

Determining the In Vivo Efficacy of Plant-Based and Probiotic-Based Antibiotic Alternatives against Mixed Infection with Salmonella enterica and Escherichia coli in Domestic Chickens

 ,
,  , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets, and Experimental Design
2.2. Mode of Infection and Monitoring of Animals
2.3. Data Collection
2.4. Pathology and Pathological Histology
2.5. Statistical Method
3. Results
3.1. Preparation of the Challenge Bacteria
3.2. Mortality
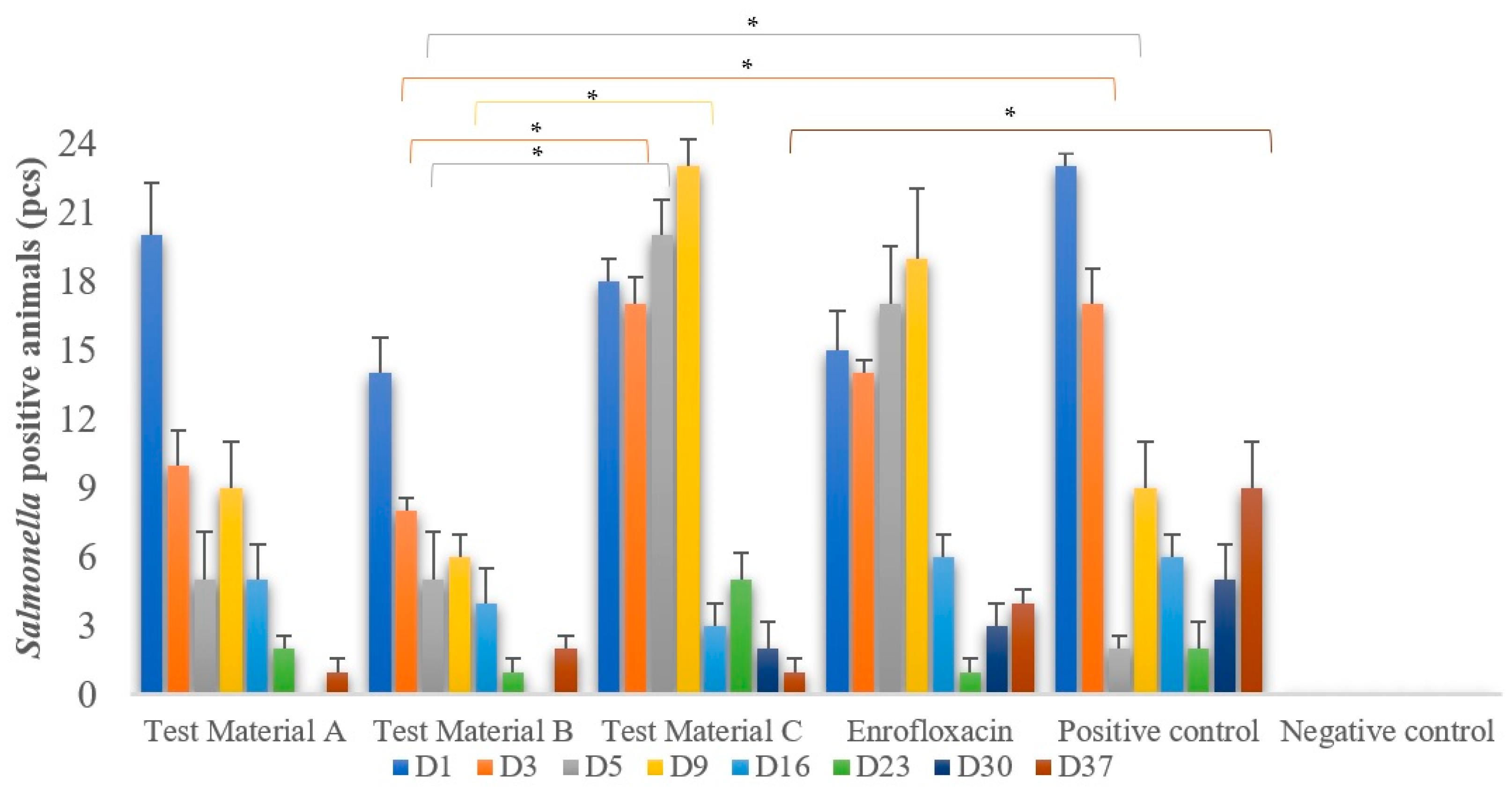
3.3. Salmonella Shedding
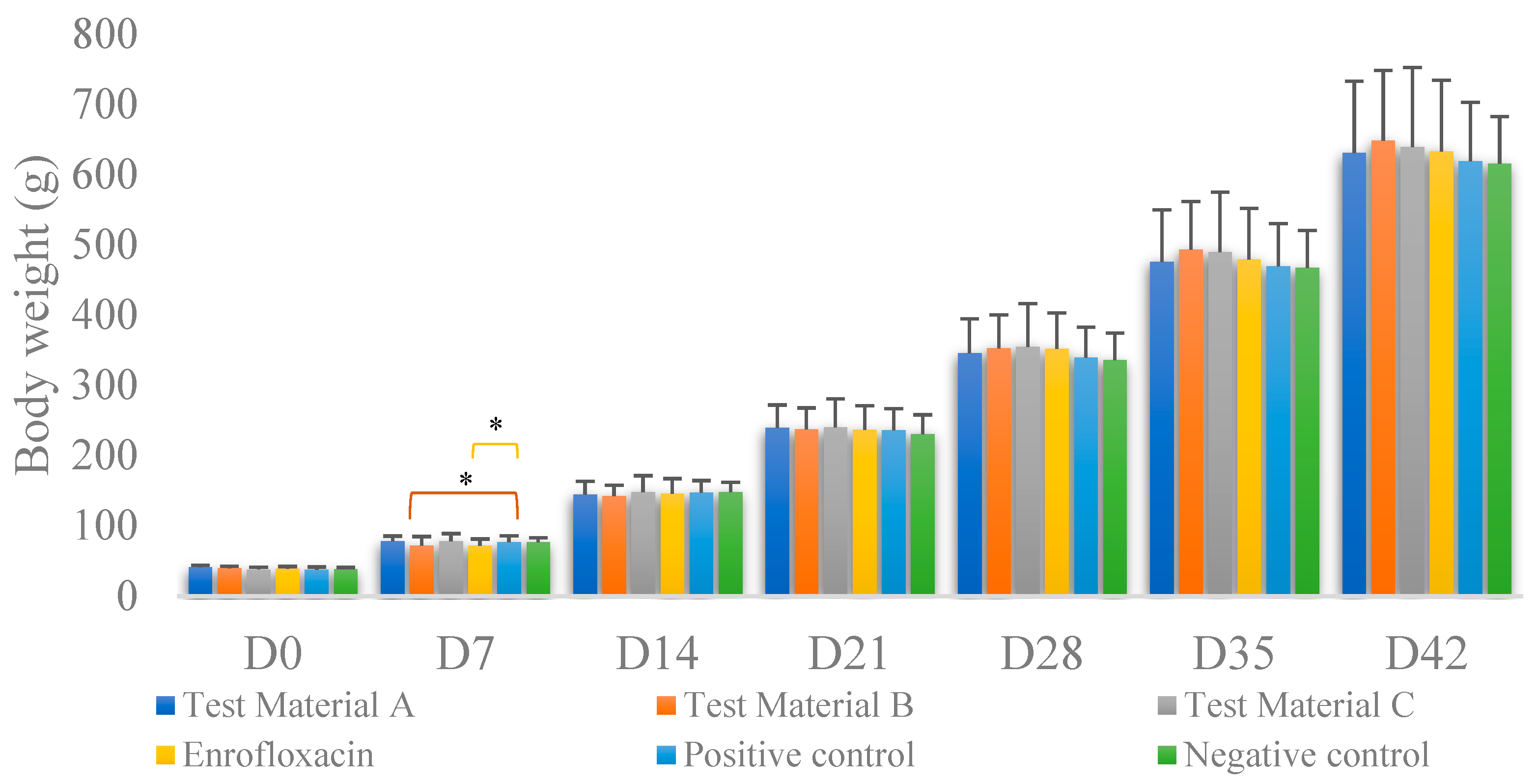
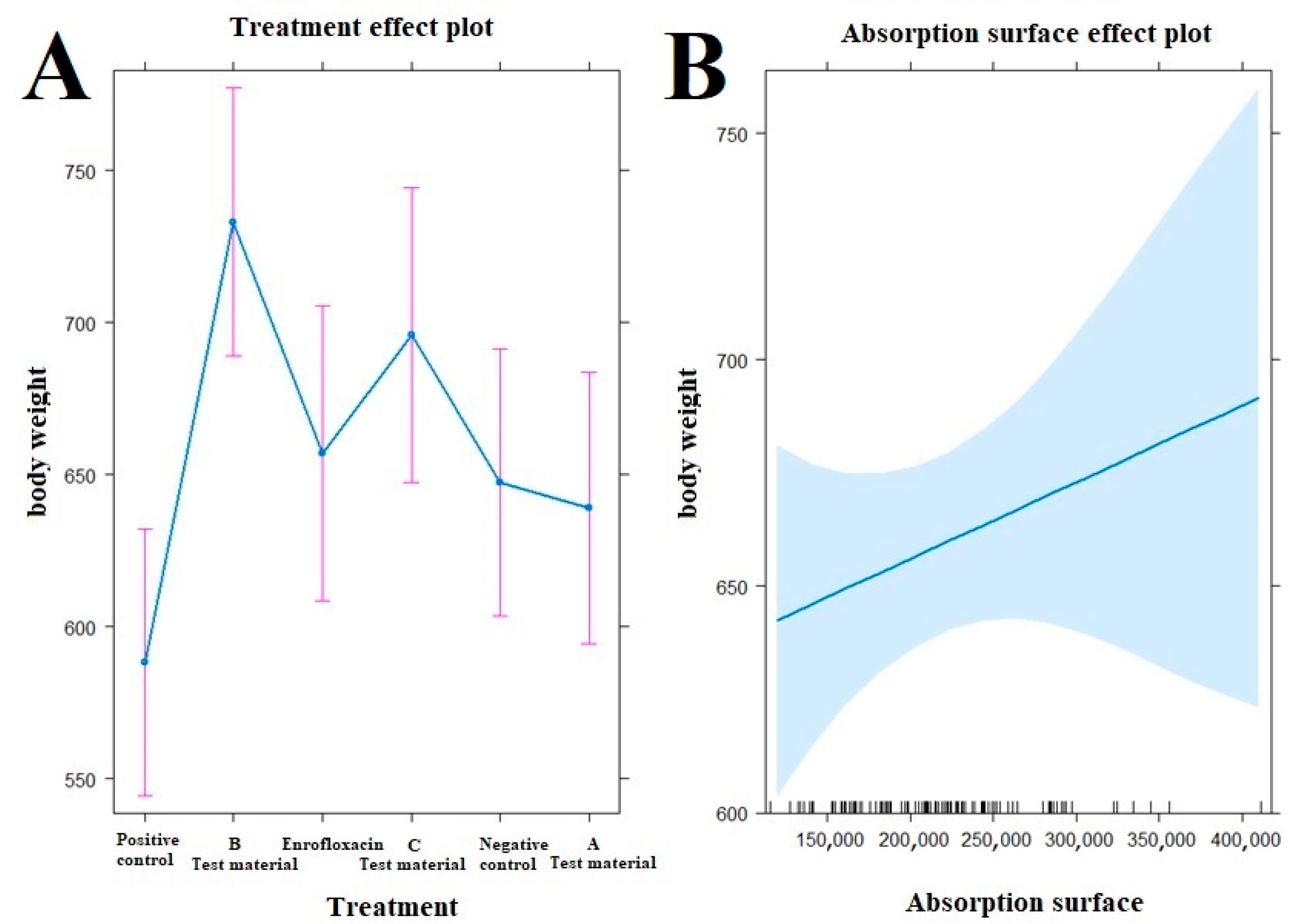
3.4. Body Weight
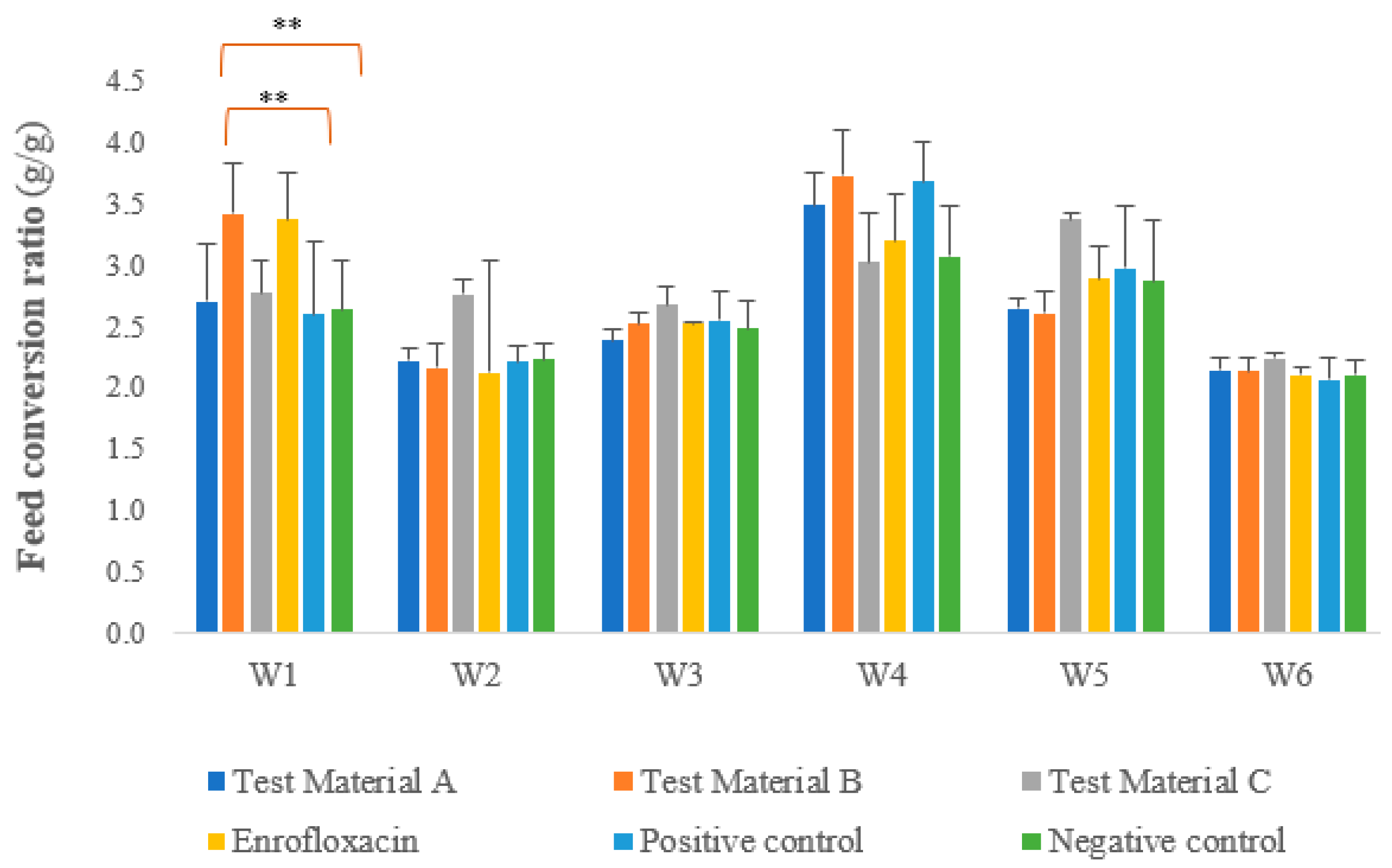
3.5. Trends in Feed Consumption and Feed Conversion Ratio
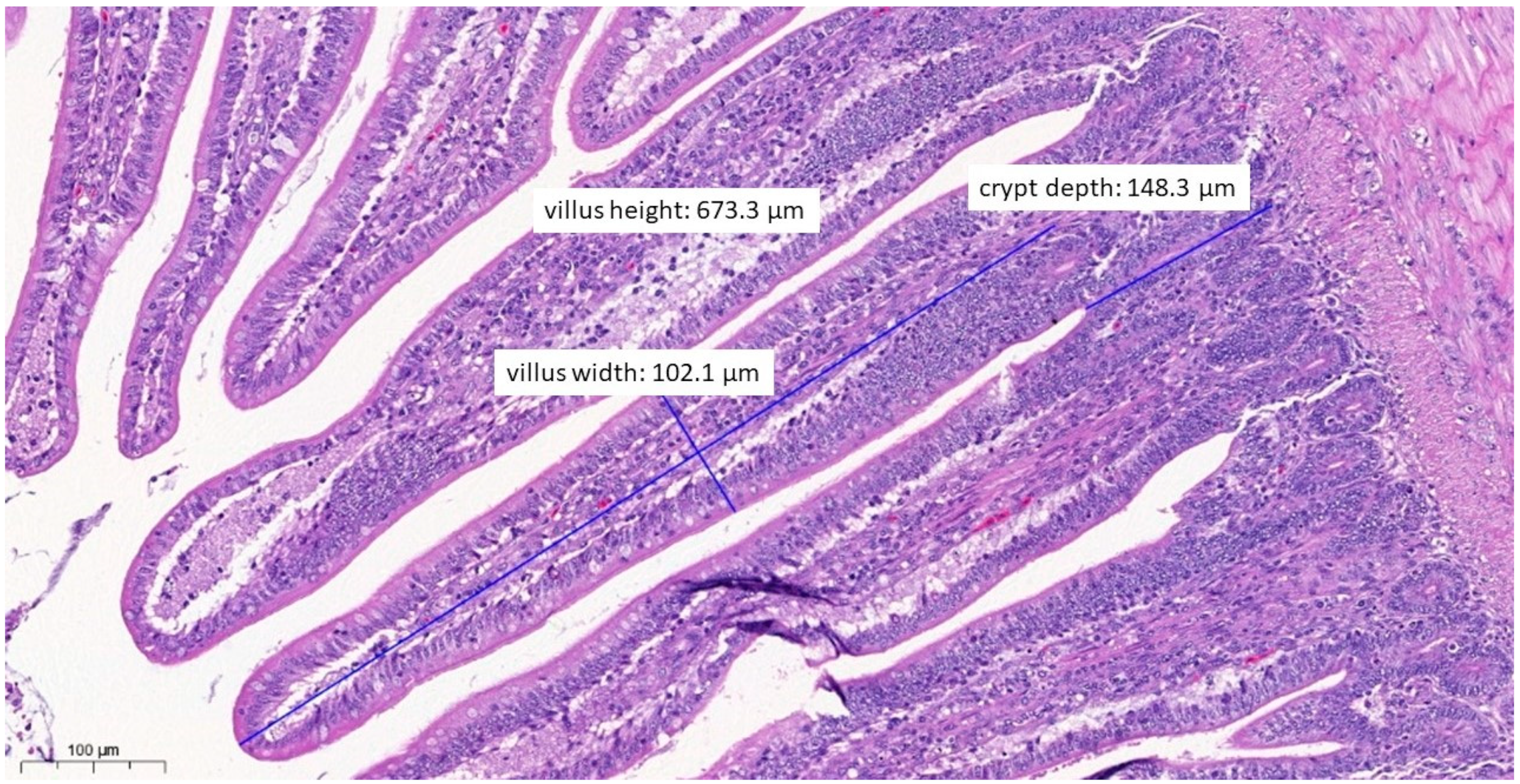
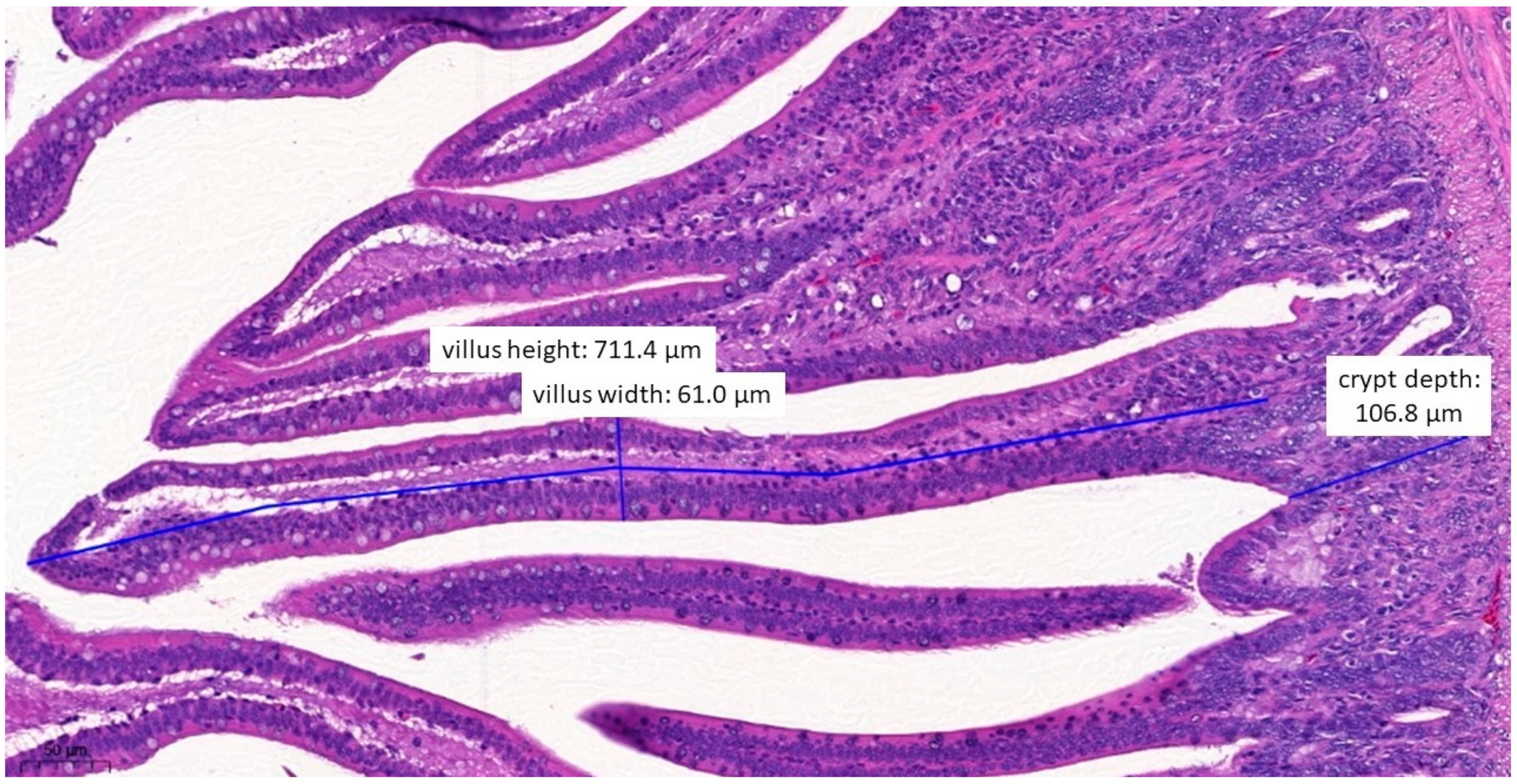
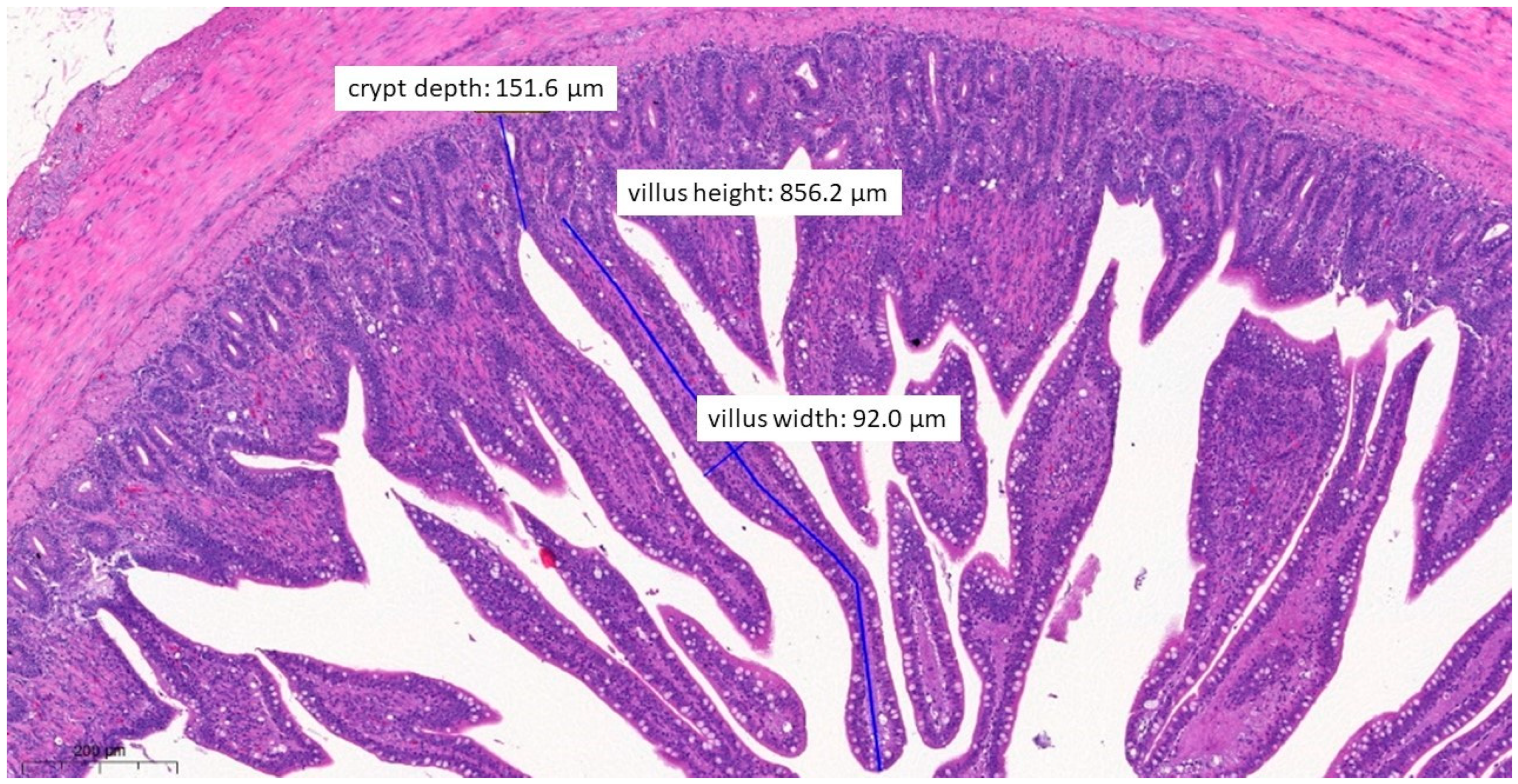
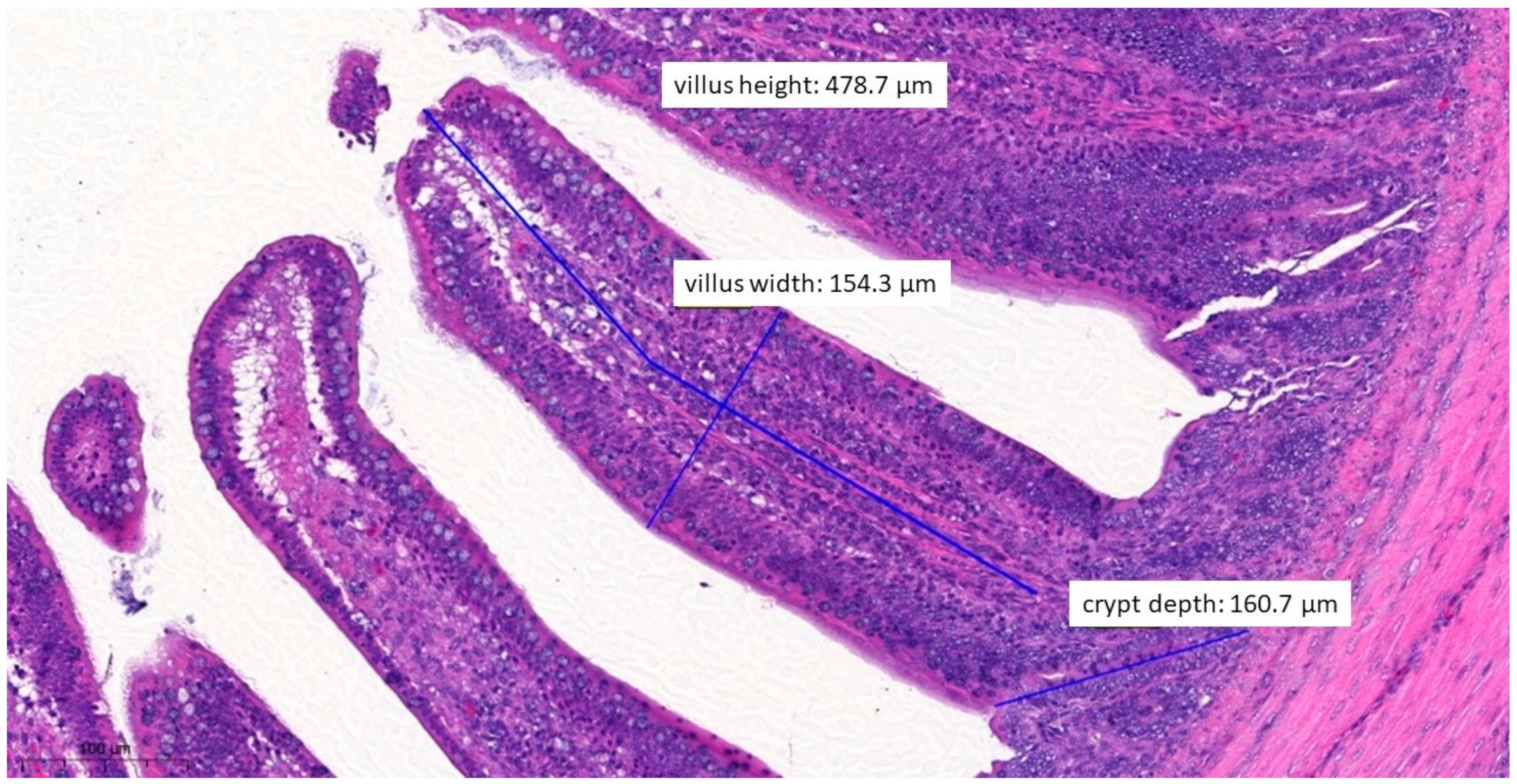
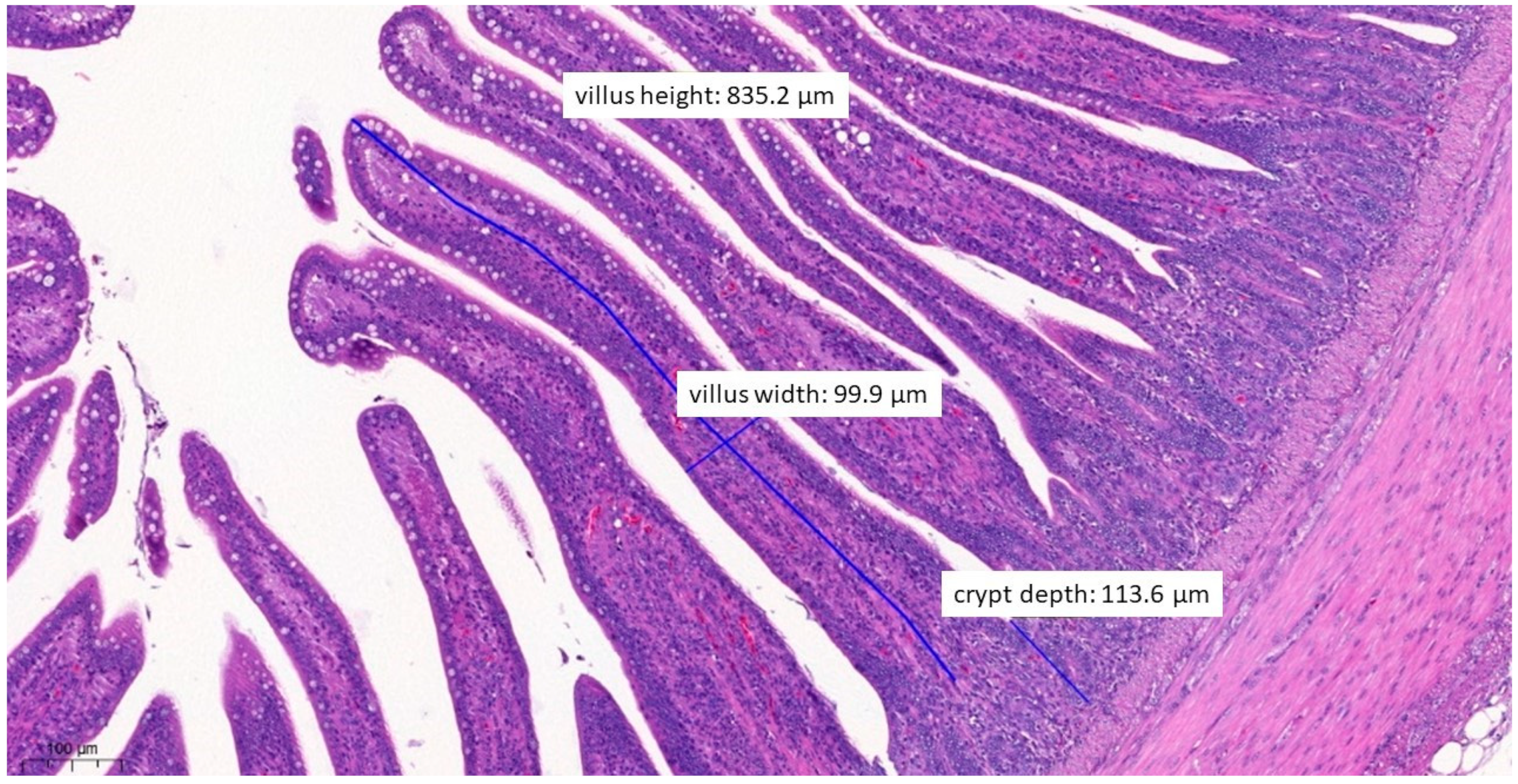
3.6. Pathological Histology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ghosh, D.; Veeraraghavan, B.; Elangovan, R.; Vivekanandan, P. Antibiotic resistance and epigenetics: More to it than meets the eye. Antimicrob. Agents Chemother. 2020, 64, e02225-19. [Google Scholar] [CrossRef]
- Baquero, F. Threats of antibiotic resistance: An obliged reappraisal. Int. Microbiol. 2021, 24, 499–506. [Google Scholar] [CrossRef]
- Mala, L.; Lalouckova, K.; Skrivanova, E. Bacterial skin infections in livestock and plant-based alternatives to their antibiotic treatment. Animals 2021, 11, 2473. [Google Scholar] [CrossRef] [PubMed]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef] [PubMed]
- Lamas, A.; Miranda, J.M.; Regal, P.; Vázquez, B.; Franco, C.M.; Cepeda, A. A Comprehensive review of non-enterica subspecies of Salmonella enterica. Microbiol. Res. 2018, 206, 60–73. [Google Scholar] [CrossRef]
- Feasey, N.A.; Dougan, G.; Kingsley, R.A.; Heyderman, R.S.; Gordon, M.A. Invasive non-typhoidal Salmonella disease: An emerging and neglected tropical disease in Africa. Lancet 2012, 379, 2489–2499. [Google Scholar] [CrossRef] [PubMed]
- El-Ghany, W.A.A. Salmonellosis: A food borne zoonotic and public health disease in Egypt. J. Infect. Dev. Ctries. 2020, 14, 674–678. [Google Scholar] [CrossRef]
