; Wang, H.; Hunt, M.G.; Liu, C.; Shapiro, I.M.; Risbud, M.V. TGFβ regulates Galectin-3 expression through canonical Smad3 signaling pathway in nucleus pulposus cells: Implications in intervertebral disc degeneration. Matrix Biol. 2016, 50, 39–52. [Google Scholar] [CrossRef]
- Mackinnon, A.C.; Gibbons, M.A.; Farnworth, S.L.; Leffler, H.; Nilsson, U.J.; Delaine, T.; Simpson, A.J.; Forbes, S.J.; Hirani, N.; Gauldie, J.; et al. Regulation of transforming growth factor-β1-driven lung fibrosis by galectin-3. Am. J. Respir. Crit. Care Med. 2012, 185, 537–546. [Google Scholar] [CrossRef]
- Dickinson, P.J.; Sturges, B.K.; Higgins, R.J.; Roberts, B.N.; Leutenegger, C.M.; Bollen, A.W.; LeCouteur, R.A. Vascular endothelial growth factor mRNA expression and peritumoral edema in canine primary central nervous system tumors. Vet. Pathol. 2008, 45, 131–139. [Google Scholar] [CrossRef]
- Rossmeisl, J.H.; Duncan, R.B.; Huckle, W.R.; Troy, G.C. Expression of vascular endothelial growth factor in tumors and plasma from dogs with primary intracranial neoplasms. Am. J. Vet. Res. 2007, 68, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Dickinson, P.J.; Roberts, B.N.; Higgins, R.J.; Leutenegger, C.M.; Bollen, A.W.; Kass, P.H.; LeCouteur, R.A. Expression of receptor tyrosine kinases VEGFR-1 (FLT-1), VEGFR-2 (KDR), EGFR-1, PDGFRalpha and c-Met in canine primary brain tumours. Vet. Comp. Oncol. 2006, 4, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Mariani, C.L.; Niman, Z.E.; Boozer, L.B.; Ruterbories, L.K.; Early, P.J.; Muñana, K.R.; Olby, N.J. Vascular endothelial growth factor concentrations in the cerebrospinal fluid of dogs with neoplastic or inflammatory central nervous system disorders. J. Vet. Intern. Med. Am. Coll. Vet. Intern. Med. 2021, 35, 1873–1883. [Google Scholar] [CrossRef]
- Lin, G.L.; Nagaraja, S.; Filbin, M.G.; Suva, M.L.; Vogel, H.; Monje, M. Non-inflammatory tumor microenvironment of diffuse intrinsic pontine glioma. Acta Neuropathol. Commun. 2018, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Seol, H.J.; Kim, E.H.; Rheey, J.; Jin, H.J.; Lee, Y.; Joo, K.M.; Lee, J.; Nam, D.H. Wnt/β-catenin signaling is a key downstream mediator of MET signaling in glioblastoma stem cells. Neuro-Oncol. 2013, 15, 161–171. [Google Scholar] [CrossRef]
- Dong, F.; Eibach, M.; Bartsch, J.W.; Dolga, A.M.; Schlomann, U.; Conrad, C.; Schieber, S.; Schilling, O.; Biniossek, M.L.; Culmsee, C.; et al. The metalloprotease-disintegrin ADAM8 contributes to temozolomide chemoresistance and enhanced invasiveness of human glioblastoma cells. Neuro-Oncol. 2015, 17, 1474–1485. [Google Scholar] [CrossRef] [PubMed]
- Recurrent MET fusion genes represent a drug target in pediatric glioblastoma. Nat. Med. 2016, 22, 1314–1320. [CrossRef]
- Wu, G.; Diaz, A.K.; Paugh, B.S.; Rankin, S.L.; Ju, B.; Li, Y.; Zhu, X.; Qu, C.; Chen, X.; Zhang, J.; et al. The genomic landscape of diffuse intrinsic pontine glioma and pediatric non-brainstem high-grade glioma. Nat. Genet. 2014, 46, 444–450. [Google Scholar] [CrossRef]
- Wen, P.Y.; Schiff, D.; Cloughesy, T.F.; Raizer, J.J.; Laterra, J.; Smitt, M.; Wolf, M.; Oliner, K.S.; Anderson, A.; Zhu, M.; et al. A phase II study evaluating the efficacy and safety of AMG 102 (rilotumumab) in patients with recurrent glioblastoma. Neuro-Oncol. 2011, 13, 437–446. [Google Scholar] [CrossRef]
