Simple Summary
Lyme borreliosis (LB) is a zoonotic disease caused by the spirochete Borrelia burgdorferi sensu lato (s.l.) transmitted to humans by the bite of infected Ixodes ticks. Borrelia miyamotoi is a spirochete that causes relapsing fever (RF) and is genetically related to Borrelia burgdorferi s.l. It is the only Borrelia species in the RF group that can be spread by the Ixodes tick. However, there have been no reports of B. miyamotoi in Egypt, and the data on LB in camels is scarce. Therefore, the current study is the first molecular study for the detection of Borrelia spp. and Borrelia miyamotoi in camels and associated ticks in Egypt. Furthermore, the identification of tick species that feed on camels demonstrates the value of cox1-based molecular approaches for identifying tick species. Here, we provide the first insight into the Borrelia miyamotoi and B. afzelii found in Egyptian camels and related ticks. Thus, it is essential to comprehend the role of the host in transmission and to constantly monitor the emergence of new diseases in Egypt.
Abstract
Tick-borne diseases (TBDs) are emerging and re-emerging infections that have a worldwide impact on human and animal health. Lyme borreliosis (LB) is a severe zoonotic disease caused by the spirochete Borrelia burgdorferi sensu lato (s.l.) transmitted to humans by the bite of infected Ixodes ticks. Borrelia miyamotoi is a spirochete that causes relapsing fever (RF) and is genetically related to Borrelia burgdorferi s.l. However, there have been no reports of B. miyamotoi in Egypt, and the data on LB in camels is scarce. Thus, the present study was conducted to screen and genetically identify Borrelia spp. and B. miyamotoi in Egyptian camels and associated ticks using polymerase chain reaction (PCR). Methods: A total of 133 blood samples and 1596 adult hard ticks were collected from Camelus dromedaries at Cairo and Giza slaughterhouses in Egypt. Tick species were identified by examining their morphology and sequencing the cytochrome C oxidase subunit 1 (cox1) gene. Borrelia spp. was detected using nested PCR on the IGS (16S-23S) gene, and positive samples were genotyped using 16S rRNA and glpQ spp. genes specific for Borrelia burgdorferi and Borrelia miyamotoi, respectively. The positive PCR products were sequenced and analyzed by phylogenetic tree. Results: Analysis of the cox1 gene sequence revealed that the adult ticks belonged to three genera; Hyalomma (H), Amblyomma (Am), and Rhipicephalus (R), as well as 12 species, including H. dromedarii, H. marginatum, H. excavatum, H. anatolicum, R. annulatus, R. pulchellus, Am. testudinarium, Am. hebraeum, Am. lipidium, Am. variegatum, Am. cohaerens and Am. gemma. Borrelia spp. was found in 8.3% (11/133) of the camel blood samples and 1.3% (21/1596) of the ticks, respectively. Sequencing of the IGS (16S-23S) gene found that B. afzelii, detected from H. dromedarii and H. marginatum, and B. crocidurae, which belongs to the RF group, was detected from one blood sample. B. burgdorferi and B. miyamotoi were discovered in the blood samples and tick species. Phylogenetic analysis of the glpQ gene showed that the B. miyamotoi in this study was of the Asian and European types. Conclusions: These results suggest that the camels can be infected by Lyme borrelia and other Borrelia bacteria species. This study also provides the first insight into the presence of Borrelia miyamotoi and B. afzelii DNA in camels and associated ticks in Egypt.
1. Background
The world population of Camelus dromedaries (one-hump dromedary camel) is estimated to be over 30 million, with Africa and the Middle East having the largest populations [1]. They are necessary for milk, meat, leather, transportation, and entertainment including tourism in Egypt. As a result, they play a critical role in the socio-economic development of many countries [2,3]. Because these camels are susceptible to several infectious illnesses, eating camel meat or having contact with them represents a significant source of zoonotic disease [1,4]. Tick infestations and TBDs are the most serious threat to camel health, causing global financial losses due to the geographic expansion of their tick vectors [5,6,7]. Amblyomma, Haemaphysalis, Hyalomma, and Rhipicephalus are the four genera of hard ticks that affect domestic animals in Egypt [8,9,10]. Moreover, tick species that severely infest dromedary camels in Egypt include H. dromedarii, H. excavatum, H. marginatum, and H. impeltatum [10,11,12]. Ticks carry a wide range of zoonotic pathogens, including Borrelia spp., which cause Lyme borreliosis (LB) and relapsing fever (RF) in humans [13]. Nearly 20 species within the Borrelia burgdorferi (sensu lato) complex were included, nine of which are known to cause animal and human LB (i.e., Borrelia afzelii, Borrelia bavariensis, Borrelia bissettii, B. burgdorferi (s.s.), Borrelia garinii, Borrelia kurtenbachii, Borrelia lusitaniae, Borrelia spielmanii and Borrelia valaisiana) [14]. Lyme borreliosis, which is spread naturally by Ixodes spp. ticks causes serious illness in humans, ranging from relatively benign skin lesions to severe cardiac, rheumatic, and neurologic signs [15,16,17]. Borrelia burgdorferi s.l. is widespread and is maintained in nature by various arthropod vectors, mammalian, birds, rats, reptiles, and many other wild species [18,19,20].