- Sarowska, J.; Olszak, T.; Jama-Kmiecik, A.; Frej-Madrzak, M.; Futoma-Koloch, B.; Gawel, A.; Drulis-Kawa, Z.; Choroszy-Krol, I. Comparative characteristics and pathogenic potential of Escherichia coli isolates originating from poultry farms, retail meat, and human urinary tract infection. Life 2022, 12, 845. [Google Scholar] [CrossRef]
- Hu, J.; Afayibo, D.J.A.; Zhang, B.; Zhu, H.; Yao, L.; Guo, W.; Wang, X.; Wang, Z.; Wang, D.; Peng, H.; et al. Characteristics, pathogenic mechanism, zoonotic potential, drug resistance, and prevention of avian pathogenic Escherichia coli (APEC). Front. Microbiol. 2022, 13, 1049391. [Google Scholar] [CrossRef]
- Kromann, S.; Jensen, H.E. In vivo models of Escherichia coli infection in poultry. Acta Vet. Scand. 2022, 64, 33. [Google Scholar] [CrossRef]
- Wilczyński, J.; Stępień-Pyśniak, D.; Wystalska, D.; Wernicki, A. Molecular and serological characteristics of avian pathogenic Escherichia coli isolated from various clinical cases of poultry colibacillosis in Poland. Animals 2022, 12, 1090. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.J.; Nicolau, A.I.; Borda, D.; Nielsen, L.; Maia, R.L.; Møretrø, T.; Ferreira, V.; Knøchel, S.; Langsrud, S.; Teixeira, P. Salmonella in eggs: From shopping to consumption—A review providing an evidence-based analysis of risk factors. Compr. Rev. Food Sci. Food Saf. 2021, 20, 2716–2741. [Google Scholar] [CrossRef]
- Gast, R.K.; Jones, D.R.; Guraya, R.; Anderson, K.E.; Karcher, D.M. Research note: Contamination of eggs by Salmonella enteritidis and Salmonella typhimurium in experimentally infected laying hens in indoor cage-free housing. Poult. Sci. 2021, 100, 101438. [Google Scholar] [CrossRef]
- Olarte, J.; Galindo, E. Salmonella typhi resistant to chloramphenicol, ampicillin, and other antimicrobial agents: Strains isolated during an extensive typhoid fever epidemic in Mexico. Antimicrob. Agents Chemother. 1973, 4, 597–601. [Google Scholar] [CrossRef]
- Wain, J.; Hendriksen, R.S.; Mikoleit, M.L.; Keddy, K.H.; Ochiai, R.L. Typhoid fever. Lancet 2015, 385, 1136–1145. [Google Scholar] [CrossRef] [PubMed]