- Affronti, M.L.; Jackman, J.G.; McSherry, F.; Herndon, J.E., 2nd; Massey, E.C., Jr.; Lipp, E.; Desjardins, A.; Friedman, H.S.; Vlahovic, G.; Vredenburgh, J.; et al. Phase II Study to Evaluate the Efficacy and Safety of Rilotumumab and Bevacizumab in Subjects with Recurrent Malignant Glioma. Oncologist 2018, 23, 889-e98. [Google Scholar] [CrossRef]
- Cloughesy, T.; Finocchiaro, G.; Belda-Iniesta, C.; Recht, L.; Brandes, A.A.; Pineda, E.; Mikkelsen, T.; Chinot, O.L.; Balana, C.; Macdonald, D.R.; et al. Randomized, Double-Blind, Placebo-Controlled, Multicenter Phase II Study of Onartuzumab Plus Bevacizumab versus Placebo Plus Bevacizumab in Patients with Recurrent Glioblastoma: Efficacy, Safety, and Hepatocyte Growth Factor and O(6)-Methylguanine-DNA Methyltransferase Biomarker Analyses. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2017, 35, 343–351. [Google Scholar] [CrossRef]
- Ajarrag, S.; St-Pierre, Y. Galectins in Glioma: Current Roles in Cancer Progression and Future Directions for Improving Treatment. Cancers 2021, 13, 5533. [Google Scholar] [CrossRef] [PubMed]
- MacKinnon, A.C.; Farnworth, S.L.; Hodkinson, P.S.; Henderson, N.C.; Atkinson, K.M.; Leffler, H.; Nilsson, U.J.; Haslett, C.; Forbes, S.J.; Sethi, T. Regulation of alternative macrophage activation by galectin-3. J. Immunol. 2008, 180, 2650–2658. [Google Scholar] [CrossRef]
- Lalancette-Hébert, M.; Gowing, G.; Simard, A.; Yuan, C.W.; Kriz, J. Selective ablation of proliferating microglial cells exacerbates ischemic injury in the brain. J. Neurosci. 2007, 27, 2596–2605. [Google Scholar] [CrossRef]
- Binh, N.H.; Satoh, K.; Kobayashi, K.; Takamatsu, M.; Hatano, Y.; Hirata, A.; Tomita, H.; Kuno, T.; Hara, A. Galectin-3 in preneoplastic lesions of glioma. J. Neuro-Oncol. 2013, 111, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Paixao Becker, A.; de Oliveira, R.S.; Saggioro, F.P.; Neder, L.; Chimelli, L.M.; Machado, H.R. In pursuit of prognostic factors in children with pilocytic astrocytomas. Childs Nerv. Syst. ChNS Off. J. Int. Soc. Pediatr. Neurosurg. 2010, 26, 19–28. [Google Scholar] [CrossRef]
- Fortuna-Costa, A.; Gomes, A.M.; Kozlowski, E.O.; Stelling, M.P.; Pavão, M.S. Extracellular galectin-3 in tumor progression and metastasis. Front. Oncol. 2014, 4, 138. [Google Scholar] [CrossRef]
- Nangia-Makker, P.; Balan, V.; Raz, A. Regulation of tumor progression by extracellular galectin-3. Cancer Microenviron. 2008, 1, 43–51. [Google Scholar] [CrossRef]
- Chen, A.; Jiang, Y.; Li, Z.; Wu, L.; Santiago, U.; Zou, H.; Cai, C.; Sharma, V.; Guan, Y.; McCarl, L.H.; et al. Chitinase-3-like 1 protein complexes modulate macrophage-mediated immune suppression in glioblastoma. J. Clin. Investig. 2021, 131, e147552. [Google Scholar] [CrossRef]
- Gutmann, D.H.; McLellan, M.D.; Hussain, I.; Wallis, J.W.; Fulton, L.L.; Fulton, R.S.; Magrini, V.; Demeter, R.; Wylie, T.; Kandoth, C.; et al. Somatic neurofibromatosis type 1 (NF1) inactivation characterizes NF1-associated pilocytic astrocytoma. Genome Res. 2013, 23, 431–439. [Google Scholar] [CrossRef]