Borrelia miyamotoi is a spirochete that is genetically related to Borrelia burgdorferi s.l. It is the only Borrelia species in the RF group that can be spread by the Ixodes tick [21,22,23]. Borrelia miyamotoi infection commonly manifests as a febrile illness accompanied by fatigue, headache, chills, myalgia, arthralgia, and nausea, with potentially fatal complications such as meningoencephalitis [22,24,25,26]. Ticks of all stages are possible vectors of B. miyamotoi because it has transstadial and transovarial transmission which is not the case with Lyme borrelia, indicating that the global distribution of B. miyamotoi-infected ticks may exceed that of B. burgdorferi-infected ticks [27,28]. B. miyamotoi has been divided into three groupings based on geographic areas and principal vector species: Asian (or Siberian) (transmitted by I. persulcatus and I. pavlovskyi), American (transmitted by I. scapularis and I. pacificus), and European (carried by I. ricinus) [29]; and a new fourth clade was recently found in I. ovatus ticks in Japan [30,31]. In Africa, RF is most common in the northern part of Africa and is caused by various Borrelia spp. such as B. hispanica, B. duttonii, and B. crocidurae [32]. However, a new species identified as B. miyamotoi has lately sparked renewed attention in this bacterial group [33]. These diseases are challenging to diagnose due to the nonspecific nature of the febrile illness, isolation difficulties, and cross-reactivity between serological techniques [12]. Therefore, it is critical to understand host transmission and to monitor for the emergence of new diseases [34]. Borrelia miyamotoi and other relapsing fever group members are distinguished by a glycerophosphodiester phosphodiesterase (glpQ) gene [22]. B. burgdorferi s.l. lacks the gene for glpQ; therefore, this gene is used to detect B. miyamotoi. In both B. burgdorferi s.l. and B. miyamotoi, an intergenic spacer (IGS) between the 16S and 23S genes is usually used to detect them [35,36].
There have been no reports of B. miyamotoi in Egypt, and information on LB in camels is limited. Thus, the present study was conducted to screen and genetically identify Borrelia spp. and B. miyamotoi in Egyptian camels and associated ticks using molecular methods. Furthermore, to investigate the status of hard tick species that infest Egyptian camels.
2. Methods
2.1. Sample Collection and Preparation
A total of 133 healthy one-humped camels (Camelus dromedarius) aged 3–5 years were investigated for tick infestation. Blood samples and hard ticks were collected from 50 and 83 healthy dromedary camels at Cairo and Giza slaughterhouses in Egypt, respectively. The study was conducted from February 2021 to November 2021. Blood samples (5 mL) were collected in tubes coated with EDTA from the jugular blood vessels of the examined camels, then transferred to the laboratory in an icebox and maintained at −20 °C until DNA extraction. A total of 1596 adult ticks (600 from Cairo and 996 from Giza) were carefully gathered from the camels, then transferred alive to the zoonoses laboratory. Adult ticks were washed twice in distilled water, dried with paper tissues, and the classification of all collected ticks to genus level was by morphological characteristics using a stereomicroscope and taxonomic keys [37].
2.2. Extraction of DNA from Blood Samples and Ticks
Morphologically identified ticks (3–5 ticks/genus) were crushed into small pieces in a mortar with liquid nitrogen, then DNA was extracted from the ticks and blood samples (200 µL) using a Thermo Scientific GeneJET Genomic DNA Purification Kit (ThermoFisher, Darmstadt, Germany) according to the manufacturer’s recommended protocol. Isolated material was stored at −20 °C until further molecular analysis.
2.3. Molecular Identification of Tick Species
Ticks were identified to species level by amplifying and sequencing an ~820 bp fragment of the cytochrome oxidase c subunit I (cox1) gene. Amplification conditioning was performed according to [38]. Table 1 lists the primers used in this study.

Table 1.
Primer sequences used in this study and PCR amplification conditions.
2.4. Molecular Identification of the Borrelia Species
A nested PCR to detect Borrelia spp. in ticks and camel blood samples was carried out using outer and inner primers to amplify the 16S-23S rRNA intergenic spacer (16S-23S IGS) [40], and the amplification was performed according to [36].
Positive samples of Borrelia spp. were examined for detection of B. burgdorferi using a conventional PCR with a primer set of BbF and BbR for the 16S rRNA gene, according to [39]. Positive samples were further processed for sequencing.
An additional nested PCR test targeting the glpQ gene was performed on the positive samples to confirm the presence of B. miyamotoi and to detect genospecies. PCR amplicons were sequenced unidirectionally using primer Q3 to confirm the presence of B. miyamotoi in PCR-positive ticks and blood DNA samples [35,36].
The primers and amplification conditions for each reaction are displayed in Table 1. For amplification reactions cosmo Taq DNA Polymerase master mix (Willowfort, UK) was used. Each set of reactions included a positive control and the negative control consisted of nuclease-free water added to the PCR mix instead of the DNA sample. PCR products were visualized on 1.5% agarose gels.
2.5. Sequencing and Phylogenetic Analysis
PCR and nested PCR products were purified using a QIAquick purification extraction kit (Qiagen, Hombrechtikon, Switzerland), and sequenced using the BigDye Terminator V3.1 sequencing kit (Applied Biosystems, Waltham, MA, USA). The sequences were assembled into contigs using ChromasPro software (ChromasPro 1.7, Technelysium Pty Ltd., Tewantin, Australia). Next the sequences were aligned with the reference sequences available in the GenBank by BLAST and analyzed using MEGA software X. The phylogenetic trees were constructed using a maximum likelihood (ML) algorithm in MEGA X software using 1000 bootstrap replicates.
3. Results
3.1. Tick Species
Table 2 shows that 1596 ticks were collected and identified from 133 one-humped camels at Cairo and Giza slaughterhouses. Ticks were collected from healthy camel heads, necks, forelegs, udders, abdomens, back legs, and tails. According to cox1 gene sequencing, the most common tick species belonged to the Hyalomma genus (85.1%). Among tick species, Hyalomma dromedarii was the most common species; it accounted for 880 (59.4%), followed by Hyalomma marginatum with 297 (20.1%), Amblyomma hebraeum with 165 (11.1%), H. excavatum with 115 (7.2%), H. anatolicum with 66 (4.4%), and Rhipicephalus annulatus with 23 (1.6%). A few Am. testudinarium 12 (0.8%), Am. lepidum 11 (0.7%), Am. variegatum 10 (0.7%), R. pulchellus 8 (0.5%), Am. cohaerens 6 (0.4%), and Am. gemma 3 (0.2%) were found.
Table 2.
Tick species collected from dromedary camels in Cairo and Giza slaughterhouses of Egypt.
