Wine Contamination with Ochratoxins: A Review




Abstract
1. Introduction
2. Ochratoxigenic Species on Grapes
3. Factors Affecting Ochratoxin A Presence on Grapes, Musts and Wines
3.1. Location (Latitude)
3.2. Biotic Factors
3.3. Weather
3.4. Period of Harvest
3.5. Agricultural and Harvesting Practices
3.6. Type of Wine
4. Fate of Ochratoxin A during Winemaking
5. Worldwide Regulations of Ochratoxin A in Wine and Grape Juice
6. Control and Detoxification Methods to Reduce OTA Content in Wine
7. Analytical Methods for OTA Determination in Wine
8. Other Mycotoxins in Wine
9. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Gil-Serna, J.; Vázquez, C.; Sardiñas, N.; González-Jaén, M.T.; Patiño, B. Revision of ochratoxin A production capacity by the main species of Aspergillus section Circumdati. Aspergillus steynii revealed as the main risk of OTA contamination. Food Control 2011, 22, 343–345. [Google Scholar] [CrossRef]
- El Khoury, A.; Atoui, A. Ochratoxin A: General overview and actual molecular status. Toxins 2010, 2, 461–493. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, J.D.; O’Keeffe, T.L. Detection and discrimination of four Aspergillus section Nigri species by PCR. Lett. Appl. Microbiol. 2015, 60, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Mycotoxins: Toxicology. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C., Tortorello, M.L., Eds.; Elsevier Ltd. Academic Press: Amsterdam, The Netherlands, 2014; pp. 887–892. ISBN 978-0-12-384733-1. [Google Scholar]
- International Agency for Research on Cancer (IARC). Some naturally occurring substances: Food items and constituents, heterocyclic aromatic amines and mycotoxins. IARC Monogr. Eval. Carcinog. Risks Hum. 1993, 56, 245–395. [Google Scholar]
- Zimmerli, B.; Dick, R. Ochratoxin A in table wine and grape juice: Occurrence and risk assessment. Food Addit. Contam. 1996, 13, 655–668. [Google Scholar] [CrossRef] [PubMed]
- Blesa, J.; Soriano, J.M.; Moltó, J.C.; Mañes, J. Factors affecting the presence of ochratoxin A in wines. Crit. Rev. Food Sci. 2006, 46, 473–478. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission Regulation EC No 1881/2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union 2006, 364, 5–24. [Google Scholar]
- Anli, R.E.; Bayram, M. Ochratoxin A in wines. Food Rev. Int. 2009, 25, 214–232. [Google Scholar] [CrossRef]
- Cabañes, F.J.; Accensi, F.; Bragulat, M.R.; Abarca, M.L.; Castellá, G.; Minguez, S.; Pons, A. What is the source of ochratoxin A in wine? Int. J. Food Microbiol. 2002, 79, 213–215. [Google Scholar] [CrossRef]
- Varga, J.; Kozakiewicz, Z. Ochratoxin A in grapes and grape derived products. Trends Food Sci. Technol. 2006, 17, 72–81. [Google Scholar] [CrossRef]
- Bau, M.; Bragulat, M.R.; Abarca, M.L.; Minguez, S.; Cabañes, F.J. Ochratoxigenic species from Spanish wine grapes. Int. J. Food Microbiol. 2005, 98, 125–130. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Bau, M.; Marín, S.; Abarca, M.L.; Ramos, A.J.; Bragulat, R. Mycobiota and ochratoxin A producing fungi from Spanish wine grapes. Int. J. Food Microbiol. 2006, 111, S40–S45. [Google Scholar] [CrossRef] [PubMed]
- García-Cela, E.; Crespo-Sempere, A.; Gil-Serna, J.; Porqueres, A.; Marín, S. Fungal diversity, incidence and mycotoxin contamination in grapes from two agro-climatic Spanish regions with emphasis on Aspergillus species. J. Sci. Food Agric. 2014, 95, 1716–1729. [Google Scholar] [CrossRef] [PubMed]