- Crump, J.A.; Sjölund-Karlsson, M.; Gordon, M.A.; Parry, C.M. Epidemiology, clinical presentation, laboratory diagnosis, antimicrobial resistance, and antimicrobial management of invasive Salmonella infections. Clin. Microbiol. Rev. 2015, 28, 901–937. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.S.; Rahman, S.R. Use of phages to treat antimicrobial-resistant Salmonella infections in poultry. Vet. Sci. 2022, 9, 438. [Google Scholar] [CrossRef]
- Palombo, E.A. Phytochemicals from traditional medicinal plants used in the treatment of diarrhoea: Modes of action and effects on intestinal function. Phytother. Res. 2006, 20, 717–724. [Google Scholar] [CrossRef]
- Schultes, R.E. The Kingdom of plants. In Medicines from the Earth; Thomson, W.A.R., Ed.; McGraw-Hill Book Co.: New York, NY, USA, 1978; 208p. [Google Scholar]
- Phanchana, M.; Harnvoravongchai, P.; Wongkuna, S.; Phetruen, T.; Phothichaisri, W.; Panturat, S.; Pipatthana, M.; Charoensutthivarakul, S.; Chankhamhaengdecha, S.; Janvilisri, T. Frontiers in antibiotic alternatives for Clostridioides difficile infection. World J. Gastroenterol. 2021, 27, 7210. [Google Scholar] [CrossRef]
- Kovács, L.; Nagy, D.; Könyves, L.; Jerzsele, Á.; Kerek, Á. Antimicrobial properties of essential oils—Animal health aspects. Magy. Állatorvosok Lapja 2023, 145, 497–510. [Google Scholar] [CrossRef]
- Kerek, Á.; Csanády, P.; Jerzsele, Á. Antibacterial efficiency of propolis—Part 1. Magy. Állatorvosok Lapja 2022, 144, 285–298. [Google Scholar]
- Kerek, Á.; Csanády, P.; Jerzsele, Á. Antiprotozoal and antifungal efficiency of propolis—Part 2. Magy. Állatorvosok Lapja 2022, 144, 691–704. [Google Scholar]
- Cowan, M.M. Plant Products as antimicrobial agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Sato, M.; Miyazaki, T.; Fujiwara, S.; Tanigaki, S.; Ohyama, M.; Tanaka, T.; Iinuma, M. Comparative study on the antibacterial activity of phytochemical flavanones against methicillin-resistant Staphylococcus aureus. J. Ethnopharmacol. 1996, 50, 27–34. [Google Scholar] [CrossRef]
- Gadde, U.; Kim, W.H.; Oh, S.T.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef]
- Várhidi, Z.; Máté, M.; Ózsvári, L. The use of probiotics in nutrition and herd health management in large hungarian dairy cattle farms. Front. Vet. Sci. 2022, 9, 957935. [Google Scholar] [CrossRef]
- Zhang, R.; Li, Z.; Gu, X.; Zhao, J.; Guo, T.; Kong, J. Probiotic Bacillus subtilis LF11 protects intestinal epithelium against Salmonella infection. Front. Cell Infect. Microbiol. 2022, 12, 837886. [Google Scholar] [CrossRef]
- Vandeplas, S.; Dauphin, R.D.; Beckers, Y.; Thonart, P.; Théwis, A. Salmonella in chicken: Current and developing strategies to reduce contamination at farm level. J. Food Prot. 2010, 73, 774–785. [Google Scholar] [CrossRef]
- Shao, Y.; Zhen, W.; Guo, F.; Hu, Z.; Zhang, K.; Kong, L.; Guo, Y.; Wang, Z. Pretreatment with probiotics Enterococcus faecium NCIMB 11181 attenuated Salmonella typhimurium-induced gut injury through modulating intestinal microbiome and immune responses with barrier function in broiler chickens. J. Anim. Sci. Biotechnol. 2022, 13, 130. [Google Scholar] [CrossRef]
- Lan, D.; Xun, X.; Hu, Y.; Li, N.; Yang, C.; Jiang, X.; Liu, Y. Research on the effect of Pediococcus pentosaceus on Salmonella enteritidis-infected chicken. Biomed. Res. Int. 2020, 2020, 6416451. [Google Scholar] [CrossRef] [PubMed]
- Tellez-Isaias, G.; Vuong, C.N.; Graham, B.D.; Selby, C.M.; Graham, L.E.; Señas-Cuesta, R.; Barros, T.L.; Beer, L.C.; Coles, M.E.; Forga, A.J.; et al. Developing probiotics, prebiotics, and organic acids to control Salmonella spp. in commercial turkeys at the University of Arkansas, USA. Ger. J. Vet. Res. 2021, 1, 7–12. [Google Scholar] [CrossRef]
- Shehata, A.; Attia, Y.; Khafaga, A.; Farooq, M.Z.; El-Seedi, H.; Eisenreich, W.; Tellez, G. Restoring healthy gut microbiome in poultry using alternative feed additives with particular attention to phytogenic substances: Challenges and prospects. Ger. J. Vet. Res. 2023, 2, 32–42. [Google Scholar] [CrossRef]
- Martin, K.; Laverty, L.; Filho, R.L.A.; Hernandez-Velasco, X.; Señas-Cuesta, R.; Gray, L.S.; Marcon, R.F.R.; Stein, A.; Coles, M.E.; Loeza, I.; et al. Evaluation of Aspergillus meal prebiotic in productive parameters, bone mineralization and intestinal integrity in broiler chickens. Ger. J. Vet. Res. 2023, 3, 27–33. [Google Scholar] [CrossRef]
- Almuzaini, A.M. Phytochemicals: Potential alternative strategy to fight Salmonella enterica serovar typhimurium. Front. Vet. Sci. 2023, 10, 1188752. [Google Scholar] [CrossRef] [PubMed]