3.2. Prevalence of the Borrelia Species in the Camel Blood Samples and Ticks
The camel blood samples and ticks were screened for the presence of Borrelia spp. using nested PCRs with the IGS gene and showed that the prevalence of Borrelia spp. in adult ticks was 1.3% (21/1596), whereas it was 8.3% (11/133) in the camel blood samples (Table 3). Sequencing of the amplicons revealed the presence of three Borrelia spp. in the examined ticks, B. afzelii and B. burgdorferi (these two genospecies belonged to the B. burgdorferi s. l. group), and B. miyamotoi (belonging to the RF group). While B. burgdorferi, B. miyamotoi, and B. crocidurae were detected in blood samples (Table 3), Borrelia afzelii was not found in blood samples but found in two tick species, while B. crocidurae was detected in only one blood sample.
Table 3.
Prevalence of Borrelia species in camel blood samples and ticks.
Borrelia burgdorferi was confirmed using the 16SrRNA, revealing that 9.1% (1/11) were positive in blood samples and 14.3% (3/21) were positive in ticks. A second nested PCR on positive blood and tick samples targeting the glpQ gene to detect B. miyamotoi revealed that B. miyamotoi had the highest prevalence in adult ticks [76.2% (16/21)] and camel blood [81.8% (9/11)] (Table 3).
Following the nested PCR and the amplicon sequencing of the IGS gene, Borrelia spp. were detected in H. dromedarii (11/880, 1.3%), R. annulatus (2/23, 8.7%), Am. lepidum (2/11, 18.1%), H. marginatum (1/297, 0.3%), Am. testudinarium (1/12, 8.3%), Am. hebraeum (1/165, 0.6%), Am. variegatum (2/10, 20%), and Am. cohaerens (1/6, 16.6%). Additionally, B. afzelii was found in H. dromedarii and H. marginatum (Table 4).
Table 4.
The prevalence of Borrelia spp., B. burgdorferi, B. miyamotoi and B. afzelii among the tick Species.
By targeting the glpQ gene, B. miyamotoi was detected in H. dromedarii (8/880, 0.9%), Am. hebraeum (1/165, 0.6%), Am. Lepidum (2/11, 18.1%), Am. cohaerens (1/6, 16.6%), Am. variegatum (2/10, 20%), and R. annulatus (2/23, 8.7%). While by targeting the 16S rRNA, B. burgdorferi was found in H. dromedarii (2/880, 0.2%) and Am. testudinarium (1/12, 8.3) (Table 4).
3.3. Phylogenetic analysis
Tick species were classified into three genera based on sequence analysis of the positive PCR products of the cox1 gene: Hyalomma, Amblyomma, and Rhipicephalus; this was validated using a sequence identity of 96 to 100% with tick species sequences in GenBank (Figure 1). The accession numbers of cox1 gene sequences of identified ticks were deposited in GenBank, listed in Table 5.
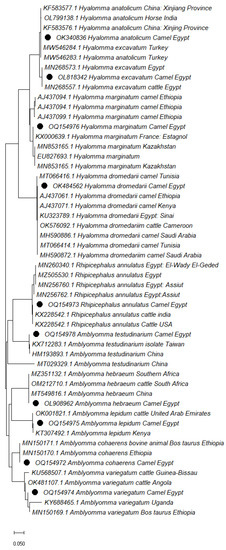
Figure 1.
Phylogenetic relationship of tick species collected from camels based on cytochrome c oxidase subunit 1 (cox1) gene. The accession numbers with black dots are from this study.
Table 5.
Accession numbers of Tick Species and Borrelia spp. in this study.
The phylogenetic tree of Borrelia spp. based on the 16S-23S IGS gene was consistent with the sequence results, identifying four genospecies found in ticks and blood; B. burgdorferi, B. afzelii, B. miyamotoi, and B. crocidurae. The B. miyamotoi found in Am. lepidum and Am. variegatum was identical to a German isolate (GenBank: MK945853.1, MK945806.1, MK945787.1, MK458687.1) and clustered with strains from France (MK732472.1) and from Sweden (MK458687.1) which was detected in human cerebrospinal fluid.
The Borrelia afzelii sequences in this study clustered in different branches and had significant similarities with German strains (GenBank: MK945805.1). Borrelia burgdorferi detected in Am. testudinarium clustered with strains detected in Ixodes pacificus (MN110090.1, MN110091.1, MN110092.1) in the USA. B. burgdorferi was simultaneously isolated from camel blood samples clustered with a strain isolated from humans (KM269456.1) in the USA. B. crocidurae found in camel blood clustered with other strains found in a person from France (LT984797.1) and a soft tick from Senegal (KF176328.1) (Figure 2).
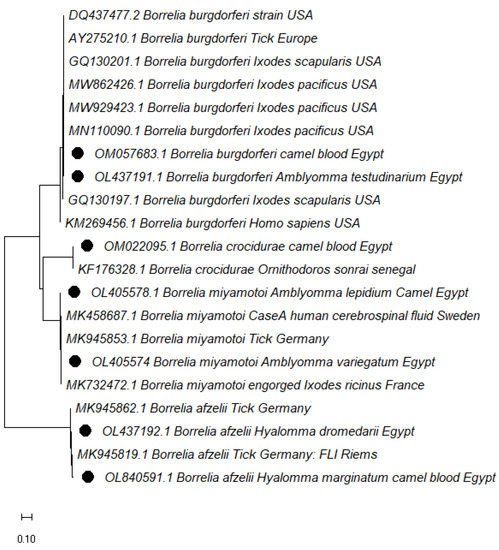
Figure 2.
Phylogenetic analysis using Maximum Likelihood method. The evolutionary history of Borrelia spp. isolates, based on the 16S-23S IGS. The accession numbers with black dots are from this study.
The presence of B. miyamotoi in ticks and camel blood samples was confirmed using glpQ sequence analysis (Figure 3). When compared to each other, most of the studied sequences were 100% identical. They shared a high level of similarity with B. miyamotoi sequences found in Ixodes persulcatus from Russia (LC538351.1), China (LC557152.1), and Japan (AB900798.1) (Asian type), as well as human blood from Russia (MK955928.1) and China (LC557152.1) (MW319188.1). The Borrelia miyamotoi sequence identified in Egyptian camels in Am. cohaerens was determined to be identical to B. miyamotoi sequences found in Ixodes ricinus from Italy (MG451835.1), and Hungary (MF678599.1), as well as in a human CSF fluid from Sweden (MK458689.1) (European type).