- Serra, R.; Abrunhosa, L.; Kozakiewicz, Z.; Venancio, A. Black Aspergillus species as ochratoxin A producers in Portuguese wine grapes. Int. J. Food Microbiol. 2003, 88, 63–68. [Google Scholar] [CrossRef]
- Serra, R.; Mendoça, C.; Venancio, A. Ochratoxin A occurrence and formation in Portuguese wine grapes at various stages of maturation. Int. J. Food Microbiol. 2006, 111, S35–S39. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Black Aspergilli and ochratoxin A production in French vineyards. Int. J. Food Microbiol. 2006, 111, S46–S52. [Google Scholar] [CrossRef] [PubMed]
- Dachoupakan, C.; Ratomahenina, R.; Martinez, V.; Guiraud, J.P.; Baccou, J.C.; Schorr-Galindo, S. Study of the phenotypic and genotypic biodiversity of potentially ochratoxigenic black aspergilli isolated from grapes. Int. J. Food Microbiol. 2009, 132, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Pietri, A.; Bertuzzi, T.; Languasco, L.; Giorni, P.; Kozakiewicz, Z. Occurrence of ochratoxin A—Producing fungi in grapes grown in Italy. J. Food Prot. 2003, 66, 633–636. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Giorni, P.; Bertuzzi, T.; Formenti, S.; Pietri, A. Black Aspergilli and ochratoxin A in grapes in Italy. Int. J. Food Microbiol. 2006, 111, S53–S60. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, S.E.; Antoniou, P.P.; Tjamos, E.C. Aspergillus spp., distribution, population composition and ochratoxin A production in wine producing vineyards in Greece. Int. J. Food Microbiol. 2006, 111, S61–S66. [Google Scholar] [CrossRef] [PubMed]
- Lasram, S.; Oueslati, S.; Mliki, A.; Ghorbel, A.; Silar, P.; Chebil, S. Ochratoxin A and ochratoxigenic black Aspergillus species in Tunisian grapes cultivated in different geographic areas. Food Control 2012, 25, 75–80. [Google Scholar] [CrossRef]
- Fredj, S.M.; Chebil, S.; Mliki, A. Isolation and characterization of ochratoxin A and aflatoxin B1 producing fungi infecting grapevines cultivated in Tunisia. Afr. J. Microbiol. Res. 2009, 3, 523–527. [Google Scholar]
- El Khoury, A.; Rizk, T.; Lteif, R.; Azouri, H.; Delia, M.L.; Lebrihi, A. Fungal contamination and aflatoxin B1 and ochratoxin A in Lebanese wine-grapes and musts. Food Chem. Toxicol. 2008, 46, 2244–2250. [Google Scholar] [CrossRef] [PubMed]
- Chulze, S.N.; Magnoli, C.E.; Dalcero, A.M. Occurrence of ochratoxin A in wine and ochratoxigenic mycoflora in grapes and dried vine fruits in South America. Int. J. Food Microbiol. 2006, 111, S5–S9. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Pitt, J.I.; Kazi, B.A.; Emmett, R.W.; Scott, E.S. Australian research on ochratoxigenic fungi and ochratoxin A. Int. J. Food Microbiol. 2006, 111, S10–S17. [Google Scholar] [CrossRef] [PubMed]
- Da Rocha, C.A.; Palacios, V.; Combina, M.; Fraga, M.E.; De Oliveira, A.; Magnoli, C.E.; Dalcero, A.M. Potential ochratoxin A producers from wine grapes in Argentina and Brazil. Food Addit. Contam. 2002, 19, 408–414. [Google Scholar] [CrossRef]
- Magnoli, C.; Violante, M.; Combina, M.; Palacio, G.; Dalcero, A. Mycoflora and ochratoxin-producing strains of Aspergillus section Nigri in wine grapes in Argentina. Lett. Appl. Microbiol. 2003, 37, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Chiotta, M.L.; Ponsone, M.L.; Combina, M.; Torres, A.M.; Chulze, S.N. Aspergillus section Nigri species isolated from different wine-grape growing regions in Argentina. Int. J. Food Microbiol. 2009, 136, 137–141. [Google Scholar] [CrossRef] [PubMed]