- An Updated Review on Probiotics as an Alternative of Antibiotics in Poultry—A Review. Available online: https://www.animbiosci.org/journal/view.php?doi=10.5713/ab.21.0485 (accessed on 18 October 2023).
- Wu, Y.-T.; Yang, W.-Y.; Samuel Wu, Y.-H.; Chen, J.-W.; Chen, Y.-C. Modulations of growth performance, gut microbiota, and inflammatory cytokines by trehalose on Salmonella typhimurium-challenged broilers. Poult. Sci. 2020, 99, 4034–4043. [Google Scholar] [CrossRef] [PubMed]
- Prakatur, I.; Miskulin, M.; Pavic, M.; Marjanovic, K.; Blazicevic, V.; Miskulin, I.; Domacinovic, M. Intestinal morphology in broiler chickens supplemented with propolis and bee pollen. Animals 2019, 9, E301. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Reiczigel, J.; Harnos, A.; Solymosi, N. Biostatisztika Nem Statisztikusoknak, Javított Utánnyomás; Pars Kft.: Budapest, Hungary, 2014. [Google Scholar]
- Linear Mixed Model—An Overview|ScienceDirect Topics. Available online: https://www.sciencedirect.com/topics/mathematics/linear-mixed-model (accessed on 4 September 2023).
- Varmuzova, K.; Matulova, M.E.; Gerzova, L.; Cejkova, D.; Gardan-Salmon, D.; Panhéleux, M.; Robert, F.; Sisak, F.; Havlickova, H.; Rychlik, I. Curcuma and scutellaria plant extracts protect chickens against inflammation and Salmonella enteritidis infection. Poult. Sci. 2015, 94, 2049–2058. [Google Scholar] [CrossRef]
- Pascual, M.; Hugas, M.; Badiola, J.I.; Monfort, J.M.; Garriga, M. Lactobacillus salivarius CTC2197 prevents Salmonella enteritidis colonization in chickens. Appl. Environ. Microbiol. 1999, 65, 4981–4986. [Google Scholar] [CrossRef]
- Luoma, A.; Markazi, A.; Shanmugasundaram, R.; Murugesan, G.R.; Mohnl, M.; Selvaraj, R. Effect of synbiotic supplementation on layer production and cecal Salmonella load during a Salmonella challenge. Poult. Sci. 2017, 96, 4208–4216. [Google Scholar] [CrossRef]
- Fernandez, F.; Hinton, M.; Van Gils, B. Dietary mannan-oligosaccharides and their effect on chicken caecal microflora in relation to Salmonella enteritidis colonization. Avian Pathol. 2002, 31, 49–58. [Google Scholar] [CrossRef]