Figure 3.
Phylogenetic relationships based on the glpQ gene sequences of B. miyamotoi the trees were constructed and analyzed using the Maximum Likelihood method. A black dot indicates the new sequences provided by the present study.
Sequence analysis of B. burgdorferi 16S rRNA found in camel blood samples and Am. testudinarium from Egypt revealed 99% identity with B. burgdorferi found in Ixodes pacificus from the USA (KY563172.1) and R. sanguineus (MH685927.1) and canine blood from Egypt (MH685928.1) (Figure 4).

Figure 4.
Phylogenetic relationships based on the 16S rRNA gene sequences of B. burgdorferi. The trees were constructed and analyzed using the Maximum Likelihood method. The accession numbers with black dots are from this study.
A list of representative sequences with GenBank accession numbers and their source (tick species or blood) is given in (Table 5).
4. Discussion
Ticks and tick-borne diseases (TBDs) have emerged as major public health issues in many countries, including developed ones [41]. Tick infestation in camels has a high economic cost because ticks considerably impact their health and productivity [42]. As the prevalence of tick-borne diseases rises, distinguishing tick species is more critical than ever to improve tick and TBD control [43]. Standard morphological identification might be difficult in blood-engorged, immature, or physically injured specimens. As a result, molecular analysis can aid in discovering new information regarding ticks [44,45]. In this study, an assessment of 133 camels with an ixodid tick infestation revealed 12 species of ticks from 3 genera-infested dromedary camels. There were four species in the Hyalomma genus, six in the Amblyomma genus, and two in the Rhipicephalus genus. The genus Hyalomma was the most frequent tick genus in this study (85.1%). H. dromedarii is the most common tick species in Egyptian camels, whether locally raised or imported, according to previous studies [10,11,46,47]. H. dromedarii infection in camels is dangerous to their health, resulting in significant loss of camels and other animal products in the Middle East and North Africa [48,49], and impacts human health to some extent. Some of the tick species collected in this study (Am. lepidum, Am. variegatum, H. excavatum, R. pulchellus, and Am. gemma) are not indigenous to Egypt. This was observed in other studies [10,46,47]. These results might be attributed to camel imports to Egypt’s marketplaces coming from Sudan, Ethiopia, Nigeria, and Somalia [10], suggesting that these camels could be transporting non-endemic tick species to Egypt.
GenBank BLAST using the sequences of the cox1 genes, validated tick species identification for three genera: Hyalomma, Amblyomma, and Rhipicephalus, and was identical to tick species in the GenBank database. According to the phylogenetic analysis (Figure 1), the cox1 gene is a valuable and accurate tick species identification marker [50]. Lyme disease is one of the most serious zoonotic diseases, with endemic areas in Central Asia, the United States, and Eastern Europe [51]. This disease has been neglected in the Egyptian camel population.
In the present study, the prevalence rate of Borrelia spp. (B. burgdorferi, B. miyamotoi, and B. crocidurae) was detected in camel blood samples using nested PCRs based on the IGS (16S-23S rRNA) gene (8.3%, 11/133). This result was greater than that found in a study of dromedary camels from Tunisia (1.3%, 3/232) [52] and Bactrian camel blood examined in China (3.6%, 5/138) [20]. In contrast, Borrelia spirochete DNA was not found in Iran because the bacteria does not remain in the blood for prolonged periods after infection [53]. In addition, the total prevalence of Borrelia spp. in ticks was 1.3% (21/1596). This result was lower than the prevalence of Borrelia spp. in Ixodes ticks from northern Germany (31.6%, 3150 individual ticks) [54]. While in Ethiopian soft ticks the Prevalence of Borrelia spp. was 3.5% (11/312) [55].
Borrelia spp. was detected in H. dromedarii, R. annulatus, Am. lepidum, H. marginatum, Am. testudinarium, Am. hebraeum, Am. variegatum, and Am. cohaerens. This finding matches that of [56,57], who found B. burgdorferi in Egyptian Ixodid ticks (R. annulatus, H. dromedarii, H. excavatum, and R. sanguineus) and soft ticks (Ornithodoros savignyi). Borrelia spp. was detected in Egypt for the first time in Am. testudinarium, Am. cohaerens, Am. hebraeum, Am. variegatum, and Am. lepidum. This is comparable to a discovery from Korea [58], where they were the first to confirm B. burgdorferi in Am. testudinarium.
This investigation found Borrelia afzelii in H. dromedarii and H. marginatum. The B. afzelii sequences found in this study were very similar to isolated strains from Germany, where B. afzelii is the most common genospecies in Europe [59]. In Europe, rodents serve as reservoir hosts for B. afzelii (e.g., mice and voles) [60]. This infection causes Lyme disease, characterized by acrodermatitis chronica atrophicans and chronic skin disease [61].
While Borrelia crocidurae was found in one blood sample, it usually causes a tick-borne relapsing fever (TBRF) transmitted to humans through the bites of soft ticks of the genus Ornithodoros. Its impact on public health is just being realized, and it remains an unrecognized and neglected disease [62,63]. These findings suggest that these camels and associated ticks may be infected with Lyme borrelia and other Borrelia species, posing a risk to humans.
The presence of B. burgdorferi was confirmed using 16S rRNA gene sequence analysis. Only one sample of B. burgdorferi was detected in camel blood and isolated from two H. dromedarii and one Am. testudinarium. According to the phylogenetic analysis, the Egyptian and United States strains are related. This could be related to bird migration, which aids in the spread of Lyme disease by dispersing B. burgdorferi-infected ticks across the country and introduces a new endemic foci [64,65].
TBRF is underdiagnosed in tropical areas due to diagnostic confusion with malaria [66]. B. miyamotoi is a spirochete that causes a relapsing fever; spread worldwide by hard Ixodes tick species. In 1994, B. miyamotoi was isolated from Ixodes persulcatus and in 2011, human B. miyamotoi illness was first identified in Russia [67]. It is frequently recognized as a human pathogen [28,31]. However, no human B. miyamotoi infections have been documented in Egypt. In our study, B. miyamotoi was identified in 6.8% of the camel blood samples using the glpQ gene.
B. miyamotoi has been found in six hard tick species, including H. dromedarii, Am. hebraeum, Am. lipidium, Am. variegatum, Am. cohaerens and R. annulatus. The prevalence of B. miyamotoi in ticks (1%) is comparable to that discovered in Northeast China (1.3% of 774 ticks) with rates of 2.6% in I. persulcatus, 0.78% in Dermacentor nuttalli, 1.3% in D. silvarum, and 0.4% in Haemaphysalis longicornis. [28], but the prevalence is lower than that reported from Slovak Republic rodent-attached ticks (3.4%, 31/900) [68]. B. miyamotoi DNA was detected in the blood of one camel and in the H. dromedarii that infested this camel during our investigation. Sequencing using accession numbers OL347931 and OL439927 corroborated the findings. The results indicate that Borrelia was transmitted to the tick during camel bacteremia or that a Borrelia-infected tick transmitted the infection to this camel during a blood meal [69]. In the results of our study’s sequences of the glpQ gene, our B. miyamotoi was genotyped into Asian and European types and was highly similar to sequences of the gene amplified from ticks and human patients. Hence, individuals in Egypt are at risk of developing relapsing fever transmitted by ticks. Unfortunately, the role of camels is still unknown. Similarly, the competence of ticks as vectors for this pathogen needs to be confirmed. More research is needed to obtain more reliable information on the role of camels and associate ticks in pathogen transmission.
5. Conclusions
In conclusion, this study sheds light on the tick species that feed on camels in Egypt and demonstrates the value of cox1-based molecular approaches for identifying tick species. The camels in Egypt harbor several neglected, emerging, and re-emerging TBDs, many of which are likely new to Egypt, where Borrelia miyamotoi and Borrelia afzelii were detected for the first time in both camel blood and ticks. Future research needs to comprehend the role of camels in the enzootic cycle of Lyme borrelia.
Author Contributions
R.A., performed the collection of samples, the molecular and sequence analysis of the studied samples. R.A., D.H., M.K. and M.A.S. performed the analysis and interpretation of the data and the writing of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
The study was carried out according to the guide-lines of the ethical committee of the faculty of veterinary medicine, Cairo University (Institutional Animal Care and Use Committee), Vet CU. IACUC (Vet CU28/04/2021/303). All experimental protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the Faculty of Veterinary Medicine, Cairo University, Egypt.
Informed Consent Statement
Informed consent was obtained from all animal owners involved in the study.
Data Availability Statement
All the data generated or analyzed in this study are included in this published article.
Acknowledgments
The authors are grateful to Hend Abdullah (Researcher in National Research Centre, Egypt) for providing us with positive DNA controls required for PCR.
Conflicts of Interest
The authors declare that they have no competing interest.
References
- Zhu, S.; Zimmerman, D.; Deem, S.L. A review of zoonotic pathogens of dromedary camels. Ecohealth 2019, 16, 356–377. [Google Scholar] [CrossRef] [PubMed]
- Qamar, M.; Ayaz, M.; Nazir, M. Isolation and identification of ectoparasites in single humped camels (Camelus dromedarius) of Cholistan area, Pakistan. Iraqi J. Vet. Sci. 2019, 32, 291–297. [Google Scholar] [CrossRef]
- Nasirian, H. Detailed new insights about tick infestations in domestic ruminant groups: A global systematic review and meta-analysis. J. Parasit. Dis. 2022, 46, 526–601. [Google Scholar] [CrossRef] [PubMed]
- Bellabidi, M.; Benaissa, M.H.; Bissati-Bouafia, S.; Harrat, Z.; Brahmi, K.; Kernif, T. Coxiella burnetii in camels (Camelus dromedarius) from Algeria: Seroprevalence, molecular characterization, and ticks (Acari: Ixodidae) vectors. Acta Trop. 2020, 206, 105443. [Google Scholar] [CrossRef]
- Rochlin, I.; Toledo, A. Emerging tick-borne pathogens of public health importance: A mini-review. J. Med. Microbiol. 2020, 69, 781. [Google Scholar] [CrossRef] [PubMed]
- Ghafar, A.; Cabezas-Cruz, A.; Galon, C.; Obregon, D.; Gasser, R.B.; Moutailler, S.; Jabbar, A. Bovine ticks harbour a diverse array of microorganisms in Pakistan. Parasites Vectors 2020, 13, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Balinandi, S.; Mugisha, L.; Bbira, J.; Kabasa, W.; Nakayiki, T.; Bakkes, D.K.; Lutwama, J.J.; Chitimia-Dobler, L.; Malmberg, M. General and local morphological anomalies in Amblyomma lepidum (Acari: Ixodidae) and Rhipicephalus decoloratus infesting cattle in Uganda. J. Med. Entomol. 2019, 56, 873–877. [Google Scholar] [CrossRef]
- Allam, N.; El Moghazy, F.M.; Abdel-Baky, S. Molecular epidemiological updates on spotted fever rickettsioses in animal species and their hard ticks settling Egyptian desert. J. Adv. Pharm. Educ. Res. 2018, 8, 65. [Google Scholar]
- Mazyad, S.; Khalaf, S. Studies on theileria and babesia infecting live and slaughtered animals in Al Arish and El Hasanah, North Sinai Governorate, Egypt. J. Egypt. Soc. Parasitol. 2002, 32, 601–610. [Google Scholar]
- Okely, M.; Anan, R.; Gad-Allah, S.; Samy, A. Hard ticks (Acari: Ixodidae) infesting domestic animals in Egypt: Diagnostic characters and a Taxonomic key to the collected species. Med. Vet. Entomol. 2021, 35, 333–351. [Google Scholar] [CrossRef]
- Abdel-Shafy, S.; Allam, N.A.; Mediannikov, O.; Parola, P.; Raoult, D. Molecular detection of spotted fever group rickettsiae associated with ixodid ticks in Egypt. Vector-Borne Zoonotic Dis. 2012, 12, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, H.H.; Elbayoumy, M.K.; Allam, A.M.; Ashry, H.M.; Abdel-Shafy, S. Molecular epidemiology of certain vector-borne bacterial microorganisms in domestic animals and their ectoparasites in Egypt. Trop. Anim. Health Prod. 2021, 53, 484. [Google Scholar] [CrossRef] [PubMed]
- Replogle, A.J.; Sexton, C.; Young, J.; Kingry, L.C.; Schriefer, M.E.; Dolan, M.; Johnson, T.L.; Connally, N.P.; Padgett, K.A.; Petersen, J.M. Isolation of Borrelia miyamotoi and other Borreliae using a modified BSK medium. Sci. Rep. 2021, 11, 1926. [Google Scholar] [CrossRef]
- Mendoza-Roldan, J.A.; Colella, V.; Lia, R.P.; Nguyen, V.L.; Barros-Battesti, D.M.; Iatta, R.; Dantas-Torres, F.; Otranto, D. Borrelia burgdorferi (sensu lato) in ectoparasites and reptiles in southern Italy. Parasites Vectors 2019, 12, 1–9. [Google Scholar] [CrossRef]
- Wodecka, B.; Leońska, A.; Skotarczak, B. A comparative analysis of molecular markers for the detection and identification of Borrelia spirochaetes in Ixodes ricinus. J. Med. Microbiol. 2010, 59, 309–314. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H., Jr. Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick-Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef] [PubMed]
- Layzell, S.J.; Bailey, D.; Peacey, M.; Nuttall, P.A. Prevalence of Borrelia burgdorferi and Borrelia miyamotoi in questing Ixodes ricinus ticks from four sites in the UK. Ticks Tick-Borne Dis. 2018, 9, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Brisson, D.; Drecktrah, D.; Eggers, C.H.; Samuels, D.S. Genetics of Borrelia burgdorferi. Annu. Rev. Genet. 2012, 46, 515–536. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, S.A.; Bormane, A.; Dinnis, R.E.; Seelig, F.; Dobson, A.D.; Aanensen, D.M.; James, M.C.; Donaghy, M.; Randolph, S.E.; Feil, E.J. Host migration impacts on the phylogeography of Lyme Borreliosis spirochaete species in Europe. Environ. Microbiol. 2011, 13, 184–192. [Google Scholar] [CrossRef]
- Zhai, B.; Niu, Q.; Liu, Z.; Yang, J.; Pan, Y.; Li, Y.; Zhao, H.; Luo, J.; Yin, H. First detection and molecular identification of Borrelia species in Bactrian camel (Camelus bactrianus) from Northwest China. Infect. Genet. Evol. 2018, 64, 149–155. [Google Scholar] [CrossRef]
- Krause, P.J.; Narasimhan, S.; Wormser, G.P.; Rollend, L.; Fikrig, E.; Lepore, T.; Barbour, A.; Fish, D. Human Borrelia miyamotoi infection in the United States. N. Engl. J. Med. 2013, 368, 291. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.J.; Fish, D.; Narasimhan, S.; Barbour, A.G. Borrelia miyamotoi infection in nature and in humans. Clin. Microbiol. Infect. 2015, 21, 631–639. [Google Scholar] [CrossRef] [PubMed]
- Boyer, P.H.; Koetsveld, J.; Zilliox, L.; Sprong, H.; Talagrand-Reboul, É.; Hansmann, Y.; de Martino, S.J.; Boulanger, N.; Hovius, J.W.; Jaulhac, B. Assessment of Borrelia miyamotoi in febrile patients and ticks in Alsace, an endemic area for Lyme borreliosis in France. Parasites Vectors 2020, 13, 1–7. [Google Scholar] [CrossRef]
- Wagemakers, A.; Staarink, P.J.; Sprong, H.; Hovius, J.W. Borrelia miyamotoi: A widespread tick-borne relapsing fever spirochete. Trends Parasitol. 2015, 31, 260–269. [Google Scholar] [CrossRef]
- Page, S.; Daschkin, C.; Anniko, S.; Krey, V.; Nicolaus, C.; Maxeiner, H.-G. First report of Borrelia miyamotoi in an Ixodes ricinus tick in Augsburg, Germany. Exp. Appl. Acarol. 2018, 74, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Pukhovskaya, N.M.; Morozova, O.V.; Vysochina, N.P.; Belozerova, N.B.; Ivanov, L.I. Prevalence of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in ixodid ticks in the Far East of Russia. Int. J. Parasitol. Parasites Wildl. 2019, 8, 192–202. [Google Scholar] [CrossRef]
- Wormser, G.P.; McKenna, D.; Scavarda, C.; Cooper, D.; El Khoury, M.Y.; Nowakowski, J.; Sudhindra, P.; Ladenheim, A.; Wang, G.; Karmen, C.L. Co-infections in persons with early Lyme disease, New York, USA. Emerg. Infect. Dis. 2019, 25, 748. [Google Scholar] [CrossRef]
- Gao, Y.; Lv, X.-L.; Han, S.-Z.; Wang, W.; Liu, Q.; Song, M. First detection of Borrelia miyamotoi infections in ticks and humans from the northeast of Inner Mongolia, China. Acta Trop. 2021, 217, 105857. [Google Scholar] [CrossRef]
- Siński, E.; Welc-Falęciak, R.; Zajkowska, J. Borrelia miyamotoi: A human tick-borne relapsing fever spirochete in Europe and its potential impact on public health. Adv. Med. Sci. 2016, 61, 255–260. [Google Scholar] [CrossRef]
- Iwabu-Itoh, Y.; Bazartseren, B.; Naranbaatar, O.; Yondonjamts, E.; Furuno, K.; Lee, K.; Sato, K.; Kawabata, H.; Takada, N.; Andoh, M. Tick surveillance for Borrelia miyamotoi and phylogenetic analysis of isolates in Mongolia and Japan. Ticks Tick-Borne Dis. 2017, 8, 850–857. [Google Scholar] [CrossRef]
- Cutler, S.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. A new Borrelia on the block: Borrelia miyamotoi—A human health risk? Eurosurveillance 2019, 24, 1800170. [Google Scholar] [CrossRef] [PubMed]
- Trape, J.-F.; Diatta, G.; Arnathau, C.; Bitam, I.; Sarih, M.h.; Belghyti, D.; Bouattour, A.; Elguero, E.; Vial, L.; Mane, Y. The epidemiology and geographic distribution of relapsing fever borreliosis in West and North Africa, with a review of the Ornithodoros erraticus complex (Acari: Ixodida). PLoS ONE 2013, 8, e78473. [Google Scholar] [CrossRef] [PubMed]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergström, S.; Vial, L.; Boulanger, N. Relapsing fevers: Neglected tick-borne diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef]
- Madison-Antenucci, S.; Kramer, L.D.; Gebhardt, L.L.; Kauffman, E. Emerging tick-borne diseases. Clin. Microbiol. Rev. 2020, 33, e00083-00018. [Google Scholar] [CrossRef]
- Fomenko, N.; Livanova, N.; Borgoyakov, V.Y.; Kozlova, I.; Shulaykina, I.; Pukhovskaya, N.; Tokarevich, K.; Livanov, S.; Doroschenko, E.; Ivanov, L. Detection of Borrelia miyamotoi in ticks Ixodes persulcatus from Russia. Parasitology 2010, 44, 201–211. [Google Scholar]
- Reiter, M.; Schötta, A.-M.; Müller, A.; Stockinger, H.; Stanek, G. A newly established real-time PCR for detection of Borrelia miyamotoi in Ixodes ricinus ticks. Ticks Tick-Borne Dis. 2015, 6, 303–308. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A. Ticks of Domestic Animals in the Mediterranean Region: A Guide to Identification of Species; University of Zaragoza: Zaragoza, Spain, 2004. [Google Scholar]
- Abdullah, H.H.; El-Molla, A.; Salib, F.A.; Allam, N.A.; Ghazy, A.A.; Abdel-Shafy, S. Morphological and molecular identification of the brown dog tick Rhipicephalus sanguineus and the camel tick Hyalomma dromedarii (Acari: Ixodidae) vectors of Rickettsioses in Egypt. Vet. World 2016, 9, 1087. [Google Scholar] [CrossRef]
- Marconi, R.T.; Garon, C.F. Identification of a third genomic group of Borrelia burgdorferi through signature nucleotide analysis and 16S rRNA sequence determination. Microbiology 1992, 138, 533–536. [Google Scholar] [CrossRef]
- Bunikis, J.; Garpmo, U.; Tsao, J.; Berglund, J.; Fish, D.; Barbour, A.G. Sequence typing reveals extensive strain diversity of the Lyme borreliosis agents Borrelia burgdorferi in North America and Borrelia afzelii in Europe. Microbiology 2004, 150, 1741–1755. [Google Scholar] [CrossRef]
- Gabriele-Rivet, V.; Arsenault, J.; Badcock, J.; Cheng, A.; Edsall, J.; Goltz, J.; Kennedy, J.; Lindsay, L.R.; Pelcat, Y.; Ogden, N.H. Different ecological niches for ticks of public health significance in Canada. PLoS ONE 2015, 10, e0131282. [Google Scholar] [CrossRef]
- Nazifi, S.; Tamadon, A.; Behzadi, M.-A.; Haddadi, S.; Raayat-Jahromi, A.-R. One-humped camels (Camelus dromedaries) hard ticks infestation in Qeshm Island, Iran. Vet. Res. Forum 2011, 2, 135–138. [Google Scholar]
- Ganjali, M.; Dabirzadeh, M.; Sargolzaie, M. Species diversity and distribution of ticks (Acari: Ixodidae) in Zabol County, eastern Iran. J. Arthropod-Borne Dis. 2014, 8, 219. [Google Scholar] [PubMed]
- Li, L.-H.; Zhang, Y.; Wang, J.-Z.; Li, X.-S.; Yin, S.-Q.; Zhu, D.; Xue, J.-B.; Li, S.-G. High genetic diversity in hard ticks from a China-Myanmar border county. Parasites Vectors 2018, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasites Vectors 2014, 7, 1–11. [Google Scholar] [CrossRef]
- Barghash, S.; Hafez, A.; Darwish, A.; El-Naga, T. Molecular detection of pathogens in ticks infesting camels in Matrouh Governorate, Egypt. J. Bacteriol. Parasitol. 2016, 7, 259–262. [Google Scholar] [CrossRef]
- Hassan, M.I.; Gabr, H.S.; Abdel-Shafy, S.; Hammad, K.M.; Mokhtar, M.M. Prevalence of tick-vectors of Theileria annulata infesting the one-humped camels in Giza, Egypt. J. Egypt. Soc. Parasitol. 2017, 47, 425–432. [Google Scholar] [CrossRef]
- Alanazi, A.; Abdullah, S.; Helps, C.; Wall, R.; Puschendorf, R.; ALHarbi, S.; Abdel-Shafy, S.; Shaapan, R. Tick-borne pathogens in ticks and blood samples collected from camels in Riyadh Province, Saudi Arabia. Int. J. Zool. Res. 2018, 14, 30–36. [Google Scholar]
- Perveen, N.; Muzaffar, S.B.; Al-Deeb, M.A. Four tick-borne microorganisms and their prevalence in Hyalomma ticks collected from livestock in United Arab Emirates. Pathogens 2021, 10, 1005. [Google Scholar] [CrossRef]