- Markovic, D.S.; Glass, R.; Synowitz, M.; Rooijen, N.; Kettenmann, H. Microglia stimulate the invasiveness of glioma cells by increasing the activity of metalloprotease-2. J. Neuropathol. Exp. Neurol. 2005, 64, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Kowal, J.; Akkari, L.; Schuhmacher, A.J.; Huse, J.T.; West, B.L.; Joyce, J.A. Inhibition of colony stimulating factor-1 receptor abrogates microenvironment-mediated therapeutic resistance in gliomas. Oncogene 2017, 36, 6049–6058. [Google Scholar] [CrossRef] [PubMed]
- Fathima Hurmath, K.; Ramaswamy, P.; Nandakumar, D.N. IL-1β microenvironment promotes proliferation, migration, and invasion of human glioma cells. Cell Biol. Int. 2014, 38, 1415–1422. [Google Scholar] [CrossRef]
- Conroy, S.; Kruyt, F.A.E.; Wagemakers, M.; Bhat, K.P.L.; den Dunnen, W.F.A. IL-8 associates with a pro-angiogenic and mesenchymal subtype in glioblastoma. Oncotarget 2018, 9, 15721–15731. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Huang, X.; Li, J.; Fan, H.; Yang, F.; Zhang, R.; Yang, Y.; Feng, S.; He, D.; Sun, W.; et al. Interleukin 10 promotes growth and invasion of glioma cells by up-regulating KPNA 2 in vitro. J. Cancer Res. Ther. 2019, 15, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, T.; Yang, N.; Xu, S.; Li, X.; Wang, D. Hypoxia and macrophages promote glioblastoma invasion by the CCL4-CCR5 axis. Oncol. Rep. 2016, 36, 3522–3528. [Google Scholar] [CrossRef]
- Wu, C.Y.J.; Chen, C.H.; Lin, C.Y.; Feng, L.Y.; Lin, Y.C.; Wei, K.C.; Huang, C.Y.; Fang, J.Y.; Chen, P.Y. CCL5 of glioma-associated microglia/macrophages regulates glioma migration and invasion via calcium-dependent matrix metalloproteinase 2. Neuro-Oncology 2020, 22, 253–266. [Google Scholar] [CrossRef]
- Roesch, S.; Rapp, C.; Dettling, S.; Herold-Mende, C. When Immune Cells Turn Bad-Tumor-Associated Microglia/Macrophages in Glioma. Int. J. Mol. Sci. 2018, 19, 436. [Google Scholar] [CrossRef]
- Toedebusch, C.M.; Garcia, V.B.; Snyder, J.C.; Jones, M.R.; Schulz, D.J.; Johnson, G.C.; Villalon, E.; Coates, J.R.; Garcia, M.L. Lumbar spinal cord microglia exhibited increased activation in aging dogs compared with young adult dogs. GeroScience 2020, 42, 169–182. [Google Scholar] [CrossRef]
- Kongsui, R.; Beynon, S.B.; Johnson, S.J.; Walker, F.R. Quantitative assessment of microglial morphology and density reveals remarkable consistency in the distribution and morphology of cells within the healthy prefrontal cortex of the rat. J. Neuroinflamm. 2014, 11, 182. [Google Scholar] [CrossRef]
- Melchior, B.; Puntambekar, S.S.; Carson, M.J. Microglia and the control of autoreactive T cell responses. Neurochem. Int. 2006, 49, 145–153. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Breed | Age (years) | Sex | Tumor Location | Tissue Analysis | |
|---|---|---|---|---|---|---|
| NCI Grade | WHO Grade | |||||
| Low | Grade II | French Bulldog | 12 | FS | Right temporal lobe | IF |
| Boxer | 9 | FS | Left piriform lobe | IF | ||
| Boston Terrier | 9 | FI | Left temporoparietal lobes | IF | ||
| Boxer | 5 | MC | Left frontal lobe | IF | ||
| English Bulldog | 8 | MC | Left frontal to occipital lobes—periventricular | IF | ||
| High | Grade III | French Bulldog | 7 | FS | Right frontoparietal lobes | IF |
| French Bulldog | 11 | FS | Left temporal lobe | IF | ||
| Boxer | 9 | FS | Left cerebrum and lateral ventricle | IF | ||
| Boxer | 8 | MI | Right temporoparietal lobes | IF | ||