- Pantelides, I.S.; Aristeidou, E.; Lazari, M.; Tsolakidou, M.D.; Tsaltas, D.; Christofidou, M.; Kafouris, D.; Christou, E.; Ioannou, N. Biodiversity and ochratoxin A profile of Aspergillus section Nigri populations isolated from wine grapes in Cyprus vineyards. Food Microbiol. 2017, 67, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Mateo, R.; López-Ocaña, L.; Valle-Algarra, F.M.; Jiménez, M. Study of Spanish grape mycobiota and ochratoxin A production by isolates of Aspergillus tubingensis and other members of Aspergillus section Nigri. Appl. Environ. Microbiol. 2006, 71, 4696–4702. [Google Scholar] [CrossRef] [PubMed]
- Garmendia, G.; Vero, S. Occurrence and biodiversity of Aspergillus section Nigri on ‘Tannat’ grapes in Uruguay. Int. J. Food Microbiol. 2016, 216, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Qi, T.F.; Renaud, J.B.; McDowell, T.; Seifert, K.A.; Yeung, K.K.C.; Sumarah, M.W. Diversity of Mycotoxin-Producing Black Aspergilli in Canadian Vineyards. J. Agric. Food Chem. 2016, 64, 1583–1589. [Google Scholar] [CrossRef] [PubMed]
- Perrone, G.; Varga, J.; Susca, A.; Frisvad, J.C.; Stea, G.; Kocsube, S.; Kozakiewicz, Z.; Samson, R.A. Aspergillus uvarum sp. nov., an uniseriate black Aspergillus species isolated from grapes in Europe. Int. J. Syst. Evol. Microbiol. 2007, 58, 1032–1039. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.D.; Leong, S.L.; Kazi, B.A.; Emmett, R.W.; Scott, E.S. Fungi and mycotoxins in vineyards and grape products. Int. J. Food Microbiol. 2007, 119, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Valero, A.; Begum, M.; Leong, S.L.; Hocking, A.D.; Ramos, A.J.; Sanchis, V.; Marín, S. Effect of germicidal UVC light on fungi isolated from grapes and raisins. Lett. Appl. Microbiol. 2007, 45, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Bellí, N.; Marín, S.; Sanchis, V.; Ramos, A.J. Influence of water activity and temperature on growth of isolates of Aspergillus section Nigri obtained from grapes. Int. J. Food Microbiol. 2004, 96, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, D.; Parra, R.; Aldred, D.; Magan, N. Water and temperature relations of growth and ochratoxin A production by Aspergillus carbonarius strains from grapes in Europe and Israel. J. Appl. Microbiol. 2004, 97, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Scott, E.S. Effect of temperature and water activity on growth and ochratoxin A production by Australian Aspergillus carbonarius and A. niger isolates on a simulated grape juice medium. Int. J. Food Microbiol. 2006, 110, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Marín, S.; Bellí, N.; Lasram, S.; Chebil, S.; Ramos, A.J.; Ghorbel, A.; Sanchis, V. Kinetics of ochratoxin A production and accumulation by Aspergillus carbonarius on synthetic grape medium at different temperature levels. J. Food Sci. 2006, 71, M196–M200. [Google Scholar] [CrossRef]
- Lucchetta, G.; Bazzo, I.; Dal Cortivo, G.; Stringher, L.; Bellotto, D.; Borgo, M.; Angelini, E. Occurrence of black Aspergilli and ochratoxin A on grapes in Italy. Toxins 2010, 2, 840–855. [Google Scholar] [CrossRef] [PubMed]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Torres, A.; Knass, P.; Dalcero, A.; Chulze, S.N. Natural occurrence of ochratoxin A in musts, wines and grape vine fruits from grapes harvested in Argentina. Toxins 2010, 2, 1984–1996. [Google Scholar] [CrossRef] [PubMed]