- Amira, A.-H.; Răileanu, C.; Tauchmann, O.; Fischer, S.; Nijhof, A.M.; Silaghi, C. Tick species identification and molecular detection of tick-borne pathogens in blood and ticks collected from cattle in Egypt. Ticks Tick-Borne Dis. 2021, 12, 101676. [Google Scholar]
- Raza, N.; Durrani, A.Z.; Saleem, M.H.; Sheikh, A.A.; Usman, M.; Mujahid, Q.; Iqbal, M.Z.; Rizwan, M. Seroprevalence of Borrelia burgdorferi sensu lato in Camel (Camelus dromedarius) in Punjab, Pakistan. Pakistan J. Zool. 2021, 64, 1–4. [Google Scholar] [CrossRef]
- Said, M.B.; Belkahia, H.; Alberti, A.; Abdi, K.; Zhioua, M.; Daaloul-Jedidi, M.; Messadi, L. First molecular evidence of Borrelia burgdorferi sensu lato in goats, sheep, cattle and camels in Tunisia. Ann. Agric. Environ. Med. 2016, 23, 442–447. [Google Scholar] [CrossRef] [PubMed]
- Sazmand, A.; Harl, J.; Eigner, B.; Hodžić, A.; Beck, R.; Hekmatimoghaddam, S.; Mirzaei, M.; Fuehrer, H.-P.; Joachim, A. Vector-borne bacteria in blood of camels in Iran: New data and literature review. Comp. Immunol. Microbiol. Infect. Dis. 2019, 65, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Knoll, S.; Springer, A.; Hauck, D.; Schunack, B.; Pachnicke, S.; Fingerle, V.; Strube, C. Distribution of Borrelia burgdorferi sl and Borrelia miyamotoi in Ixodes tick populations in Northern Germany, co-infections with Rickettsiales and assessment of potential influencing factors. Med. Vet. Entomol. 2021, 35, 595–606. [Google Scholar] [CrossRef]
- Bankole, A.A.; Kumsa, B.; Mamo, G.; Ogo, N.I.; Elelu, N.; Morgan, W.; Cutler, S.J. Comparative Analysis of Tick-Borne Relapsing Fever Spirochaetes from Ethiopia and Nigeria. Pathogens 2023, 12, 81. [Google Scholar] [CrossRef] [PubMed]
- Adham, F.K.; El-Samie-Abd, E.M.; Gabre, R.M.; El Hussein, H. Detection of tick blood parasites in Egypt using PCR assay II-Borrelia burgdorferi sensu lato. J. Egypt. Soc. Parasitol 2010, 40, 553–564. [Google Scholar] [PubMed]
- Elhelw, R.A.; El-Enbaawy, M.I.; Samir, A. Lyme borreliosis: A neglected zoonosis in Egypt. Acta Trop. 2014, 140, 188–192. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Lee, S.-H.; Shin, S.; Kwak, D. Molecular Identification of Borrelia spp. from Ticks in Pastures Nearby Livestock Farms in Korea. Insects 2021, 12, 1011. [Google Scholar] [CrossRef]
- Răileanu, C.; Tauchmann, O.; Vasić, A.; Wöhnke, E.; Silaghi, C. Borrelia miyamotoi and Borrelia burgdorferi (sensu lato) identification and survey of tick-borne encephalitis virus in ticks from north-eastern Germany. Parasites Vectors 2020, 13, 106. [Google Scholar] [CrossRef]
- Kurtenbach, K.; De Michelis, S.; Etti, S.; Schäfer, S.M.; Sewell, H.-S.; Brade, V.; Kraiczy, P. Host association of Borrelia burgdorferi sensu lato–the key role of host complement. Trends Microbiol. 2002, 10, 74–79. [Google Scholar] [CrossRef]
- Wang, Y.; Li, S.; Wang, Z.; Zhang, L.; Cai, Y.; Liu, Q. Prevalence and identification of Borrelia burgdorferi sensu lato genospecies in ticks from Northeastern China. Vector-Borne Zoonotic Dis. 2019, 19, 309–315. [Google Scholar] [CrossRef]
- Parola, P.; Diatta, G.; Socolovschi, C.; Mediannikov, O.; Tall, A.; Bassene, H.; Trape, J.F.; Raoult, D. Tick-borne relapsing fever borreliosis, rural Senegal. Emerg. Infect. Dis. 2011, 17, 883. [Google Scholar] [CrossRef]
- Ndiaye, E.H.I.; Diouf, F.S.; Ndiaye, M.; Bassene, H.; Raoult, D.; Sokhna, C.; Parola, P.; Diatta, G. Tick-borne relapsing fever Borreliosis, a major public health problem overlooked in Senegal. PLoS Negl. Trop. Dis. 2021, 15, e0009184. [Google Scholar] [CrossRef]
- Reed, K.D.; Meece, J.K.; Henkel, J.S.; Shukla, S.K. Birds, migration and emerging zoonoses: West Nile virus, Lyme disease, influenza A and enteropathogens. Clin. Med. Res. 2003, 1, 5–12. [Google Scholar] [CrossRef]
- James, C.A. The Epidemiology of Ixodes scapularis and Borrelia burgdorferi Collected from Pet Dogs in an Emerging Lyme Disease Risk Area of Southeastern Ontario, Canada. Doctoral Dissertation, University of Guelph, Guelph, ON, Canada, 2017. [Google Scholar]
- Boulanger, N.; Boyer, P.; Talagrand-Reboul, E.; Hansmann, Y. Ticks and tick-borne diseases. Med. Et Mal. Infect. 2019, 49, 87–97. [Google Scholar] [CrossRef]
- Platonov, A.E.; Karan, L.S.; Kolyasnikova, N.M.; Makhneva, N.A.; Toporkova, M.G.; Maleev, V.V.; Fish, D.; Krause, P.J. Humans infected with relapsing fever spirochete Borrelia miyamotoi, Russia. Emerg. Infect. Dis. 2011, 17, 1816. [Google Scholar] [CrossRef]
- Heglasová, I.; Rudenko, N.; Golovchenko, M.; Zubriková, D.; Miklisová, D.; Stanko, M. Ticks, fleas and rodent-hosts analyzed for the presence of Borrelia miyamotoi in Slovakia: The first record of Borrelia miyamotoi in a Haemaphysalis inermis tick. Ticks Tick-Borne Dis. 2020, 11, 101456. [Google Scholar] [CrossRef]
- de la Fuente, J.; Contreras, M.; Estrada-Peña, A.; Cabezas-Cruz, A. Targeting a global health problem: Vaccine design and challenges for the control of tick-borne diseases. Vaccine 2017, 35, 5089–5094. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).