| Boxer | 7 | FS | White matter of left parietal, temporal, and occipital lobes | IF | ||
| Boxer | 13 | FI | Left temporal lobe | Protein Array | ||
| English Bulldog | 12 | M | Left intraventricular septum, lateral ventricle | Protein Array | ||
| English Bulldog | 9 | FS | Left frontal lobe | WB, Protein Array | ||
| Rottweiler Mix | 10 | MC | Left frontal lobe extending into caudate nucleus | WB, Protein Array | ||
| Boxer | 8 | MI | Right piriform and temporal lobes | WB, Protein Array | ||
| Boxer | 4 | MC | Lateral ventricle with broad base in caudate nucleus and periventricular white matter | WB, Protein Array | ||
| Control Cases | Breed | Age (years) | Sex | Cause of Death | Tissue Analysis | |
| French Bulldog | 3 | MC | Euthanasia: anti-coagulant toxicity; subdural and parenchymal spinal cord hemorrhage | IF | ||
| Golden Retriever | 10 | MC | Euthanasia: primary spinal cord vasculitis | IF | ||
| French Bulldog | 3 | MC | Euthanasia: ascending/descending myelomalcia following L3-L5 intervertebral disc extrusion | IF | ||
| French Bulldog | 3 | MC | Euthanasia: ascending/descending myelomalcia following L3-L4 intervertebral disc extrusion | WB, Protein Array | ||
| Beagle Mix | 7 | FS | Euthanasia: acute T3-L3 myelopathy; paraplegic, absent nociception. | WB, Protein Array | ||
| Boxer | 10 | MC | Euthanasia: prostatic mass with urethral obstruction | WB, Protein Array | ||
| Boxer | 7 | FS | Euthanasia: L4-S1 peripheral nerve sheath tumor | WB, Protein Array | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toedebusch, R.G.; Wei, N.-W.; Simafranca, K.T.; Furth-Jacobus, J.A.; Brust-Mascher, I.; Stewart, S.L.; Dickinson, P.J.; Woolard, K.D.; Li, C.-F.; Vernau, K.M.; et al. Intra- and Intertumoral Microglia/Macrophage Infiltration and Their Associated Molecular Signature Is Highly Variable in Canine Oligodendroglioma: A Preliminary Evaluation. Vet. Sci. 2023, 10, 403. https://doi.org/10.3390/vetsci10060403
Toedebusch RG, Wei N-W, Simafranca KT, Furth-Jacobus JA, Brust-Mascher I, Stewart SL, Dickinson PJ, Woolard KD, Li C-F, Vernau KM, et al. Intra- and Intertumoral Microglia/Macrophage Infiltration and Their Associated Molecular Signature Is Highly Variable in Canine Oligodendroglioma: A Preliminary Evaluation. Veterinary Sciences. 2023; 10(6):403. https://doi.org/10.3390/vetsci10060403
Chicago/Turabian StyleToedebusch, Ryan G., Ning-Wei Wei, Kulani T. Simafranca, Jennie A. Furth-Jacobus, Ingrid Brust-Mascher, Susan L. Stewart, Peter J. Dickinson, Kevin D. Woolard, Chai-Fei Li, Karen M. Vernau, and et al. 2023. "Intra- and Intertumoral Microglia/Macrophage Infiltration and Their Associated Molecular Signature Is Highly Variable in Canine Oligodendroglioma: A Preliminary Evaluation" Veterinary Sciences 10, no. 6: 403. https://doi.org/10.3390/vetsci10060403
APA StyleToedebusch, R. G., Wei, N.-W., Simafranca, K. T., Furth-Jacobus, J. A., Brust-Mascher, I., Stewart, S. L., Dickinson, P. J., Woolard, K. D., Li, C.-F., Vernau, K. M., Meyers, F. J., & Toedebusch, C. M. (2023). Intra- and Intertumoral Microglia/Macrophage Infiltration and Their Associated Molecular Signature Is Highly Variable in Canine Oligodendroglioma: A Preliminary Evaluation. Veterinary Sciences, 10(6), 403. https://doi.org/10.3390/vetsci10060403