- Hocking, A.D.; Varelis, P.; Pitt, J.I.; Cameron, S.F.; Leong, S.L. Occurrence of ochratoxin A in Australian wine. Aust. J. Grape Wine Res. 2003, 9, 72–78. [Google Scholar] [CrossRef]
- Ng, W.; Mankotia, M.; Pantazopoulos, P.; Neil, R.J.; Scott, P.M. Ochratoxin A in wine and grape juice sold in Canada. Food Addit. Contam. 2004, 21, 971–981. [Google Scholar] [CrossRef] [PubMed]
- Vega, M.; Ríos, G.; Baer, D.; Mardones, C.; Tessini, C.; Herlitz, E.; Saelzer, R.; Ruiz, M.A. Ochratoxin A occurrence in wines produced in Chile. Food Control 2012, 28, 147–150. [Google Scholar] [CrossRef]
- Wu, J.; Tan, Y.; Wang, Y.; Xu, R. Occurrence of ochratoxin A in wine and beer samples from China. Food Addit. Contam. B 2011, 4, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, L.; Li, J.; Zhu, B.; Ma, L. Occurrence of ochratoxin A in Chinese wines: Influence of local meteorological parameters. Eur. Food Res. Technol. 2013, 236, 277–283. [Google Scholar] [CrossRef]
- Mikulikova, R.; Belakova, S.; Benesoba, K.; Svoboda, Z. Study of ochratoxin A content in South Moravian and foreign wines by the UPLC method with fluorescence detection. Food Chem. 2012, 133, 55–59. [Google Scholar] [CrossRef]
- Burdaspal, P.; Legarda, T. Occurrence of ochratoxin A in sweet wines produced in Spain and other countries. Food Addit. Contam. 2007, 24, 976–986. [Google Scholar] [CrossRef] [PubMed]
- Stefanaki, I.; Foufa, E.; Tsatsou-Dritsa, A.; Dais, P. Ochratoxin A concentrations in Greek domestic wines and dried vine fruits. Food Addit. Contam. 2003, 20, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Soufleros, E.H.; Tricard, C.; Bouloumpasi, E.C. Occurrence of ochratoxin A in Greek wines. J. Sci. Food Agric. 2003, 83, 173–179. [Google Scholar] [CrossRef]
- Labrinea, E.P.; Natskoulis, P.I.; Spiropoulos, A.E.; Magan, N.; Tassou, C.C. A survey of ochratoxin A occurrence in Greek wines. Food Addit. Contam. B 2011, 4, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Varga, J.; Kiss, R.; Mátrai, T.; Mátrai, T.; Téren, J. Detection of ochratoxin A in Hungarian wines and beers. Acta Aliment. 2005, 34, 381–392. [Google Scholar] [CrossRef]
- Pietri, A.; Bertuzzi, T.; Pallaroni, L.; Piva, G. Occurrence of ochratoxin A in Italian wines. Food Addit. Contam. 2001, 18, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Spadaro, D.; Lorè, A.; Garibaldi, A.; Gullino, M.L. Occurrence of ochratoxin A before bottling in DOC and DOCG wines produced in Piedmont (Northern Italy). Food Control 2010, 21, 1294–1297. [Google Scholar] [CrossRef]
- Visconti, A.; Pascale, M.; Centonze, G. Determination of ochratoxin A in wine by means of immunoaffinity column clean-up and high-performance liquid chromatography. J. Chromatogr. A 1999, 864, 89–101. [Google Scholar] [CrossRef]
- Brera, C.; Debegnach, F.; Minardi, V.; Prantera, E.; Pannunzi, E.; Faleo, S.; De Santis, B.; Miraglia, M. Ochratoxin A contamination in Italian wine samples and evaluation of the exposure in the Italian population. J. Agric. Food Chem. 2008, 56, 10611–10618. [Google Scholar] [CrossRef] [PubMed]
- Di Stefano, V.; Pitonzo, R.; Avellone, G.; Di Fiore, A.; Monte, L.; Ogorka, A.Z. Determination of aflatoxins and ochratoxins in Sicilian Sweet Wines by High-Performance Liquid Chromatography with Fluorometric Detection and Immunoaffinity Cleanup. Food Anal. Method 2015, 8, 569–577. [Google Scholar] [CrossRef]
- Rotaru, S.; Israel-Roming, F.; Campeanu, G.; Deciu, G. Correlation of ochratoxin A level in wine with vine environment. Rom. Biotechnol. Lett. 2011, 16, 126–130. [Google Scholar]
- Bellí, N.; Marín, S.; Duaigües, A.; Ramos, A.J.; Sanchis, V. Ochratoxin A in wines, musts and grape juices from Spain. J. Sci. Food Agric. 2004, 84, 591–594. [Google Scholar] [CrossRef]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Occurrence of ochratoxin A in Rioja Alavesa wines. Food Chem. 2011, 126, 302–305. [Google Scholar] [CrossRef]
- Murillo-Arbizu, M.T.; Amézqueta, S.; González-Peñas, E.; López de Cerain, A. Occurrence of Ochratoxin A in Southern Spanish Generous Wines under the Denomination of Origin “Jerez-Xérès-Sherry and Manzanilla Sanlúcar de Barrameda”. Toxins 2010, 2, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Shepard, G.S.; Fabiani, A.; Stockenstro, S.; Mschicileli, N.; Sewram, V. Quantitation of ochratoxin A in South African wines. J. Agric. Food Chem. 2003, 51, 1102–1106. [Google Scholar] [CrossRef] [PubMed]
- Stander, M.A.; Steyn, P.S. Survey of ochratoxin A in South African wines. S. Afr. J. Enol. Vitic. 2002, 23, 9–13. [Google Scholar] [CrossRef]
- Var, I.; Kabak, B. Occurrence of ochratoxin A in Turkish wines. Microchem. J. 2007, 86, 241–247. [Google Scholar] [CrossRef]
- Grazioli, B.; Fumi, M.D.; Silva, A. The role of processing on ochratoxin A content in Italian must and wine: A study on naturally contaminated grapes. Int. J. Food Microbiol. 2006, 111, S93–S96. [Google Scholar] [CrossRef] [PubMed]
- Miraglia, M.; Brera, C. Task 3.2.7. Assessment of Dietary Intake of Ochratoxin A by the Population of EU Member States. 2002. Available online: https://ec.europa.eu/food/sites/food/files/safety/docs/cs_contaminants_catalogue_ochratoxin_task_3-2-7_en.pdf (accessed on 25 October 2017).
- Otteneder, H.; Majerus, P. Occurrence of ochratoxin A (OTA) in wines: Influence of the type of wine and its geographical origin. Food Addit. Contam. 2000, 17, 793–798. [Google Scholar] [CrossRef] [PubMed]
- Remiro, R.; Irigoyen, A.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Levels of ochratoxin in Mediterranean red wines. Food Control 2013, 32, 63–68. [Google Scholar] [CrossRef]
- Cozzi, G.; Pascale, M.; Perrone, G.; Visconti, A.; Logrieco, A. Effect of Lobesia botrana damages on black aspergilli rot and ochratoxin A content in grapes. Int. J. Food Microbiol. 2006, 111, S88–S92. [Google Scholar] [CrossRef] [PubMed]
- Covarelli, L.; Beccari, G.; Marini, A.; Tosi, L. A review on the occurrence and control of ochratoxigenic fungal species and ochratoxin A in dehydrated grapes, non-fortified dessert wines and dried vine fruit in the Mediterranean area. Food Control 2012, 26, 347–356. [Google Scholar] [CrossRef]
- Cozzi, G.; Haidukowski, M.; Perrone, G.; Visconti, A.; Logrieco, A. Influence of Lobesia botrana field control on black aspergilli rot and ochratoxin A contamination in grapes. J. Food Prot. 2009, 72, 894–897. [Google Scholar] [CrossRef] [PubMed]
- Cozzi, G.; Somma, S.; Haidukowski, M.; Logrieco, A. Ochratoxin A management in vineyards by Lobesia botrana biocontrol. Toxins 2013, 5, 49–59. [Google Scholar] [CrossRef] [PubMed]
- The Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Food Additives and Contaminants. Ochratoxin A. WHO Food Additives Series 2008, No. 59. Available online: http://apps.who.int/iris/bitstream/ 10665/43823/1/9789241660594_eng.pdf (accessed on 25 October 2017).
- López de Cerain, A.; González-Peñas, E.; Jiménez, A.M.; Bello, J. