
Mammary Glands of Women, Female Dogs and Female Rats: Similarities and Differences to Be Considered in Breast Cancer Research

 , ,
, ,  ,
,  ,
,  ,
,  and
and 
Abstract
Simple Summary
Abstract
1. Introduction
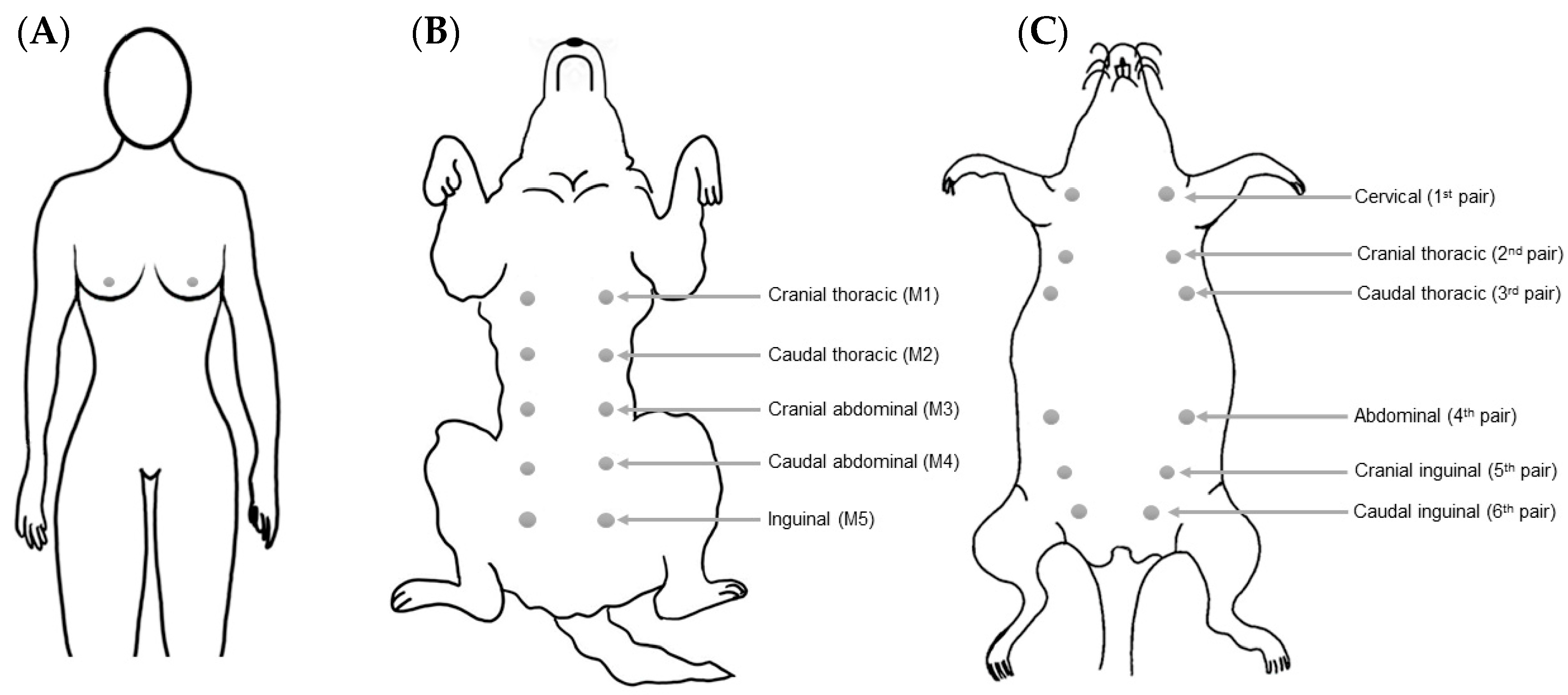
2. Breast/Mammary Gland Anatomy
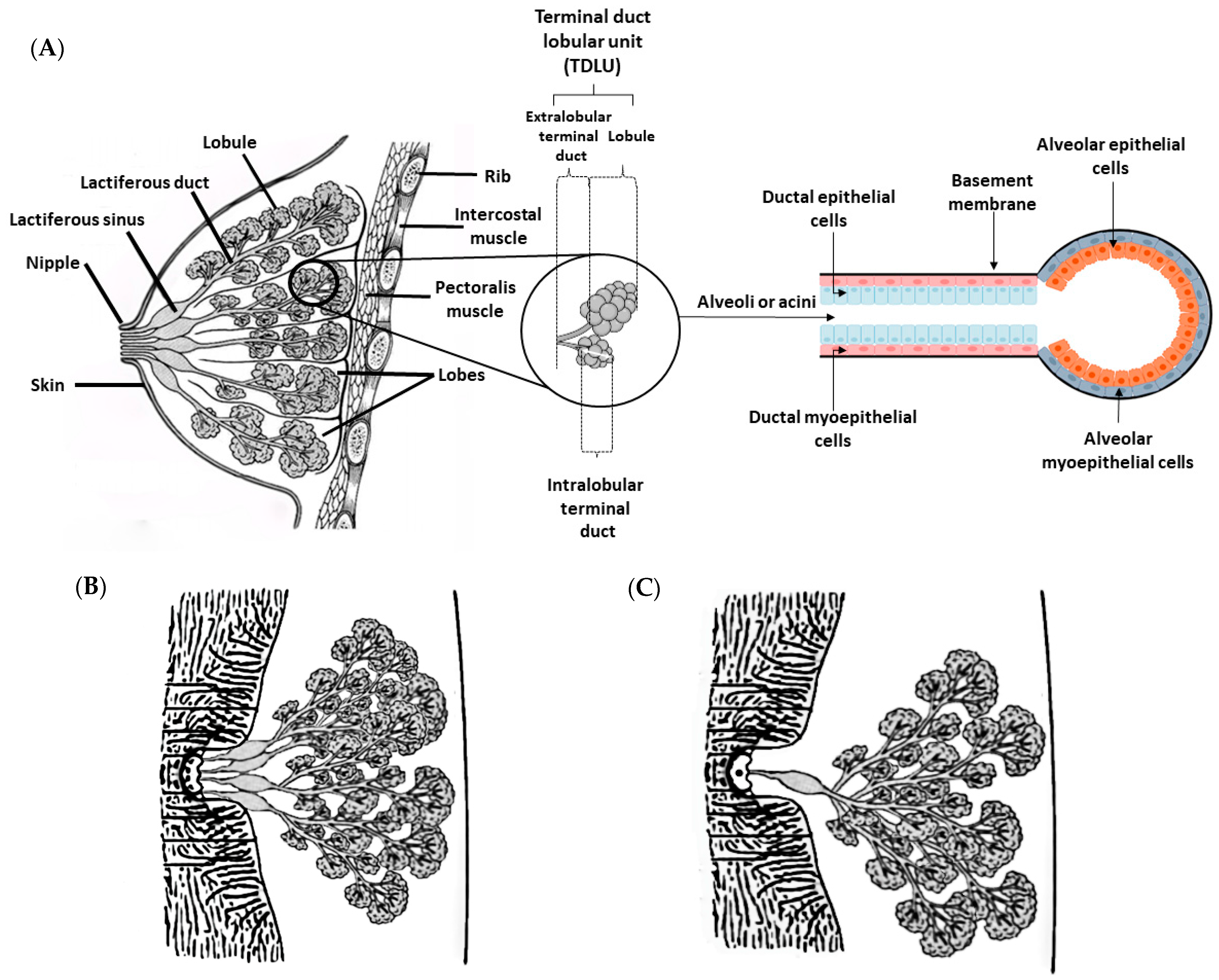
3. Physiological and Histological Features of the Mammary Gland
3.1. Supply and Venous Drainage
3.2. Lymphatic Drainage
3.3. Innervation
4. Breast/Mammary Cancer
4.1. Epidemiology
4.2. Risk Factors and Breast Cancer Development
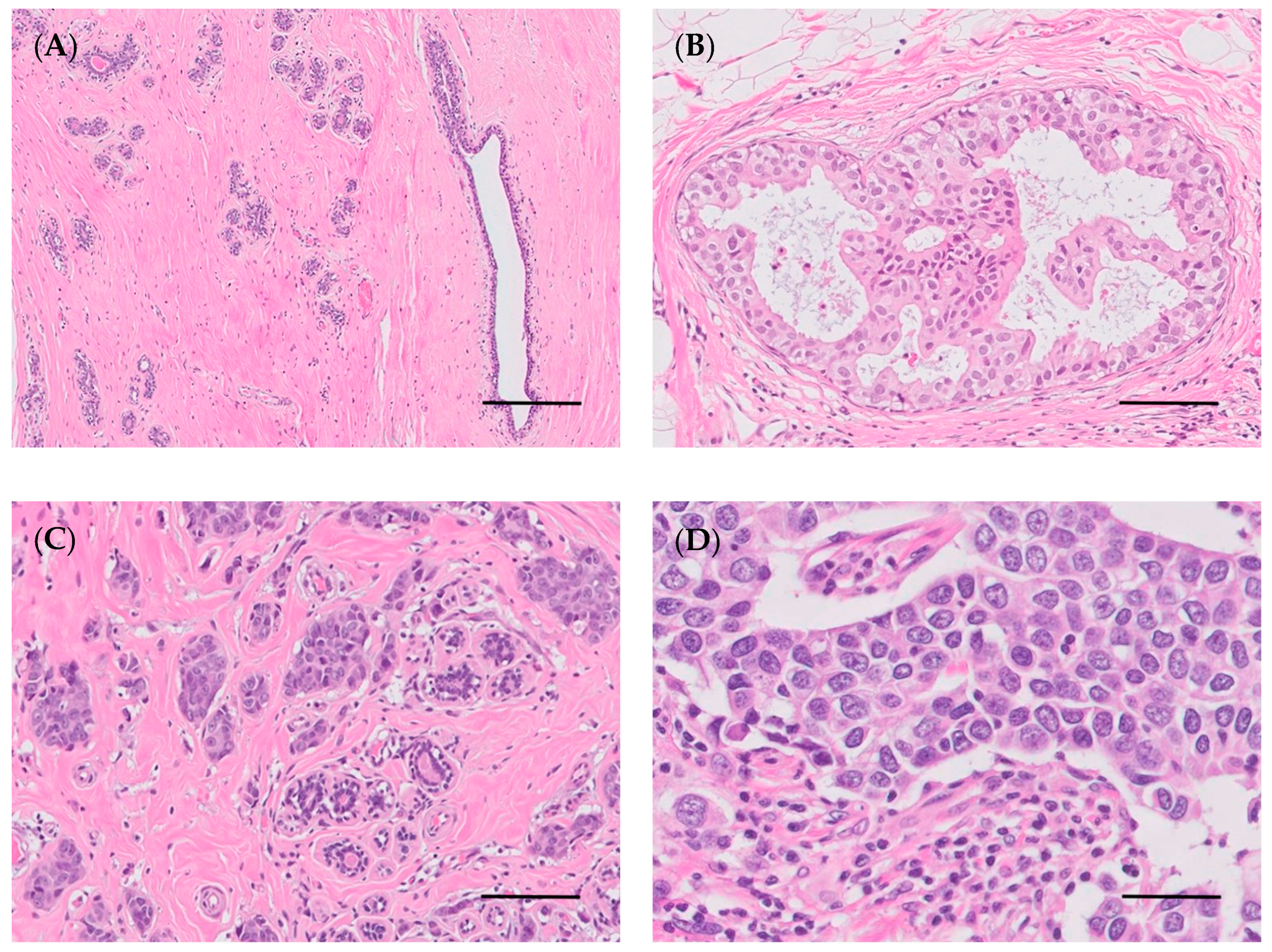
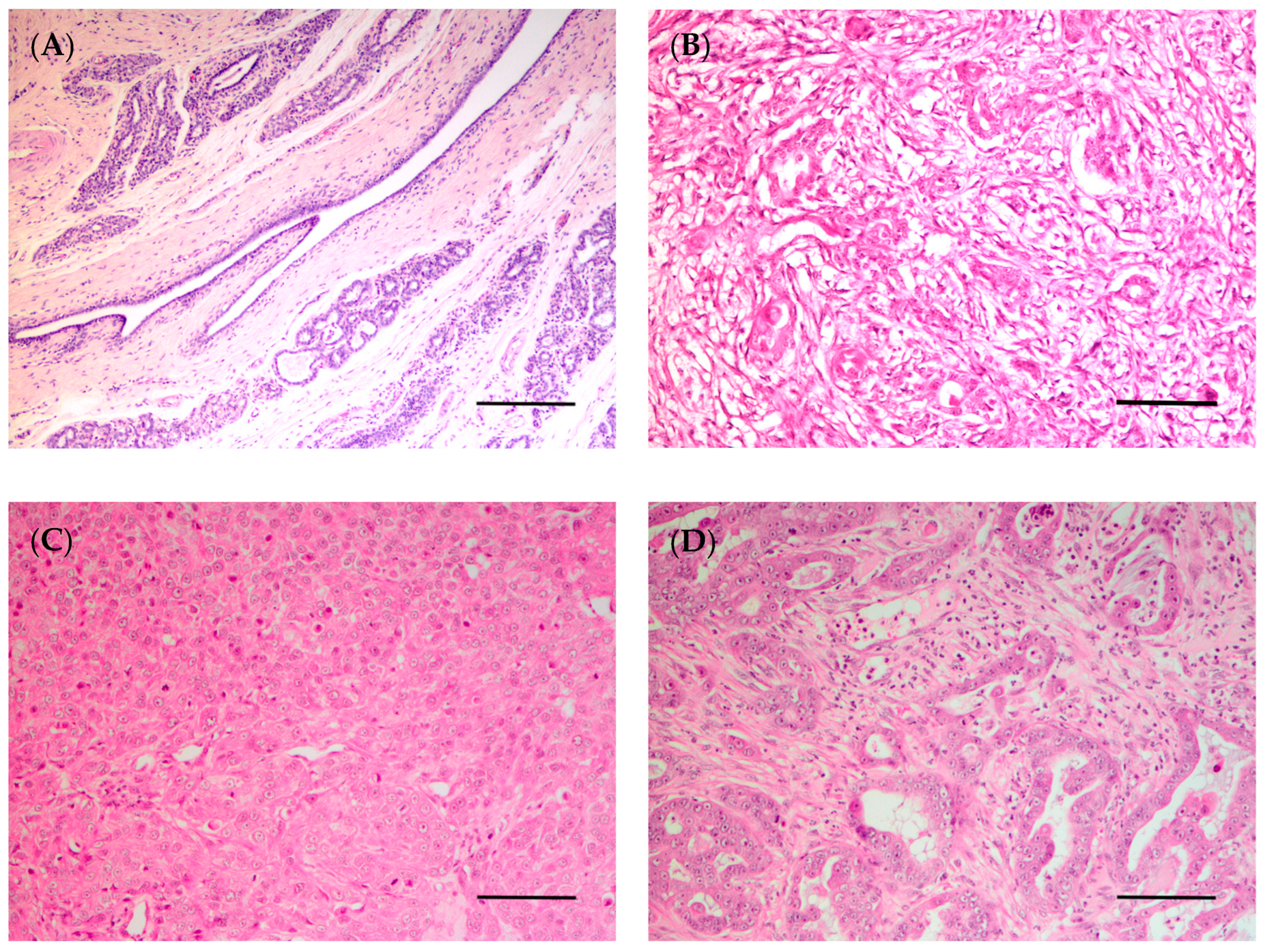
4.3. Histological and Molecular Classification of Breast/Mammary Tumors
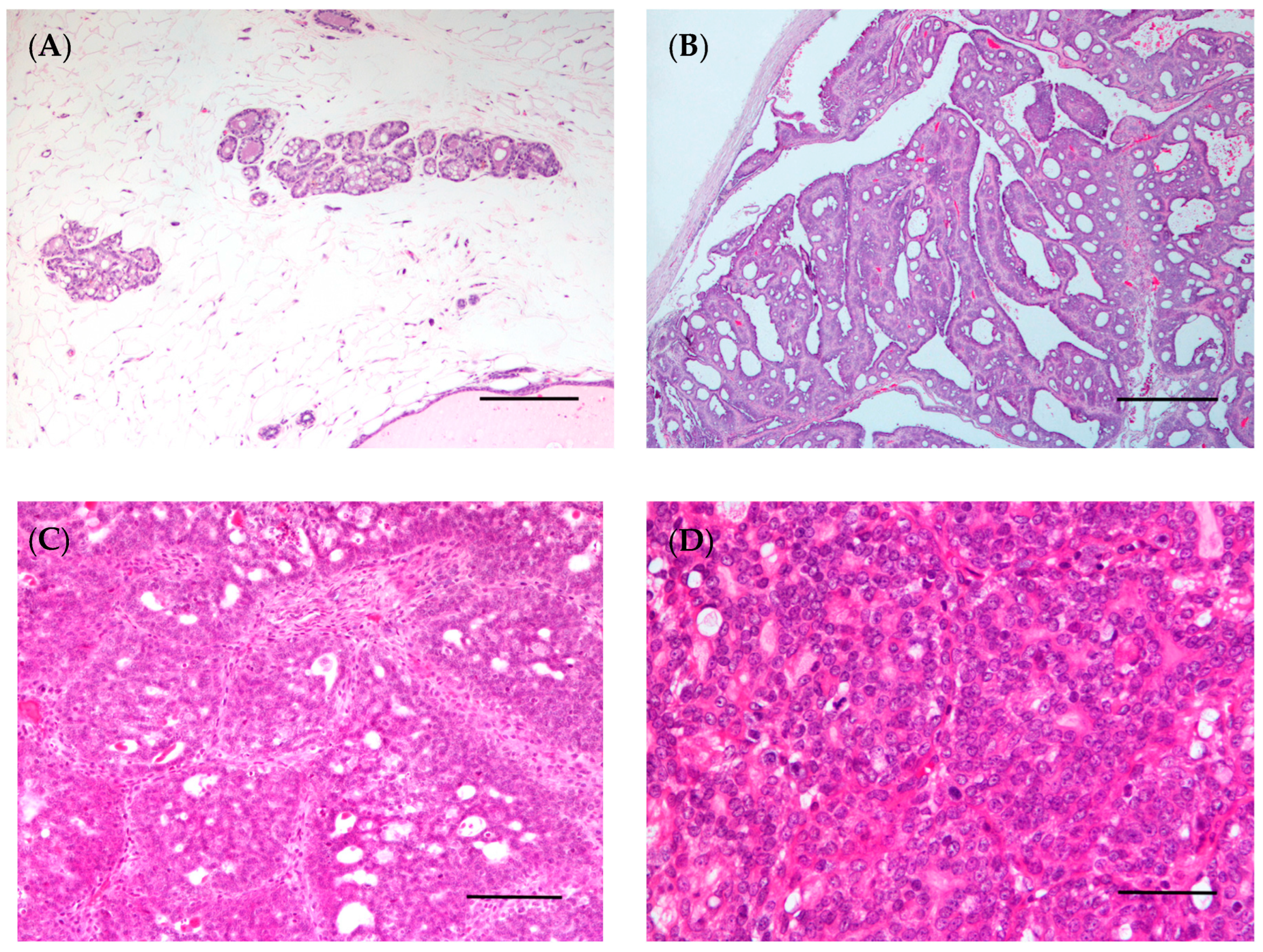
5. Advantages and Disadvantages of Spontaneous Tumors and Induced Animal Models for Studying Breast Cancer in Women
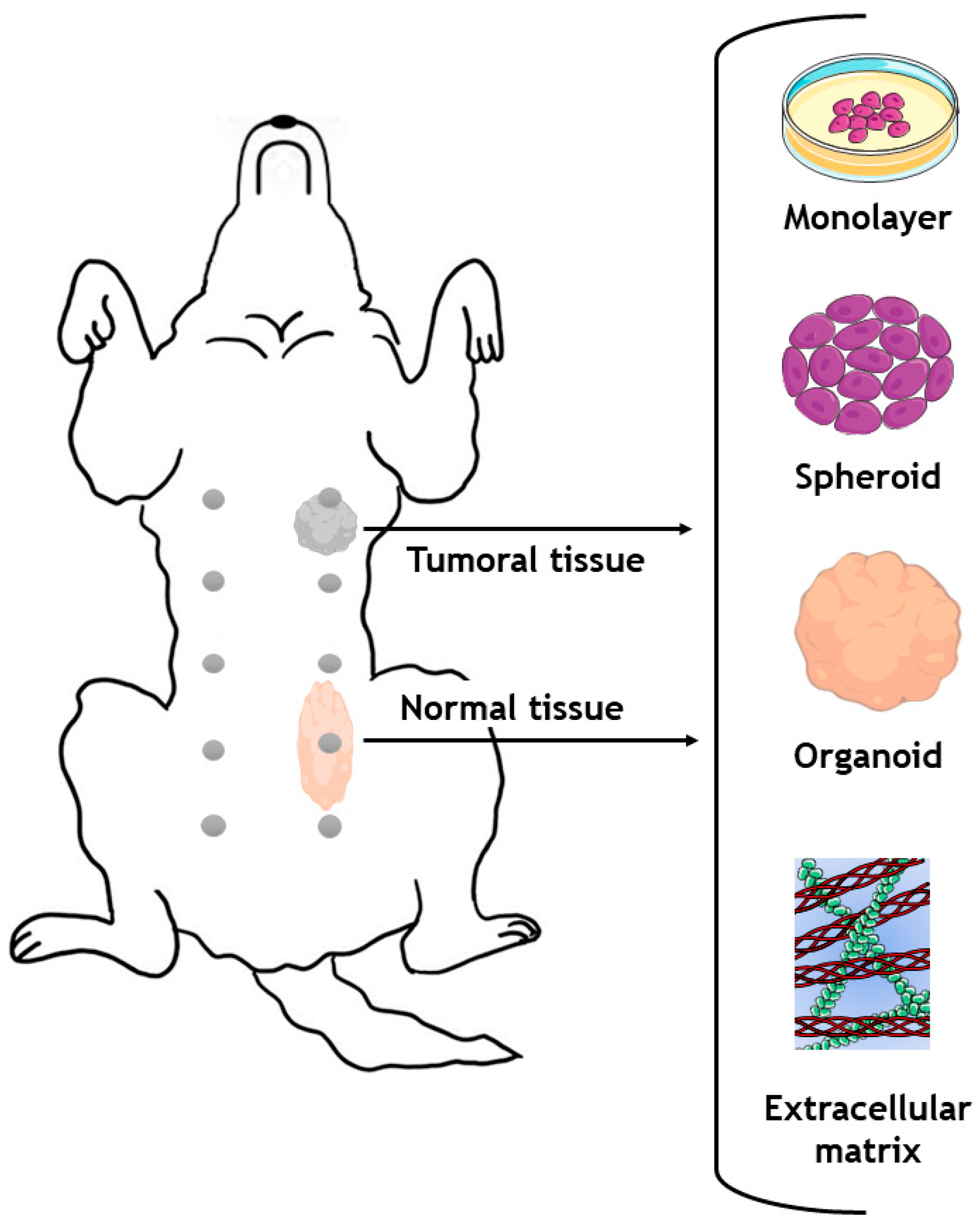
6. Spontaneous Tumors and Animal Models as a Source of Samples for Alternative 3D In Vitro Models
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Borecka, P.; Ratajczak-Wielgomas, K.; Ciaputa, R.; Kandefer-Gola, M.; Janus, I.; Piotrowska, A.; Kmiecik, A.; Podhorska-Okolów, M.; Dzięgiel, P.; Nowak, M. Expression of Periostin in Cancer-Associated Fibroblasts in Mammary Cancer in Female Dogs. In Vivo 2020, 34, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Tao, Z.; Shi, A.; Lu, C.; Song, T.; Zhang, Z.; Zhao, J. Breast Cancer: Epidemiology and Etiology. Cell Biochem. Biophys. 2015, 72, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Abadie, J.; Nguyen, F.; Loussouarn, D.; Peña, L.; Gama, A.; Rieder, N.; Belousov, A.; Bemelmans, I.; Jaillardon, L.; Ibisch, C.; et al. Canine Invasive Mammary Carcinomas as Models of Human Breast Cancer. Part 2: Immunophenotypes and Prognostic Significance. Breast Cancer Res. Treat. 2017, 167, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Li, W.; Chen, C.-S. Breast Cancer Animal Models and Applications. Zool. Res. 2020, 41, 477–494. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, A.; Faustino-Rocha, A.I.; Colaço, B.; Oliveira, P.A. Experimental Mammary Carcinogenesis—Rat Models. Life Sci. 2017, 173, 116–134. [Google Scholar] [CrossRef]
- Liu, N.; Johnson, K.J.; Ma, C.X. Male Breast Cancer: An Updated Surveillance, Epidemiology, and End Results Data Analysis. Clin. Breast Cancer 2018, 18, e997–e1002. [Google Scholar] [CrossRef] [PubMed]
- Ferzoco, R.M.; Ruddy, K.J. The Epidemiology of Male Breast Cancer. Curr. Oncol. Rep. 2016, 18, 1–6. [Google Scholar] [CrossRef]
- Saba, C.F.; Rogers, K.S.; Newman, S.J.; Mauldin, G.E.; Vail, D.M. Mammary Gland Tumors in Male Dogs. J. Vet. Intern. Med. 2007, 21, 1056–1059. [Google Scholar] [CrossRef]
- Salas, Y.; Márquez, A.; Diaz, D.; Romero, L. Epidemiological Study of Mammary Tumors in Female Dogs Diagnosed during the Period 2002-2012: A Growing Animal Health Problem. PLoS ONE 2015, 10, e0127381. [Google Scholar] [CrossRef]
- Pastor, N.; Caballé, N.C.; Santella, M.; Ezquerra, L.J.; Tarazona, R.; Duran, E. Epidemiological Study of Canine Mammary Tumors: Age, Breed, Size and Malignancy. Austral. J. Vet. Sci. 2018, 50, 143–147. [Google Scholar] [CrossRef]
- Okuda, M.; Takahashi, H.; Oyaizu, T.; Tsubura, A.; Morii, S.; Oishi, Y.; Fujii, T. Morphological Observations on Sexual Dimorphism in Rat Mammary Glands. J. Toxicol. Pathol. 1992, 5, 205–214. [Google Scholar] [CrossRef]
- Schiebinger, L. Why Mammals Are Called Mammals: Gender Politics in Eighteenth-Century Natural History. Am. Hist. Rev. 1993, 98, 382. [Google Scholar] [CrossRef] [PubMed]
- Schaller, O.; Constantinescu, G.M.; Constantinescu, G.C. (Eds.) Illustrated Veterinary Anatomical Nomenclature; Enke: Stuttgart, Germany, 1992; ISBN 978-3-432-99591-5. [Google Scholar]
- Silver, I.A. Symposium on Mammary Neoplasia in the Dog and Cat—I The Anatomy of the Mammary Gland of the Dog and Cat. J. Small Anim. Pract. 1966, 7, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Evans, H.E.; De Lahunta, A.; Miller, M.E. Miller’s Anatomy of the Dog; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 978-0-323-26623-9. [Google Scholar]
- Cardiff, R.D.; Jindal, S.; Treuting, P.M.; Going, J.J.; Gusterson, B.; Thompson, H.J. Mammary Gland. In Comparative Anatomy and Histology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 487–509. ISBN 978-0-12-802900-8. [Google Scholar]
- Hvid, H.; Thorup, I.; Oleksiewicz, M.B.; Sjögren, I.; Jensen, H.E. An Alternative Method for Preparation of Tissue Sections from the Rat Mammary Gland. Exp. Toxicol. Pathol. 2011, 63, 317–324. [Google Scholar] [CrossRef]
- Maeda, K.; Ohkura, S.; Tsukamura, H. Physiology of Reproduction. In The Laboratory Rat; Academic Press: San Diego, CA, USA, 2000; pp. 145–176. ISBN 9781281514509. [Google Scholar]
- Lucas, J.N.; Rudmann, D.G.; Credille, K.M.; Irizarry, A.R.; Peter, A.; Snyder, P.W. The Rat Mammary Gland: Morphologic Changes as an Indicator of Systemic Hormonal Perturbations Induced by Xenobiotics. Toxicol. Pathol. 2007, 35, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Eighmy, J.J.; Sharma, A.K.; Blackshear, P.E. Mammary Gland. In Boorman’s Pathology of the Rat; Elsevier: Amsterdam, The Netherlands, 2018; pp. 369–388. ISBN 978-0-12-391448-4. [Google Scholar]
- Russo, I.H.; Russo, J. Mammary Gland Neoplasia in Long-Term Rodent Studies. Environ. Health Perspect. 1996, 104, 938–967. [Google Scholar] [CrossRef] [PubMed]
- Propper, A.Y.; Howard, B.A.; Veltmaat, J.M. Prenatal Morphogenesis of Mammary Glands in Mouse and Rabbit. J. Mammary Gland. Biol. Neoplasia 2013, 18, 93–104. [Google Scholar] [CrossRef]
- Stolnicu, S. Histology of the Normal Breast, Normal Changes, and Abnormalities of Breast Development. In Practical Atlas of Breast Pathology; Stolnicu, S., Alvarado-Cabrero, I., Eds.; Springer: Cham, Switzerland, 2018; pp. 1–25. ISBN 978-3-319-93256-9. [Google Scholar]
- Macias, H.; Hinck, L. Mammary Gland Development: Mammary Gland Development. WIREs Dev. Biol. 2012, 1, 533–557. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, C.; Kettler, M.D.; Swirsky, M.E.; Miller, V.I.; Scott, C.; Krause, R.; Hadro, J.A. Male Breast Disease: Pictorial Review with Radiologic-Pathologic Correlation. RadioGraphics 2013, 33, 763–779. [Google Scholar] [CrossRef] [PubMed]
- Alex, A.; Bhandary, E.; McGuire, K.P. Anatomy and Physiology of the Breast during Pregnancy and Lactation. In Diseases of the Breast during Pregnancy and Lactation; Advances in Experimental Medicine and Biology; Alipour, S., Omranipour, R., Eds.; Springer: Cham, Switzerland, 2020; Volume 1252, pp. 3–7. ISBN 978-3-030-41595-2. [Google Scholar]
- Rehfeld, A.; Nylander, M.; Karnov, K. The Breast. In Compendium of Histology; Springer: Cham, Switzerland, 2017; pp. 593–601. ISBN 978-3-319-41871-1. [Google Scholar]
- Morris, E.A. The Normal Breast. In Breast MRI: Diagnosis and Intervention; Morris, E.A., Liberman, L., Eds.; Springer: New York, NY, USA, 2005; pp. 23–44. ISBN 978-0-387-27595-6. [Google Scholar]
- Brennan, P.A.; Standring, S.; Wiseman, S.; Gray, H. (Eds.) Gray’s Surgical Anatomy, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2019; ISBN 978-0-7020-7386-1. [Google Scholar]
- Watson, C.J.; Khaled, W.T. Mammary Development in the Embryo and Adult: A Journey of Morphogenesis and Commitment. Development 2008, 135, 995–1003. [Google Scholar] [CrossRef]
- Muschler, J.; Streuli, C.H. Cell-Matrix Interactions in Mammary Gland Development and Breast Cancer. Cold Spring Harb. Perspect. Biol. 2010, 2, a003202. [Google Scholar] [CrossRef] [PubMed]
- Hurley, W.L.; Loor, J.J.; Lacasse, P. Growth, Development and Involution. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2016; p. B978008100596521408X. ISBN 978-0-08-100596-5. [Google Scholar]
- Sopel, M. The Myoepithelial Cell: Its Role in Normal Mammary Glands and Breast Cancer. Folia Morphol. 2010, 69, 1–14. [Google Scholar]
- Haaksma, C.J.; Schwartz, R.J.; Tomasek, J.J. Myoepithelial Cell Contraction and Milk Ejection Are Impaired in Mammary Glands of Mice Lacking Smooth Muscle Alpha-Actin1. Biol. Reprod. 2011, 85, 13–21. [Google Scholar] [CrossRef]
- Raymond, K.; Cagnet, S.; Kreft, M.; Janssen, H.; Sonnenberg, A.; Glukhova, M.A. Control of Mammary Myoepithelial Cell Contractile Function by A3β1 Integrin Signalling: Integrin Signalling in the Myoepithelium. EMBO J. 2011, 30, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Gudjonsson, T.; Adriance, M.C.; Sternlicht, M.D.; Petersen, O.W.; Bissell, M.J. Myoepithelial Cells: Their Origin and Function in Breast Morphogenesis and Neoplasia. J. Mammary Gland Biol. Neoplasia 2005, 10, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Banik, U.; Parasuraman, S.; Adhikary, A.K.; Othman, N.H. Curcumin: The Spicy Modulator of Breast Carcinogenesis. J. Exp. Clin. Cancer Res. 2017, 36, 98. [Google Scholar] [CrossRef] [PubMed]
- Lowe, J.S.; Anderson, P.G.; Stevens, A. Stevens & Lowe’s Human Histology, 4th ed.; Elsevier: Philadelphia, PA, USA, 2015; ISBN 978-0-7234-3502-0. [Google Scholar]
- Hassiotou, F.; Geddes, D. Anatomy of the Human Mammary Gland: Current Status of Knowledge. Clin. Anat. 2013, 26, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Daniel, C.W.; Smith, G.H. The Mammary Gland: A Model for Development. J. Mammary Gland Biol. Neoplasia 1999, 4, 3–8. [Google Scholar] [CrossRef]
- Biswas, S.K.; Banerjee, S.; Baker, G.W.; Kuo, C.-Y.; Chowdhury, I. The Mammary Gland: Basic Structure and Molecular Signaling during Development. Int. J. Mol. Sci. 2022, 23, 3883. [Google Scholar] [CrossRef]
- Sleeckx, N.; de Rooster, H.; Veldhuis Kroeze, E.; Van Ginneken, C.; Van Brantegem, L. Canine Mammary Tumours, an Overview: Canine Mammary Tumours. Reprod. Domest. Anim. 2011, 46, 1112–1131. [Google Scholar] [CrossRef]
- Schlafer, D.H.; Foster, R.A. Female Genital System. In Jubb, Kennedy & Palmer’s Pathology of Domestic Animals: Volume 3; Elsevier: Amsterdam, The Netherlands, 2016; pp. 358–464.e1. ISBN 978-0-7020-5319-1. [Google Scholar]
- Sorenmo, K.U.; Rasotto, R.; Zappulli, V.; Goldschmidt, M.H. Development, Anatomy, Histology, Lymphatic Drainage, Clinical Features, and Cell Differentiation Markers of Canine Mammary Gland Neoplasms. Vet. Pathol. 2011, 48, 85–97. [Google Scholar] [CrossRef]
- Budras, K.-D. (Ed.) Anatomy of the Dog. In Vet, 5th ed.; Schlütersche: Hannover, Germany, 2007; ISBN 978-3-89993-018-4. [Google Scholar]
- Goldschmidt, M.H.; Peña, L.; Zappulli, V. Tumors of the Mammary Gland. In Tumors in Domestic Animals; Meuten, D.J., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2017; Volume 5. [Google Scholar]
- Zappulli, V.; Peña, L.; Rasotto, R.; Goldschmidt, M.H.; Gama, A.; Scruggs, J.L.; Kiupel, M. Volume 2: Mammary Tumors. In Surgical Pathology of Tumors of Domestic Animals; Davis-Thompson DVM Foundation: Gurnee, IL, USA, 2019; pp. 1–270. [Google Scholar]
- Lezama-García, K.; Mariti, C.; Mota-Rojas, D.; Martínez-Burnes, J.; Barrios-García, H.; Gazzano, A. Maternal Behaviour in Domestic Dogs. Int. J. Vet. Sci. Med. 2019, 7, 20–30. [Google Scholar] [CrossRef]
- van Zwieten, M.J. Normal Anatomy and Pathology of the Rat Mammary Gland. In The Rat as Animal Model in Breast Cancer Research; Springer Netherlands: Dordrecht, The Netherlands, 1984; pp. 53–134. ISBN 978-94-010-9000-1. [Google Scholar]
- Masso-Welch, P.A. A Developmental Atlas of Rat Mammary Gland Histology. J. Mammary Gland Biol. Neoplasia 2000, 5, 165–185. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, R.A.; Lawrence, R.M. Anatomy of the Breast. In Breastfeeding; Elsevier: Amsterdam, The Netherlands, 2011; pp. 40–61. ISBN 978-1-4377-0788-5. [Google Scholar]
- Dabbs, D.J. Breast Pathology, 2nd ed.; Elsevier: Philadelphia, PA, USA, 2017; ISBN 978-0-323-46236-5. [Google Scholar]
- Hammond, D.C. Applied Anatomy. In Atlas of Aesthetic Breast Surgery; Elsevier: Amsterdam, The Netherlands, 2009; pp. 1–9. ISBN 978-1-4160-3184-0. [Google Scholar]
- Patton, K.T.; Thibodeau, G.A. Anatomy & Physiology, 8th ed.; Mosby: St. Louis, MO, USA, 2013; ISBN 978-0-323-08357-7. [Google Scholar]
- Mugea, T.T.; Shiffman, M.A. (Eds.) Aesthetic Surgery of the Breast; Springer: Berlin, Germany, 2015; ISBN 978-3-662-43406-2. [Google Scholar]
- Collivignarelli, F.; Tamburro, R.; Aste, G.; Falerno, I.; Del Signore, F.; Simeoni, F.; Patsikas, M.; Gianfelici, J.; Terragni, R.; Attorri, V.; et al. Lymphatic Drainage Mapping with Indirect Lymphography for Canine Mammary Tumors. Animals 2021, 11, 1115. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Mohammed, S. Breast Cancer Metastasis and the Lymphatic System. Oncol. Lett. 2015, 10, 1233–1239. [Google Scholar] [CrossRef]
- Yu, J.; Mu, Q.; Fung, M.; Xu, X.; Zhu, L.; Ho, R.J.Y. Challenges and Opportunities in Metastatic Breast Cancer Treatments: Nano-Drug Combinations Delivered Preferentially to Metastatic Cells May Enhance Therapeutic Response. Pharmacol. Ther. 2022, 236, 108108. [Google Scholar] [CrossRef] [PubMed]
- Mumprecht, V.; Detmar, M. Lymphangiogenesis and Cancer Metastasis. J. Cell. Mol. Med. 2009, 13, 1405–1416. [Google Scholar] [CrossRef]
- Patsikas, M.N.; Dessiris, A. The Lymph Drainage of the Mammary Glands in the Bitch: A Lymphographic Study. Part I: The 1st, 2nd, 4th and 5th Mammary Glands. Anatom. Histol. Embryol. 1996, 25, 131–138. [Google Scholar] [CrossRef]
- Pereira, C.T.; Rahal, S.C.; Carvalho Balieiro, J.C.; Ribeiro, A.A.C.M. Lymphatic Drainage on Healthy and Neoplasic Mammary Glands in Female Dogs: Can It Really Be Altered? Anatom. Histol. Embryol. 2003, 32, 282–290. [Google Scholar] [CrossRef]
- Sheng, F.; Inoue, Y.; Kiryu, S.; Watanabe, M.; Ohtomo, K. Lymph Drainage from the Mammary Glands in Mice. Acad. Radiol. 2011, 18, 512–517. [Google Scholar] [CrossRef]
- Pinho, M.S.; Gulbenkian, S. Innervation of the Canine Mammary Gland: An Immunohistochemical Study. Histol. Histopathol. 2007, 22, 1175–1184. [Google Scholar] [CrossRef] [PubMed]
- Blacklock, A.D.; Smith, P.G. Estrogen Increases Calcitonin Gene-Related Peptide-Immunoreactive Sensory Innervation of Rat Mammary Gland. J. Neurobiol. 2004, 59, 192–204. [Google Scholar] [CrossRef]
- Van de Putte, P.; Blockmans, D.; De Rop, C.; Versyck, B. Pectoral Nerve Block Type II as the Sole Anesthetic for Removal of a Large Axillary Tumor: A Case Report. AA Pract. 2020, 14, e01201. [Google Scholar] [CrossRef]