- Tabashsum, Z.; Peng, M.; Alvarado-Martinez, Z.; Aditya, A.; Bhatti, J.; Romo, P.B.; Young, A.; Biswas, D. Competitive reduction of poultry-borne enteric bacterial pathogens in chicken gut with bioactive Lactobacillus casei. Sci. Rep. 2020, 10, 16259. [Google Scholar] [CrossRef] [PubMed]
- Robinson, K.; Assumpcao, A.L.F.V.; Arsi, K.; Donoghue, A.; Jesudhasan, P.R.R. Ability of garlic and ginger oil to reduce Salmonella in post-harvest poultry. Animals 2022, 12, 2974. [Google Scholar] [CrossRef] [PubMed]
- Nair, A.; Balasaravanan, T.; Jadhav, S.; Mohan, V.; Kumar, C. Harnessing the antibacterial activity of Quercus infectoria and Phyllanthus emblica against antibiotic-resistant Salmonella typhi and Salmonella enteritidis of poultry origin. Vet. World 2020, 13, 1388–1396. [Google Scholar] [CrossRef] [PubMed]
- Al-Garadi, M.A.; Qaid, M.M.; Alqhtani, A.H.; Pokoo-Aikins, A.; Al-Mufarrej, S.I. In vitro phytochemical analysis and antibacterial and antifungal efficacy assessment of ethanolic and aqueous extracts of Rumex nervosus leaves against selected bacteria and fungi. Vet. World 2022, 15, 2725–2737. [Google Scholar] [CrossRef] [PubMed]
- El-Desoukey, R.M.A.; Albarakaty, F.M.; Alzamel, N.M.; AlZain, M.N. Ethnobotanical, phytochemical and antimicrobial activity of Halexylon salicornicum (Ramth) as a graze and Promising shrub against selected animal microbes. Saudi J. Biol. Sci. 2022, 29, 103328. [Google Scholar] [CrossRef] [PubMed]
- Kollanoor-Johny, A.; Mattson, T.; Baskaran, S.A.; Amalaradjou, M.A.; Babapoor, S.; March, B.; Valipe, S.; Darre, M.; Hoagland, T.; Schreiber, D.; et al. Reduction of Salmonella enterica serovar enteritidis colonization in 20-day-old broiler chickens by the plant-derived compounds trans-cinnamaldehyde and eugenol. Appl. Environ. Microbiol. 2012, 78, 2981–2987. [Google Scholar] [CrossRef] [PubMed]
- Kollanoor Johny, A.; Darre, M.J.; Donoghue, A.M.; Donoghue, D.J.; Venkitanarayanan, K. Antibacterial effect of trans-cinnamaldehyde, eugenol, carvacrol, and thymol on Salmonella enteritidis and Campylobacter jejuni in chicken cecal contents in vitro. J. Appl. Poult. Res. 2010, 19, 237–244. [Google Scholar] [CrossRef]
- Gurram, S.; Chinni Preetam, V.; Vijaya Lakshmi, K.; Raju, M.V.L.N.; Venkateswarlu, M.; Bora, S. Synergistic effect of probiotic, chicory root powder and coriander seed powder on growth performance, antioxidant activity and gut health of broiler chickens. PLoS ONE 2022, 17, e0270231. [Google Scholar] [CrossRef]
- Zou, Q.; Meng, W.; Li, C.; Wang, T.; Li, D. Feeding broilers with wheat germ, hops and grape seed extract mixture improves growth performance. Front. Physiol. 2023, 14, 1144997. [Google Scholar] [CrossRef]
- Leyva-Diaz, A.A.; Hernandez-Patlan, D.; Solis-Cruz, B.; Adhikari, B.; Kwon, Y.M.; Latorre, J.D.; Hernandez-Velasco, X.; Fuente-Martinez, B.; Hargis, B.M.; Lopez-Arellano, R.; et al. Evaluation of curcumin and copper acetate against Salmonella Typhimurium infection, intestinal permeability, and cecal microbiota composition in broiler chickens. J. Anim. Sci. Biotechnol. 2021, 12, 23. [Google Scholar] [CrossRef]
- Liu, H.Y.; Ivarsson, E.; Jönsson, L.; Holm, L.; Lundh, T.; Lindberg, J.E. Growth performance, digestibility, and gut development of broiler chickens on diets with inclusion of chicory (Cichorium intybus L.). Poult. Sci. 2011, 90, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Chen, L.; Zheng, K.; Ma, Y.-J.; He, R.-X.; Arowolo, M.A.; Zhou, Y.-J.; Xiao, D.-F.; He, J.-H. Effects of fenugreek seed extracts on growth performance and intestinal health of broilers. Poult. Sci. 2022, 101, 101939. [Google Scholar] [CrossRef] [PubMed]
- Jerzsele, Á.; Somogyi, Z.; Szalai, M.; Kovács, D. Effects of fermented wheat germ extract on artificial Salmonella typhimurium infection in broiler chickens. Magy. Állatorvosok Lapja 2020, 142, 77–85. [Google Scholar]
- Stipkovits, L.; Lapis, K.; Hidvégi, M.; Kósa, E.; Glávits, R.; Resetár, A. Testing the efficacy of fermented wheat germ extract against Mycoplasma gallisepticum infection of chickens. Poult. Sci. 2004, 83, 1844–1848. [Google Scholar] [CrossRef]
- Rafai, P.; Papp, Z.; Jakab, L.; Tuboly, T.; Jurkovich, V.; Brydl, E.; Ózsvári, L.; Kósa, E. The effect of fermented wheat germ extract on production parameters and immune status of growing pigs. J. Anim. Feed. Sci. 2011, 20, 36–46. [Google Scholar] [CrossRef]
- Khoobani, M.; Hasheminezhad, S.-H.; Javandel, F.; Nosrati, M.; Seidavi, A.; Kadim, I.T.; Laudadio, V.; Tufarelli, V. Effects of dietary chicory (Chicorium intybus L.) and probiotic blend as natural feed additives on performance traits, blood biochemistry, and gut microbiota of broiler chickens. Antibiotics 2019, 9, 5. [Google Scholar] [CrossRef]
- Kareem, K.Y.; Loh, T.C.; Foo, H.L.; Akit, H.; Samsudin, A.A. Effects of dietary postbiotic and inulin on growth performance, IGF1 and GHR mRNA expression, faecal microbiota and volatile fatty acids in broilers. BMC Vet. Res. 2016, 12, 163. [Google Scholar] [CrossRef]
- Olasz, Á.; Jerzsele, Á.; Balta, L.; Dobra, P.F.; Kerek, Á. In vivo efficacy of different extracts of propolis in broiler salmonellosis. Magy. Állatorvosok Lapja 2023, 145, 461–475. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kerek, Á.; Szabó, Á.; Dobra, P.F.; Bárdos, K.; Ózsvári, L.; Fehérvári, P.; Bata, Z.; Molnár-Nagy, V.; Jerzsele, Á. Determining the In Vivo Efficacy of Plant-Based and Probiotic-Based Antibiotic Alternatives against Mixed Infection with Salmonella enterica and Escherichia coli in Domestic Chickens. Vet. Sci. 2023, 10, 706. https://doi.org/10.3390/vetsci10120706
Kerek Á, Szabó Á, Dobra PF, Bárdos K, Ózsvári L, Fehérvári P, Bata Z, Molnár-Nagy V, Jerzsele Á. Determining the In Vivo Efficacy of Plant-Based and Probiotic-Based Antibiotic Alternatives against Mixed Infection with Salmonella enterica and Escherichia coli in Domestic Chickens. Veterinary Sciences. 2023; 10(12):706. https://doi.org/10.3390/vetsci10120706
Chicago/Turabian StyleKerek, Ádám, Ábel Szabó, Péter Ferenc Dobra, Krisztina Bárdos, László Ózsvári, Péter Fehérvári, Zsófia Bata, Viviána Molnár-Nagy, and Ákos Jerzsele. 2023. "Determining the In Vivo Efficacy of Plant-Based and Probiotic-Based Antibiotic Alternatives against Mixed Infection with Salmonella enterica and Escherichia coli in Domestic Chickens" Veterinary Sciences 10, no. 12: 706. https://doi.org/10.3390/vetsci10120706
APA StyleKerek, Á., Szabó, Á., Dobra, P. F., Bárdos, K., Ózsvári, L., Fehérvári, P., Bata, Z., Molnár-Nagy, V., & Jerzsele, Á. (2023). Determining the In Vivo Efficacy of Plant-Based and Probiotic-Based Antibiotic Alternatives against Mixed Infection with Salmonella enterica and Escherichia coli in Domestic Chickens. Veterinary Sciences, 10(12), 706. https://doi.org/10.3390/vetsci10120706