- Jaspars, J.J.P.; Posma, A.N.; van Immerseel, A.A.H.; Groot, A.C.G. The Cutaneous Innervation of the Female Breast and Nipple-Areola Complex: Implications for Surgery. Br. J. Plast. Surg. 1997, 50, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Narayan, P.; Flynn, J.; Zhang, Z.; Gillespie, E.F.; Mueller, B.; Xu, A.J.; Cuaron, J.; McCormick, B.; Khan, A.J.; Cahlon, O.; et al. Perineural Invasion as a Risk Factor for Locoregional Recurrence of Invasive Breast Cancer. Sci. Rep. 2021, 11, 12781. [Google Scholar] [CrossRef]
- Huang, D.; Su, S.; Cui, X.; Shen, X.; Zeng, Y.; Wu, W.; Chen, J.; Chen, F.; He, C.; Liu, J.; et al. Nerve Fibers in Breast Cancer Tissues Indicate Aggressive Tumor Progression. Medicine 2014, 93, e172. [Google Scholar] [CrossRef] [PubMed]
- Gysler, S.M.; Drapkin, R. Tumor Innervation: Peripheral Nerves Take Control of the Tumor Microenvironment. J. Clin. Investig. 2021, 131, e147276. [Google Scholar] [CrossRef]
- Liebig, C.; Ayala, G.; Wilks, J.A.; Berger, D.H.; Albo, D. Perineural Invasion in Cancer: A Review of the Literature. Cancer 2009, 115, 3379–3391. [Google Scholar] [CrossRef]
- Kamiya, A.; Hayama, Y.; Kato, S.; Shimomura, A.; Shimomura, T.; Irie, K.; Kaneko, R.; Yanagawa, Y.; Kobayashi, K.; Ochiya, T. Genetic Manipulation of Autonomic Nerve Fiber Innervation and Activity and Its Effect on Breast Cancer Progression. Nat. Neurosci. 2019, 22, 1289–1305. [Google Scholar] [CrossRef] [PubMed]
- Vilhena, H.; Figueira, A.C.; Schmitt, F.; Canadas, A.; Chaves, R.; Gama, A.; Dias-Pereira, P. Canine and Feline Spontaneous Mammary Tumours as Models of Human Breast Cancer. In Pets as Sentinels, Forecasters and Promoters of Human Health; Pastorinho, M.R., Sousa, A.C.A., Eds.; Springer: Cham, Switzerland, 2020; pp. 173–207. ISBN 978-3-030-30733-2. [Google Scholar]
- Howard, J.; Wyse, C.; Argyle, D.; Quinn, C.; Kelly, P.; McCann, A. Exosomes as Biomarkers of Human and Feline Mammary Tumours; A Comparative Medicine Approach to Unravelling the Aggressiveness of TNBC. Biochim. Et Biophys. Acta BBA Rev. Cancer 2020, 1874, 188431. [Google Scholar] [CrossRef]
- Russo, J.; Russo, I.H. Experimentally Induced Mammary Tumors in Rats. Breast Cancer Res. Treat. 1996, 39, 7–20. [Google Scholar] [CrossRef]
- Sammarco, A.; Gomiero, C.; Sacchetto, R.; Beffagna, G.; Michieletto, S.; Orvieto, E.; Cavicchioli, L.; Gelain, M.E.; Ferro, S.; Patruno, M.; et al. Wnt/β-Catenin and Hippo Pathway Deregulation in Mammary Tumors of Humans, Dogs, and Cats. Vet. Pathol. 2020, 57, 774–790. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Siegel, R.; Ward, E.; Murray, T.; Xu, J.; Thun, M.J. Cancer Statistics, 2015. CA A Cancer J. Clin. 2015, 57, 43–66. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Hallberg, Ö.; Johansson, O. Sleep on the Right Side—Get Cancer on the Left? Pathophysiology 2010, 17, 157–160. [Google Scholar] [CrossRef]
- Hess, K.R.; Varadhachary, G.R.; Taylor, S.H.; Wei, W.; Raber, M.N.; Lenzi, R.; Abbruzzese, J.L. Metastatic Patterns in Adenocarcinoma. Cancer 2006, 106, 1624–1633. [Google Scholar] [CrossRef]
- Kvistad, K.A.; Rydland, J.; Smethurst, H.-B.; Lundgren, S.; Fjøsne, H.E.; Haraldseth, O. Axillary Lymph Node Metastases in Breast Cancer: Preoperative Detection with Dynamic Contrast-Enhanced MRI. Eur. Radiol. 2000, 10, 1464–1471. [Google Scholar] [CrossRef] [PubMed]
- Sorenmo, K. Canine Mammary Gland Tumors. Vet. Clin. North Am. Small Anim. Pract. 2003, 33, 573–596. [Google Scholar] [CrossRef]
- Bulman-Fleming, J. Mammary Cancer. In Clinical Small Animal Internal Medicine; Bruyette, D.S., Bexfield, N., Chretin, J.D., Kidd, L., Kube, S., Langston, C., Owen, T.J., Oyama, M.A., Peterson, N., Reiter, L.V., et al., Eds.; Wiley: Hoboken, NJ, USA, 2020; pp. 1321–1325. ISBN 978-1-118-49706-7. [Google Scholar]
- Moe, L. Population-Based Incidence of Mammary Tumours in Some Dog Breeds. J. Reprod. Fertil. Suppl. 2001, 57, 439–443. [Google Scholar] [PubMed]
- Dobson, J.M.; Samuel, S.; Milstein, H.; Rogers, K.; Wood, J.L.N. Canine Neoplasia in the UK: Estimates of Incidence Rates from a Population of Insured Dogs. J. Small Anim. Pract. 2002, 43, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Egenvall, A.; Bonnett, B.N.; Öhagen, P.; Olson, P.; Hedhammar, Å.; Euler, H. von Incidence of and Survival after Mammary Tumors in a Population of over 80,000 Insured Female Dogs in Sweden from 1995 to 2002. Prev. Vet. Med. 2005, 69, 109–127. [Google Scholar] [CrossRef]
- Canadas, A.; França, M.; Pereira, C.; Vilaça, R.; Vilhena, H.; Tinoco, F.; Silva, M.J.; Ribeiro, J.; Medeiros, R.; Oliveira, P.; et al. Canine Mammary Tumors: Comparison of Classification and Grading Methods in a Survival Study. Vet. Pathol. 2019, 56, 208–219. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Worley, D.R.; Zapulli, V. Tumors of the Mammary Gland. In Withrow and Macewen’s Small Animal Clinical Oncology; Vail, D.M., Thamm, D.H., Liptak, J.M., Eds.; Elsevier: Maryland Heights, MO, USA, 2019; pp. 604–625. ISBN 978-0-323-59496-7. [Google Scholar]
- Santos, T.R.; Castro, J.R.; Andrade, J.C.; Silva, A.C.R.; Silva, G.M.F.; Ferreira, F.A.; Headley, S.A.; Saut, J.P.E. Risk Factors Associated with Mammary Tumors in Female Dogs. Pesq. Vet. Bras. 2020, 40, 466–473. [Google Scholar] [CrossRef]
- Nguyen, F.; Peña, L.; Ibisch, C.; Loussouarn, D.; Gama, A.; Rieder, N.; Belousov, A.; Campone, M.; Abadie, J. Canine Invasive Mammary Carcinomas as Models of Human Breast Cancer. Part 1: Natural History and Prognostic Factors. Breast Cancer Res. Treat. 2018, 167, 635–648. [Google Scholar] [CrossRef]
- Sorenmo, K.U.; Kristiansen, V.M.; Cofone, M.A.; Shofer, F.S.; Breen, A.-M.; Langeland, M.; Mongil, C.M.; Grondahl, A.M.; Teige, J.; Goldschmidt, M.H. Canine Mammary Gland Tumours; a Histological Continuum from Benign to Malignant; Clinical and Histopathological Evidence. Vet. Comp. Oncol. 2009, 7, 162–172. [Google Scholar] [CrossRef]
- Klopfleisch, R.; Lenze, D.; Hummel, M.; Gruber, A.D. Metastatic Canine Mammary Carcinomas Can Be Identified by a Gene Expression Profile That Partly Overlaps with Human Breast Cancer Profiles. BMC Cancer 2010, 10, 618. [Google Scholar] [CrossRef] [PubMed]
- Oglesbee, B.L.; Oglesbee, B.L. (Eds.) Blackwell’s Five-Minute Veterinary Consult: Small Mammal, 2nd ed.; Five Minute Veterinary Consult; Wiley-Blackwell: Chichester, UK, 2011; ISBN 978-0-8138-2018-7. [Google Scholar]
- Dinse, G.E.; Peddada, S.D.; Harris, S.F.; Elmore, S.A. Comparison of NTP Historical Control Tumor Incidence Rates in Female Harlan Sprague Dawley and Fischer 344/N Rats. Toxicol. Pathol. 2010, 38, 765–775. [Google Scholar] [CrossRef]
- Planas-Silva, M.D.; Rutherford, T.M.; Stone, M.C. Prevention of Age-Related Spontaneous Mammary Tumors in Outbred Rats by Late Ovariectomy. Cancer Detect. Prev. 2008, 32, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Sharp, P.; Villano, J.S. (Eds.) The Laboratory Rat; CRC Press: Boca Raton, FL, USA, 2012; ISBN 978-0-429-15240-5. [Google Scholar]
- Russo, J.; Tay, L.K.; Russo, I.H. Differentiation of the Mammary Gland and Susceptibility to Carcinogenesis. Breast Cancer Res. Treat. 1982, 2, 5–73. [Google Scholar] [CrossRef] [PubMed]
- Russo, J. Significance of Rat Mammary Tumors for Human Risk Assessment. Toxicol. Pathol. 2015, 43, 145–170. [Google Scholar] [CrossRef]
- Faustino-Rocha, A.I.; Gama, A.; Oliveira, P.A.; Alvarado, A.; Neuparth, M.J.; Ferreira, R.; Ginja, M. Effects of Lifelong Exercise Training on Mammary Tumorigenesis Induced by MNU in Female Sprague–Dawley Rats. Clin. Exp. Med. 2017, 17, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Faustino-Rocha, A.I.; Ferreira, R.; Oliveira, P.A.; Gama, A.; Ginja, M. N-Methyl-N-Nitrosourea as a Mammary Carcinogenic Agent. Tumor Biol. 2015, 36, 9095–9117. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, A.; Lopes, A.C.; Faustino-Rocha, A.I.; Cabrita, A.M.S.; Ferreira, R.; Oliveira, P.A.; Colaço, B. Prognostic Factors in MNU and DMBA-Induced Mammary Tumors in Female Rats. Pathol. Res. Pract. 2017, 213, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Perše, M.; Cerar, A.; Injac, R.; Štrukelj, B. N-Methylnitrosourea Induced Breast Cancer in Rat, the Histopathology of the Resulting Tumours and Its Drawbacks as a Model. Pathol. Oncol. Res. 2009, 15, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Faustino-Rocha, A.; Oliveira, P.A.; Pinho-Oliveira, J.; Teixeira-Guedes, C.; Soares-Maia, R.; da Costa, R.G.; Colaço, B.; Pires, M.J.; Colaço, J.; Ferreira, R.; et al. Estimation of Rat Mammary Tumor Volume Using Caliper and Ultrasonography Measurements. Lab. Anim. 2013, 42, 217–224. [Google Scholar] [CrossRef]
- Flister, M.J.; Joshi, A.; Bergom, C.; Rui, H. Mapping Mammary Tumor Traits in the Rat. Methods Mol. Biol. 2019, 2018, 249–267. [Google Scholar] [CrossRef]
- Hiler, S.M.; Mina, A.; Mina, L.A. Breast Breast Cancer Risk Factors. In Breast Cancer Prevention and Treatment; Mina, L.A., Storniolo, A.M., Kipfer, H.D., Hunter, C., Ludwig, K.K., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 5–12. ISBN 9783319194370. [Google Scholar]
- San Miguel, Y.; Gomez, S.L.; Murphy, J.D.; Schwab, R.B.; McDaniels-Davidson, C.; Canchola, A.J.; Molinolo, A.A.; Nodora, J.N.; Martinez, M.E. Age-Related Differences in Breast Cancer Mortality According to Race/Ethnicity, Insurance, and Socioeconomic Status. BMC Cancer 2020, 20, 228. [Google Scholar] [CrossRef]
- Dai, Q.; Liu, B.; Du, Y. Meta-Analysis of the Risk Factors of Breast Cancer Concerning Reproductive Factors and Oral Contraceptive Use. Front. Med. 2009, 3, 452–458. [Google Scholar] [CrossRef]
- Kim, Y.; Yoo, K.-Y.; Goodman, M.T. Differences in Incidence, Mortality and Survival of Breast Cancer by Regions and Countries in Asia and Contributing Factors. Asian Pac. J. Cancer Prev. 2015, 16, 2857–2870. [Google Scholar] [CrossRef]
- Thakur, P.; Seam, R.; Gupta, M.; Gupta, M.; Sharma, M.; Fotedar, V. Breast Cancer Risk Factor Evaluation in a Western Himalayan State: A Case–Control Study and Comparison with the Western World. South Asian J. Cancer 2017, 6, 106. [Google Scholar] [CrossRef]
- Melbye, M.; Wohlfahrt, J.; Andersen, A.-M.N.; Westergaard, T.; Andersen, P.K. Preterm Delivery and Risk of Breast Cancer. Br. J. Cancer 1999, 80, 609–613. [Google Scholar] [CrossRef] [PubMed]
- Innes, K.E.; Byers, T.E. First Pregnancy Characteristics and Subsequent Breast Cancer Risk among Young Women. Int. J. Cancer 2004, 112, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Momenimovahed, Z.; Salehiniya, H. Epidemiological Characteristics of and Risk Factors for Breast Cancer in the World. Breast Cancer Targets Ther. 2019, 11, 151–164. [Google Scholar] [CrossRef] [PubMed]
- Gleicher, N. Why Are Reproductive Cancers More Common in Nulliparous Women? Reprod. BioMedicine Online 2013, 26, 416–419. [Google Scholar] [CrossRef] [PubMed]
- Kapil, U.; Sareen, N.; Singh, P.; Bhadoria, A. Reproductive Factors and Breast Cancer: A Case-Control Study in Tertiary Care Hospital of North India. Indian J. Cancer 2013, 50, 316. [Google Scholar] [CrossRef]
- Mørch, L.S.; Skovlund, C.W.; Hannaford, P.C.; Iversen, L.; Fielding, S.; Lidegaard, Ø. Contemporary Hormonal Contraception and the Risk of Breast Cancer. N. Engl. J. Med. 2017, 377, 2228–2239. [Google Scholar] [CrossRef]
- Zolfaroli, I.; Tarín, J.J.; Cano, A. Hormonal Contraceptives and Breast Cancer: Clinical Data. Eur. J. Obstet. Gynecol. Reprod. Biol. 2018, 230, 212–216. [Google Scholar] [CrossRef]
- Manson, J.E.; Chlebowski, R.T.; Stefanick, M.L.; Aragaki, A.K.; Rossouw, J.E.; Prentice, R.L.; Anderson, G.; Howard, B.V.; Thomson, C.A.; LaCroix, A.Z.; et al. Menopausal Hormone Therapy and Health Outcomes During the Intervention and Extended Poststopping Phases of the Women’s Health Initiative Randomized Trials. JAMA 2013, 310, 1353. [Google Scholar] [CrossRef]
- The 2017 Hormone Therapy Position Statement of The North American Menopause Society. Menopause 2017, 24, 728–753. [CrossRef]
- Writing Group for the Women’s Health Initiative Investigators Risks and Benefits of Estrogen Plus Progestin in Healthy Postmenopausal Women: Principal Results From the Women’s Health Initiative Randomized Controlled Trial. JAMA J. Am. Med. Assoc. 2002, 288, 321–333. [CrossRef]
- Press, D.J.; Sullivan-Halley, J.; Ursin, G.; Deapen, D.; McDonald, J.A.; Strom, B.L.; Norman, S.A.; Simon, M.S.; Marchbanks, P.A.; Folger, S.G.; et al. Breast Cancer Risk and Ovariectomy, Hysterectomy, and Tubal Sterilization in the Women’s Contraceptive and Reproductive Experiences Study. Am. J. Epidemiol. 2011, 173, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Mohanty, S.S.; Mohanty, P.K. Obesity as Potential Breast Cancer Risk Factor for Postmenopausal Women. Genes Dis. 2021, 8, 117–123. [Google Scholar] [CrossRef] [PubMed]
- Farvid, M.S.; Sidahmed, E.; Spence, N.D.; Mante Angua, K.; Rosner, B.A.; Barnett, J.B. Consumption of Red Meat and Processed Meat and Cancer Incidence: A Systematic Review and Meta-Analysis of Prospective Studies. Eur. J. Epidemiol. 2021, 36, 937–951. [Google Scholar] [CrossRef]
- Möller, S.; Mucci, L.A.; Harris, J.R.; Scheike, T.; Holst, K.; Halekoh, U.; Adami, H.-O.; Czene, K.; Christensen, K.; Holm, N.V.; et al. The Heritability of Breast Cancer among Women in the Nordic Twin Study of Cancer. Cancer Epidemiol. Biomark. Prev. 2016, 25, 145–150. [Google Scholar] [CrossRef]
- Pouptsis, A.; Swafe, L.; Patwardhan, M.; Stavraka, C. Surgical and Systemic Treatment of Hereditary Breast Cancer: A Mini-Review With a Focus on BRCA1 and BRCA2 Mutations. Front. Oncol. 2020, 10, 553080. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, A.; Pharoah, P.D.P.; Narod, S.; Risch, H.A.; Eyfjord, J.E.; Hopper, J.L.; Loman, N.; Olsson, H.; Johannsson, O.; Borg, Å.; et al. Average Risks of Breast and Ovarian Cancer Associated with BRCA1 or BRCA2 Mutations Detected in Case Series Unselected for Family History: A Combined Analysis of 22 Studies. Am. J. Hum. Genet. 2003, 72, 1117–1130. [Google Scholar] [CrossRef]
- Breast Cancer Association Consortium; Dorling, L.; Carvalho, S.; Allen, J.; González-Neira, A.; Luccarini, C.; Wahlström, C.; Pooley, K.A.; Parsons, M.T.; Fortuno, C.; et al. Breast Cancer Risk Genes—Association Analysis in More than 113,000 Women. N. Engl. J. Med. 2021, 384, 428–439. [Google Scholar] [CrossRef] [PubMed]
- Park, S.-Y.; Kolonel, L.N.; Lim, U.; White, K.K.; Henderson, B.E.; Wilkens, L.R. Alcohol Consumption and Breast Cancer Risk among Women from Five Ethnic Groups with Light to Moderate Intakes: The Multiethnic Cohort Study: Alcohol and Breast Cancer. Int. J. Cancer 2014, 134, 1504–1510. [Google Scholar] [CrossRef]
- Vieira, R.; Tobar, J.S.S.; Dardes, R.; Santos Thuler, L.C. Alcohol Consumption as a Risk Factor for Breast Cancer Development: A Case-Control Study in Brazil. Asian Pac. J. Cancer Prev. 2018, 19, 703–707. [Google Scholar] [CrossRef]
- Miller, E.R.; Wilson, C.; Chapman, J.; Flight, I.; Nguyen, A.-M.; Fletcher, C.; Ramsey, I. Connecting the Dots between Breast Cancer, Obesity and Alcohol Consumption in Middle-Aged Women: Ecological and Case Control Studies. BMC Public Health 2018, 18, 460. [Google Scholar] [CrossRef]
- Xue, F.; Willett, W.C.; Rosner, B.A.; Hankinson, S.E.; Michels, K.B. Cigarette Smoking and the Incidence of Breast Cancer. Arch. Intern. Med. 2011, 171, 125–133. [Google Scholar] [CrossRef]
- Guo, W.; Fensom, G.K.; Reeves, G.K.; Key, T.J. Physical Activity and Breast Cancer Risk: Results from the UK Biobank Prospective Cohort. Br. J. Cancer 2020, 122, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Taylor, E.F.; Burley, V.J.; Greenwood, D.C.; Cade, J.E. Meat Consumption and Risk of Breast Cancer in the UK Women’s Cohort Study. Br. J. Cancer 2007, 96, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Rosner, B.; Eliassen, A.H.; Toriola, A.T.; Chen, W.Y.; Hankinson, S.E.; Willett, W.C.; Berkey, C.S.; Colditz, G.A. Weight and Weight Changes in Early Adulthood and Later Breast Cancer Risk: Weight Change and Breast Cancer Risk. Int. J. Cancer 2017, 140, 2003–2014. [Google Scholar] [CrossRef] [PubMed]
- Lope, V.; Martín, M.; Castelló, A.; Ruiz, A.; Casas, A.M.; Baena-Cañada, J.M.; Antolín, S.; Ramos-Vázquez, M.; García-Sáenz, J.Á.; Muñoz, M.; et al. Overeating, Caloric Restriction and Breast Cancer Risk by Pathologic Subtype: The EPIGEICAM Study. Sci. Rep. 2019, 9, 3904. [Google Scholar] [CrossRef]
- Abdelmegeed, S.; Mohammed, S. Canine Mammary Tumors as a Model for Human Disease (Review). Oncol. Lett. 2018, 15, 8195–8205. [Google Scholar] [CrossRef]
- Ariyarathna, H.; de Silva, N.; Aberdein, D.; Kodikara, D.; Jayasinghe, M.; Adikari, R.; Munday, J. Clinicopathological Diversity of Canine Mammary Gland Tumors in Sri Lanka: A One-Year Survey on Cases Presented to Two Veterinary Practices. Vet. Sci. 2018, 5, 46. [Google Scholar] [CrossRef]
- Beauvais, W.; Cardwell, J.M.; Brodbelt, D.C. The Effect of Neutering on the Risk of Mammary Tumours in Dogs—A Systematic Review. J. Small Anim. Pract. 2012, 53, 314–322. [Google Scholar] [CrossRef]
- MacPhail, C. Surgery of the Reproductive and Genital Systems. In Small Animal Surgery; Fossum, T.W., Ed.; Elsevier: Philadelphia, PA, USA, 2019; ISBN 978-0-323-44344-9. [Google Scholar]
- Rivera, P.; Melin, M.; Biagi, T.; Fall, T.; Häggström, J.; Lindblad-Toh, K.; von Euler, H. Mammary Tumor Development in Dogs Is Associated with BRCA1 and BRCA2. Cancer Res. 2009, 69, 8770–8774. [Google Scholar] [CrossRef]
- Lim, H.-Y.; Im, K.-S.; Kim, N.-H.; Kim, H.-W.; Shin, J.-I.; Yhee, J.-Y.; Sur, J.-H. Effects of Obesity and Obesity-Related Molecules on Canine Mammary Gland Tumors. Vet. Pathol. 2015, 52, 1045–1051. [Google Scholar] [CrossRef]
- Sonnenschein, E.G.; Glickman, L.T.; Goldschmidt, M.H.; McKee, L.J. Body Conformation, Diet, and Risk of Breast Cancer in Pet Dogs: A Case-Control Study. Am. J. Epidemiol. 1991, 133, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Alenza, D.P.; Rutteman, G.R.; Peña, L.; Beynen, A.C.; Cuesta, P. Relation between Habitual Diet and Canine Mammary Tumors in a Case-Control Study. J. Vet. Intern. Med. 1998, 12, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Salas, Y.J.; Aburto, E.; Alonso, R.; Márquez-Alvarado, A.A.; Corona-Monjaras, H.; Romero-Romero, L. Association of Histological Features with Potential Risk Factors and Survival in Canine Mammary Tumors. Vet. Méx. OA 2016, 3. [Google Scholar] [CrossRef]
- Itoh, T.; Uchida, K.; Ishikawa, K.; Kushima, K.; Kushima, E.; Tamada, H.; Moritake, T.; Nakao, H.; Shii, H. Clinicopathological Survey of 101 Canine Mammary Gland Tumors: Differences between Small-Breed Dogs and Others. J. Vet. Med. Sci. 2005, 67, 345–347. [Google Scholar] [CrossRef]
- Harbeck, N.; Penault-Llorca, F.; Cortes, J.; Gnant, M.; Houssami, N.; Poortmans, P.; Ruddy, K.; Tsang, J.; Cardoso, F. Breast Cancer. Nat. Rev. Dis. Prim. 2019, 5, 66. [Google Scholar] [CrossRef]
- Malhotra, G.K.; Zhao, X.; Band, H.; Band, V. Histological, Molecular and Functional Subtypes of Breast Cancers. Cancer Biol. Ther. 2010, 10, 955–960. [Google Scholar] [CrossRef]
- Allred, D.C. Ductal Carcinoma in Situ: Terminology, Classification, and Natural History. J. Natl. Cancer Inst. Monogr. 2010, 2010, 134–138. [Google Scholar] [CrossRef]
- Sharma, G.N.; Dave, R.; Sanadya, J.; Sharma, P.; Sharma, K.K. Various Types and Management of Breast Cancer: An Overview. J. Adv. Pharm. Technol. Res. 2010, 1, 109–126. [Google Scholar]
- Elston, C.W.; Ellis, I.O. Pathological Prognostic Factors in Breast Cancer. I. The Value of Histological Grade in Breast Cancer: Experience from a Large Study with Long-Term Follow-Up. Histopathology 1991, 19, 403–410. [Google Scholar] [CrossRef]
- Dalton, L.W.; Page, D.L.; Dupont, W.D. Histologic Grading of Breast Carcinoma. A Reproducibility Study. Cancer 1994, 73, 2765–2770. [Google Scholar] [CrossRef]
- Dalle, J.-R.; Leow, W.K.; Racoceanu, D.; Tutac, A.E.; Putti, T.C. Automatic Breast Cancer Grading of Histopathological Images. In Proceedings of the 2008 30th Annual International Conference of the IEEE Engineering in Medicine and Biology Society, IEEE, Vancouver, BC, USA, 17 June 2008; pp. 3052–3055. [Google Scholar]
- Boix-Montesinos, P.; Soriano-Teruel, P.M.; Armiñán, A.; Orzáez, M.; Vicent, M.J. The Past, Present, and Future of Breast Cancer Models for Nanomedicine Development. Adv. Drug Deliv. Rev. 2021, 173, 306–330. [Google Scholar] [CrossRef] [PubMed]
- Kalli, S.; Semine, A.; Cohen, S.; Naber, S.P.; Makim, S.S.; Bahl, M. American Joint Committee on Cancer’s Staging System for Breast Cancer, Eighth Edition: What the Radiologist Needs to Know. RadioGraphics 2018, 38, 1921–1933. [Google Scholar] [CrossRef] [PubMed]
- Rakha, E.A.; Reis-Filho, J.S.; Baehner, F.; Dabbs, D.J.; Decker, T.; Eusebi, V.; Fox, S.B.; Ichihara, S.; Jacquemier, J.; Lakhani, S.R.; et al. Breast Cancer Prognostic Classification in the Molecular Era: The Role of Histological Grade. Breast Cancer Res. 2010, 12, 207. [Google Scholar] [CrossRef]
- Rakha, E.A.; El-Sayed, M.E.; Lee, A.H.S.; Elston, C.W.; Grainge, M.J.; Hodi, Z.; Blamey, R.W.; Ellis, I.O. Prognostic Significance of Nottingham Histologic Grade in Invasive Breast Carcinoma. J. Clin. Oncol. 2008, 26, 3153–3158. [Google Scholar] [CrossRef]
- Payne, S.J.L.; Bowen, R.L.; Jones, J.L.; Wells, C.A. Predictive Markers in Breast Cancer—The Present. Histopathology 2007, 52, 82–90. [Google Scholar] [CrossRef]
- Tsang, J.Y.S.; Tse, G.M. Molecular Classification of Breast Cancer. Adv. Anat. Pathol. 2020, 27, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Rakha, E.A.; Reis-Filho, J.S.; Ellis, I.O. Basal-like Breast Cancer: A Critical Review. J. Clin. Oncol. 2008, 26, 2568–2581. [Google Scholar] [CrossRef]
- Badowska-Kozakiewicz, A.M.; Budzik, M.P. Immunohistochemical Characteristics of Basal-like Breast Cancer. Contemp. Oncol. 2016, 6, 436–443. [Google Scholar] [CrossRef]
- Al-thoubaity, F.K. Molecular Classification of Breast Cancer: A Retrospective Cohort Study. Ann. Med. Surg. 2020, 49, 44–48. [Google Scholar] [CrossRef]
- Waks, A.G.; Winer, E.P. Breast Cancer Treatment: A Review. JAMA 2019, 321, 288. [Google Scholar] [CrossRef]
- Feng, Y.; Spezia, M.; Huang, S.; Yuan, C.; Zeng, Z.; Zhang, L.; Ji, X.; Liu, W.; Huang, B.; Luo, W.; et al. Breast Cancer Development and Progression: Risk Factors, Cancer Stem Cells, Signaling Pathways, Genomics, and Molecular Pathogenesis. Genes Dis. 2018, 5, 77–106. [Google Scholar] [CrossRef]
- Hashmi, A.A.; Aijaz, S.; Khan, S.M.; Mahboob, R.; Irfan, M.; Zafar, N.I.; Nisar, M.; Siddiqui, M.; Edhi, M.M.; Faridi, N.; et al. Prognostic Parameters of Luminal A and Luminal B Intrinsic Breast Cancer Subtypes of Pakistani Patients. World J. Surg. Oncol. 2018, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Partridge, A.H.; Hughes, M.E.; Warner, E.T.; Ottesen, R.A.; Wong, Y.-N.; Edge, S.B.; Theriault, R.L.; Blayney, D.W.; Niland, J.C.; Winer, E.P.; et al. Subtype-Dependent Relationship Between Young Age at Diagnosis and Breast Cancer Survival. J. Clin. Oncol. 2016, 34, 3308–3314. [Google Scholar] [CrossRef]
- Lau, K.H.; Tan, A.M.; Shi, Y. New and Emerging Targeted Therapies for Advanced Breast Cancer. Int. J. Mol. Sci. 2022, 23, 2288. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, N.; Iqbal, N. Human Epidermal Growth Factor Receptor 2 (HER2) in Cancers: Overexpression and Therapeutic Implications. Mol. Biol. Int. 2014, 2014, 852748. [Google Scholar] [CrossRef] [PubMed]
- Vrbic, S.; Pejcic, I.; Filipovic, S.; Kocic, B.; Vrbic, M. Current and Future Anti-HER2 Therapy in Breast Cancer. J. Buon 2013, 18, 4–16. [Google Scholar]
- Yao, M.; Fu, P. Advances in Anti-HER2 Therapy in Metastatic Breast Cancer. Chin. Clin. Oncol. 2018, 7, 27. [Google Scholar] [CrossRef]
- Pascual, T.; Martin, M.; Fernández-Martínez, A.; Paré, L.; Alba, E.; Rodríguez-Lescure, Á.; Perrone, G.; Cortés, J.; Morales, S.; Lluch, A.; et al. A Pathology-Based Combined Model to Identify PAM50 Non-Luminal Intrinsic Disease in Hormone Receptor-Positive HER2-Negative Breast Cancer. Front. Oncol. 2019, 9, 303. [Google Scholar] [CrossRef]
- Yin, L.; Duan, J.-J.; Bian, X.-W.; Yu, S. Triple-Negative Breast Cancer Molecular Subtyping and Treatment Progress. Breast Cancer Res. 2020, 22, 61. [Google Scholar] [CrossRef]
- Peddi, P.F.; Ellis, M.J.; Ma, C. Molecular Basis of Triple Negative Breast Cancer and Implications for Therapy. Int. J. Breast Cancer 2012, 2012, 1–7. [Google Scholar] [CrossRef]
- Greenup, R.; Buchanan, A.; Lorizio, W.; Rhoads, K.; Chan, S.; Leedom, T.; King, R.; McLennan, J.; Crawford, B.; Kelly Marcom, P.; et al. Prevalence of BRCA Mutations Among Women with Triple-Negative Breast Cancer (TNBC) in a Genetic Counseling Cohort. Ann. Surg. Oncol. 2013, 20, 3254–3258. [Google Scholar] [CrossRef]
- Goldschmidt, M.; Peña, L.; Rasotto, R.; Zappulli, V. Classification and Grading of Canine Mammary Tumors. Vet. Pathol. 2011, 48, 117–131. [Google Scholar] [CrossRef]
- Burrai, G.P.; Gabrieli, A.; Moccia, V.; Zappulli, V.; Porcellato, I.; Brachelente, C.; Pirino, S.; Polinas, M.; Antuofermo, E. A Statistical Analysis of Risk Factors and Biological Behavior in Canine Mammary Tumors: A Multicenter Study. Animals 2020, 10, 1687. [Google Scholar] [CrossRef]
- Pastor, N.; Espadas, L.; Santella, M.; Ezquerra, L.J.; Tarazona, R.; Durán, M.E. Comparison between Histological Features and Strain Elastographic Characteristics in Canine Mammary Carcinomas. Vet. Sci. 2021, 9, 9. [Google Scholar] [CrossRef]
- Gonçalves, A.D.B.B.; de Pinho, T.C.R.; Toledo, F.A.O.; Balabram, D.; Ferreira, E.; Campos, M.R.A.; Cassali, G.D. Extracapsular Extension and Tumor Implants in Lymph Nodes with Canine Mammary Carcinoma Metastasis: Study of the Impact on Survival. Res. Vet. Sci. 2021, 139, 87–93. [Google Scholar] [CrossRef]
- Levi, M.; Muscatello, L.V.; Brunetti, B.; Benazzi, C.; Parenti, F.; Gobbo, F.; Avallone, G.; Bacci, B.; Zambon, E.; Valenti, P.; et al. High Intrinsic Expression of P-Glycoprotein and Breast Cancer Resistance Protein in Canine Mammary Carcinomas Regardless of Immunophenotype and Outcome. Animals 2021, 11, 658. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; Lai, Y.-C.; Rahman, M.M.; Husna, A.A.; Hasan, M.N.; Hatai, H.; Miyoshi, N.; Yamato, O.; Miura, N. NGS-Identified MiRNAs in Canine Mammary Gland Tumors Show Unexpected Expression Alterations in QPCR Analysis. In Vivo 2022, 36, 1628–1636. [Google Scholar] [CrossRef]
- Tanaka, Y.; Koyama, K.; Horiuchi, N.; Watanabe, K.; Kobayashi, Y. Relationship between Histological Grade and Histopathological Appearance in Canine Mammary Carcinomas. J. Comp. Pathol. 2020, 179, 59–64. [Google Scholar] [CrossRef] [PubMed]
- Pinello, K.; Baldassarre, V.; Steiger, K.; Paciello, O.; Pires, I.; Laufer-Amorim, R.; Oevermann, A.; Niza-Ribeiro, J.; Aresu, L.; Rous, B.; et al. Vet-ICD-O-Canine-1, a System for Coding Canine Neoplasms Based on the Human ICD-O-3.2. Cancers 2022, 14, 1529. [Google Scholar] [CrossRef] [PubMed]
- Peña, L.; Andrés, P.J.D.; Clemente, M.; Cuesta, P.; Pérez-Alenza, M.D. Prognostic Value of Histological Grading in Noninflammatory Canine Mammary Carcinomas in a Prospective Study With Two-Year Follow-Up: Relationship With Clinical and Histological Characteristics. Vet. Pathol. 2013, 50, 94–105. [Google Scholar] [CrossRef]
- Rutteman, G.R.; Withrow, S.J.; MacEwen, E. Tumors of the Mammary Gland. In Small Animal Clinical Oncology; WB Saunders: Philadelphia, PA, USA, 2001; pp. 455–477. [Google Scholar]
- Gama, A.; Alves, A.; Schmitt, F. Identification of Molecular Phenotypes in Canine Mammary Carcinomas with Clinical Implications: Application of the Human Classification. Virchows Arch. 2008, 453, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Amirkhani Namagerdi, A.; d’Angelo, D.; Ciani, F.; Iannuzzi, C.A.; Napolitano, F.; Avallone, L.; De Laurentiis, M.; Giordano, A. Triple-Negative Breast Cancer Comparison With Canine Mammary Tumors From Light Microscopy to Molecular Pathology. Front. Oncol. 2020, 10, 563779. [Google Scholar] [CrossRef] [PubMed]
- Pastor, N.; Ezquerra, L.J.; Santella, M.; Caballé, N.C.; Tarazona, R.; Durán, M.E. Prognostic Significance of Immunohistochemical Markers and Histological Classification in Malignant Canine Mammary Tumours. Vet. Comp. Oncol. 2020, 18, 753–762. [Google Scholar] [CrossRef]
- Peña, L.; Gama, A.; Goldschmidt, M.H.; Abadie, J.; Benazzi, C.; Castagnaro, M.; Díez, L.; Gärtner, F.; Hellmén, E.; Kiupel, M.; et al. Canine Mammary Tumors: A Review and Consensus of Standard Guidelines on Epithelial and Myoepithelial Phenotype Markers, HER2, and Hormone Receptor Assessment Using Immunohistochemistry. Vet. Pathol. 2014, 51, 127–145. [Google Scholar] [CrossRef]
- Im, K.S.; Kim, N.H.; Lim, H.Y.; Kim, H.W.; Shin, J.I.; Sur, J.H. Analysis of a New Histological and Molecular-Based Classification of Canine Mammary Neoplasia. Vet. Pathol. 2014, 51, 549–559. [Google Scholar] [CrossRef]
- Machado, M.C.A.; Ocarino, N.M.; Serakides, R.; Moroz, L.R.; Sementilli, A.; Damasceno, K.A.; Cruz, C.H.; Estrela-Lima, A. Triple-Negative Mammary Carcinoma in Two Male Dogs. J. Vet. Diagn. Investig. 2020, 32, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Sassi, F.; Benazzi, C.; Castellani, G.; Sarli, G. Molecular-Based Tumour Subtypes of Canine Mammary Carcinomas Assessed by Immunohistochemistry. BMC Vet. Res. 2010, 6, 5. [Google Scholar] [CrossRef]
- Varallo, G.; Gelaleti, G.; Maschio-Signorini, L.; Moschetta, M.; Lopes, J.; De Nardi, A.; Tinucci-Costa, M.; Rocha, R.; de Campos Zuccari, D. Prognostic Phenotypic Classification for Canine Mammary Tumors. Oncol. Lett. 2019, 8, 6545–6553. [Google Scholar] [CrossRef]
- Jaillardon, L.; Loussouarn, D.; Abadie, J.; Siliart, B.; Campone, M.; Nguyen, F. Abstract 85: Prognostic Stratification of Naturally Occurring Canine Triple Negative Invasive Mammary Carcinoma According to IGF1-R and AR Expression. In Proceedings of the Tumor Biology; American Association for Cancer Research, San Diego, CA, USA, 1 October 2014; p. 85. [Google Scholar]
- Kim, H.-W.; Lim, H.-Y.; Shin, J.-I.; Seung, B.-J.; Ju, J.-H.; Sur, J.-H. Breed- and Age-Related Differences in Canine Mammary Tumors. Can. J. Vet. Res. 2016, 80, 146–155. [Google Scholar]
- Kim, N.H.; Lim, H.Y.; Im, K.S.; Kim, J.H.; Sur, J.-H. Identification of Triple-Negative and Basal-like Canine Mammary Carcinomas Using Four Basal Markers. J. Comp. Pathol. 2013, 148, 298–306. [Google Scholar] [CrossRef]
- Chandra, M.; Riley, M.G.I.; Johnson, D.E. Spontaneous Neoplasms in Aged Sprague-Dawley Rats. Arch. Toxicol. 1992, 66, 496–502. [Google Scholar] [CrossRef] [PubMed]
- Rasotto, R.; Zappulli, V.; Castagnaro, M.; Goldschmidt, M.H. A Retrospective Study of Those Histopathologic Parameters Predictive of Invasion of the Lymphatic System by Canine Mammary Carcinomas. Vet. Pathol. 2012, 49, 330–340. [Google Scholar] [CrossRef]
- Walsh, K. Spontaneous Neoplasms in Control Wistar Rats. Fundam. Appl. Toxicol. 1994, 22, 65–72. [Google Scholar] [CrossRef]
- Russo, J.; Russo, I.H. Atlas and Histologic Classification of Tumors of the Rat Mammary Gland. J. Mammary Gland Biol. Neoplasia 2000, 5, 187–200. [Google Scholar] [CrossRef]
- Alvarado, A.; Faustino-Rocha, A.I.; Ferreira, R.; Mendes, R.; Duarte, J.A.; Pires, M.J.; Colaço, B.; Oliveira, P.A. Prognostic Factors in an Exercised Model of Chemically-Induced Mammary Cancer. Anticancer Res. 2016, 36, 2181–2188. [Google Scholar]
- Pinho, S.S.; Carvalho, S.; Cabral, J.; Reis, C.A.; Gärtner, F. Canine Tumors: A Spontaneous Animal Model of Human Carcinogenesis. Transl. Res. 2012, 159, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.; von Euler, H. Molecular Biological Aspects on Canine and Human Mammary Tumors. Vet. Pathol. 2011, 48, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Queiroga, F.L.; Raposo, T.; Carvalho, M.I.; Prada, J.; Pires, I. Canine Mammary Tumours as a Model to Study Human Breast Cancer: Most Recent Findings. In Vivo 2011, 25, 455–465. [Google Scholar]
- Murphy, S. Mammary Tumours in Dogs and Cats. InPractice 2008, 30, 334–339. [Google Scholar] [CrossRef]
- Kristiansen, V.M.; Peña, L.; Díez Córdova, L.; Illera, J.C.; Skjerve, E.; Breen, A.M.; Cofone, M.A.; Langeland, M.; Teige, J.; Goldschmidt, M.; et al. Effect of Ovariohysterectomy at the Time of Tumor Removal in Dogs with Mammary Carcinomas: A Randomized Controlled Trial. J. Vet. Intern. Med. 2016, 30, 230–241. [Google Scholar] [CrossRef]
- Miller, J.L.; Bartlett, A.P.; Harman, R.M.; Majhi, P.D.; Jerry, D.J.; Van de Walle, G.R. Induced Mammary Cancer in Rat Models: Pathogenesis, Genetics, and Relevance to Female Breast Cancer. J. Mammary Gland Biol. Neoplasia 2022, 27, 185–210. [Google Scholar] [CrossRef] [PubMed]
- Dufour, C.R.; Xia, H.; B’chir, W.; Perry, M.-C.; Kuzmanov, U.; Gainullina, A.; Dejgaard, K.; Scholtes, C.; Ouellet, C.; Zuo, D.; et al. Integrated Multi-Omics Analysis of Adverse Cardiac Remodeling and Metabolic Inflexibility upon ErbB2 and ERRα Deficiency. Commun. Biol. 2022, 5, 955. [Google Scholar] [CrossRef] [PubMed]
- Ledet, M.M.; Harman, R.M.; Fan, J.C.; Schmitt-Matzen, E.; Diaz-Rubio, M.E.; Zhang, S.; Van de Walle, G.R. Secreted Sphingomyelins Modulate Low Mammary Cancer Incidence Observed in Certain Mammals. Sci. Rep. 2020, 10, 20580. [Google Scholar] [CrossRef] [PubMed]
- Alvarado, A.; Gil da Costa, R.M.; Faustino-Rocha, A.I.; Ferreira, R.; Lopes, C.; Oliveira, P.A.; Colaço, B. Effects of Exercise Training on Breast Cancer Metastasis in a Rat Model. Int. J. Exp. Pathol. 2017, 98, 40–46. [Google Scholar] [CrossRef]
- Ferreira, L.P.; Gaspar, V.M.; Mano, J.F. Design of Spherically Structured 3D in Vitro Tumor Models -Advances and Prospects. Acta Biomater. 2018, 75, 11–34. [Google Scholar] [CrossRef]
- Ajjarapu, S.M.; Tiwari, A.; Kumar, S. Applications and Utility of Three-Dimensional In Vitro Cell Culture for Therapeutics. Future Pharmacol. 2023, 3, 213–228. [Google Scholar] [CrossRef]
- Barbosa, M.A.G.; Xavier, C.P.R.; Pereira, R.F.; Petrikaitė, V.; Vasconcelos, M.H. 3D Cell Culture Models as Recapitulators of the Tumor Microenvironment for the Screening of Anti-Cancer Drugs. Cancers 2021, 14, 190. [Google Scholar] [CrossRef]
- Pampaloni, F.; Stelzer, E.; Masotti, A. Three-Dimensional Tissue Models for Drug Discovery and Toxicology. BIOT 2009, 3, 103–117. [Google Scholar] [CrossRef]
- Freires, I.A.; de Sardi, J.C.O.; de Castro, R.D.; Rosalen, P.L. Alternative Animal and Non-Animal Models for Drug Discovery and Development: Bonus or Burden? Pharm. Res. 2017, 34, 681–686. [Google Scholar] [CrossRef]
- Doke, S.K.; Dhawale, S.C. Alternatives to Animal Testing: A Review. Saudi Pharm. J. 2015, 23, 223–229. [Google Scholar] [CrossRef]
- Kapałczyńska, M.; Kolenda, T.; Przybyła, W.; Zajączkowska, M.; Teresiak, A.; Filas, V.; Ibbs, M.; Bliźniak, R.; Łuczewski, Ł.; Lamperska, K. 2D and 3D Cell Cultures—A Comparison of Different Types of Cancer Cell Cultures. Arch. Med. Sci. 2018, 14, 910–919. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.E. Is It Time for Reviewer 3 to Request Human Organ Chip Experiments Instead of Animal Validation Studies? Adv. Sci. 2020, 7, 2002030. [Google Scholar] [CrossRef] [PubMed]
- Inglebert, M.; Dettwiler, M.; Hahn, K.; Letko, A.; Drogemuller, C.; Doench, J.; Brown, A.; Memari, Y.; Davies, H.R.; Degasperi, A.; et al. A Living Biobank of Canine Mammary Tumor Organoids as a Comparative Model for Human Breast Cancer. Sci. Rep. 2022, 12, 18051. [Google Scholar] [CrossRef]
- Raffo-Romero, A.; Aboulouard, S.; Bouchaert, E.; Rybicka, A.; Tierny, D.; Hajjaji, N.; Fournier, I.; Salzet, M.; Duhamel, M. Establishment and Characterization of Canine Mammary Tumoroids for Translational Research. BMC Biol. 2023, 21, 23. [Google Scholar] [CrossRef] [PubMed]
- Guillen, K.P.; Fujita, M.; Butterfield, A.J.; Scherer, S.D.; Bailey, M.H.; Chu, Z.; DeRose, Y.S.; Zhao, L.; Cortes-Sanchez, E.; Yang, C.-H.; et al. A Human Breast Cancer-Derived Xenograft and Organoid Platform for Drug Discovery and Precision Oncology. Nat. Cancer 2022, 3, 232–250. [Google Scholar] [CrossRef]
- Nowak, Ł.; Krajewski, W.; Małkiewicz, B.; Szydełko, T.; Pawlak, A. Characteristics and Applications of Canine In Vitro Models of Bladder Cancer in Veterinary Medicine: An Up-to-Date Mini Review. Animals 2022, 12, 516. [Google Scholar] [CrossRef] [PubMed]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-Dimensional Cell Culture Systems and Their Applications in Drug Discovery and Cell-Based Biosensors. ASSAY Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef]
- Gutleb, H.N.R.; Gutleb, A.C. A Short History of the Consideration of Sex Differences in Biomedical Research—Lessons for the In Vitro Community from Animal Models and Human Clinical Trials. Altern. Lab. Anim. 2023, 51, 026119292311567. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of Cancer: The Next Generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Ferreira, L.P.; Gaspar, V.M.; Mendes, L.; Duarte, I.F.; Mano, J.F. Organotypic 3D Decellularized Matrix Tumor Spheroids for High-Throughput Drug Screening. Biomaterials 2021, 275, 120983. [Google Scholar] [CrossRef]
- Białkowska, K.; Komorowski, P.; Bryszewska, M.; Miłowska, K. Spheroids as a Type of Three-Dimensional Cell Cultures-Examples of Methods of Preparation and the Most Important Application. Int. J. Mol. Sci. 2020, 21, 6225. [Google Scholar] [CrossRef]
- Ngo, H.; Amartumur, S.; Tran, V.T.A.; Tran, M.; Diep, Y.N.; Cho, H.; Lee, L.P. In Vitro Tumor Models on Chip and Integrated Microphysiological Analysis Platform (MAP) for Life Sciences and High-Throughput Drug Screening. Biosensors 2023, 13, 231. [Google Scholar] [CrossRef] [PubMed]
- Pinto, B.; Henriques, A.C.; Silva, P.M.A.; Bousbaa, H. Three-Dimensional Spheroids as In Vitro Preclinical Models for Cancer Research. Pharmaceutics 2020, 12, 1186. [Google Scholar] [CrossRef]
- Jensen, C.; Teng, Y. Is It Time to Start Transitioning From 2D to 3D Cell Culture? Front. Mol. Biosci. 2020, 7, 33. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Organogenesis in a Dish: Modeling Development and Disease Using Organoid Technologies. Science 2014, 345, 1247125. [Google Scholar] [CrossRef] [PubMed]
- Andrew Octavian, S.; Ying Pei, W. Organoids as Reliable Breast Cancer Study Models: An Update. Int. J. Oncol. Res. 2018, 1, 8. [Google Scholar] [CrossRef]
- Devarasetty, M.; Mazzocchi, A.R.; Skardal, A. Applications of Bioengineered 3D Tissue and Tumor Organoids in Drug Development and Precision Medicine: Current and Future. BioDrugs 2018, 32, 53–68. [Google Scholar] [CrossRef]
- Gunti, S.; Hoke, A.T.K.; Vu, K.P.; London, N.R. Organoid and Spheroid Tumor Models: Techniques and Applications. Cancers 2021, 13, 874. [Google Scholar] [CrossRef] [PubMed]
- Kamatar, A.; Gunay, G.; Acar, H. Natural and Synthetic Biomaterials for Engineering Multicellular Tumor Spheroids. Polymers 2020, 12, 2506. [Google Scholar] [CrossRef] [PubMed]
- Osuna de la Peña, D.; Trabulo, S.M.D.; Collin, E.; Liu, Y.; Sharma, S.; Tatari, M.; Behrens, D.; Erkan, M.; Lawlor, R.T.; Scarpa, A.; et al. Bioengineered 3D Models of Human Pancreatic Cancer Recapitulate in Vivo Tumour Biology. Nat. Commun. 2021, 12, 5623. [Google Scholar] [CrossRef]
- Ng, S.; Tan, W.J.; Pek, M.M.X.; Tan, M.-H.; Kurisawa, M. Mechanically and Chemically Defined Hydrogel Matrices for Patient-Derived Colorectal Tumor Organoid Culture. Biomaterials 2019, 219, 119400. [Google Scholar] [CrossRef] [PubMed]
- Moura, B.S.; Monteiro, M.V.; Ferreira, L.P.; Lavrador, P.; Gaspar, V.M.; Mano, J.F. Advancing Tissue Decellularized Hydrogels for Engineering Human Organoids. Adv. Funct. Mater. 2022, 32, 2202825. [Google Scholar] [CrossRef]
- Ferreira, L.P.; Gaspar, V.M.; Mano, J.F. Decellularized Extracellular Matrix for Bioengineering Physiomimetic 3D in Vitro Tumor Models. Trends Biotechnol. 2020, 38, 1397–1414. [Google Scholar] [CrossRef]
- Cardoso, T.C.; Sakamoto, S.S.; Stockmann, D.; Souza, T.F.B.; Ferreira, H.L.; Gameiro, R.; Vieira, F.V.; Louzada, M.J.Q.; Andrade, A.L.; Flores, E.F. A Three-dimensional Cell Culture System as an in Vitro Canine Mammary Carcinoma Model for the Expression of Connective Tissue Modulators. Vet. Comp. Oncol. 2017, 15, 582–593. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feature | Woman | Female Dog | Female Rat |
|---|---|---|---|
| Number of lactiferous ducts | 10–25 lactiferous ducts | 6–16 lactiferous ducts | 1 lactiferous duct |
| Morphofunctional unit | Tubuloalveolar | Tubuloalveolar | Tubuloalveolar |
| Cells | Luminal and myoepithelial | Luminal and myoepithelial | Luminal and myoepithelial |
| Development of functional glandular unit | Mature only in pregnancy | Mature only in pregnancy | Mature only in pregnancy |
| Vascularization | Internal thoracic artery Superior thoracic artery Lateral thoracic artery Acromiothoracic artery Thoracodorsal artery Lateral branches of posterior intercostal artery [51,52] | Internal thoracic artery (M1 and M2) Cranial superficial epigastric artery (M3) Caudal superficial epigastric and external pudendal artery (M4 and M5) [15,38] | Superficial cervical, internal thoracic External thoracic and axillary artery (1st and second pairs) Iliolumbar, superficial epigastric and external pudendal arteries (third to sixth pairs) [18] |
| Lymphatic drainage | Axillary lymph nodes and internal mammary lymph nodes (the main) Interpectoral, internal thoracic, supraclavicular, and infraclavicular lymph nodes [48,52] | Axillary lymph nodes (M1, M2 and M3) Superficial inguinal lymph nodes (M3, M4 and M5) [60,61] | Proper axillary node (all pairs) Accessory axillary lymph nodes (1st and second pair) Superficial cervical lymph node (1st pair) Inguinal lymph nodes (fourth, fifth and sixth pairs) [62] |
| Nerves | Third to sixth intercostal nerves second to sixth thoracic intercostal nerves Supraclavicular nerves [52,66] | Fourth, fifth, and sixth thoracic ventral nerves (M1) Sixth and seventh thoracic ventral nerves (M2) Genitofemoral nerve (M3, M4 and M5) [15] | NR |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ferreira, T.; Gama, A.; Seixas, F.; Faustino-Rocha, A.I.; Lopes, C.; Gaspar, V.M.; Mano, J.F.; Medeiros, R.; Oliveira, P.A. Mammary Glands of Women, Female Dogs and Female Rats: Similarities and Differences to Be Considered in Breast Cancer Research. Vet. Sci. 2023, 10, 379. https://doi.org/10.3390/vetsci10060379
Ferreira T, Gama A, Seixas F, Faustino-Rocha AI, Lopes C, Gaspar VM, Mano JF, Medeiros R, Oliveira PA. Mammary Glands of Women, Female Dogs and Female Rats: Similarities and Differences to Be Considered in Breast Cancer Research. Veterinary Sciences. 2023; 10(6):379. https://doi.org/10.3390/vetsci10060379
Chicago/Turabian StyleFerreira, Tiago, Adelina Gama, Fernanda Seixas, Ana I. Faustino-Rocha, Carlos Lopes, Vítor M. Gaspar, João F. Mano, Rui Medeiros, and Paula A. Oliveira. 2023. "Mammary Glands of Women, Female Dogs and Female Rats: Similarities and Differences to Be Considered in Breast Cancer Research" Veterinary Sciences 10, no. 6: 379. https://doi.org/10.3390/vetsci10060379
APA StyleFerreira, T., Gama, A., Seixas, F., Faustino-Rocha, A. I., Lopes, C., Gaspar, V. M., Mano, J. F., Medeiros, R., & Oliveira, P. A. (2023). Mammary Glands of Women, Female Dogs and Female Rats: Similarities and Differences to Be Considered in Breast Cancer Research. Veterinary Sciences, 10(6), 379. https://doi.org/10.3390/vetsci10060379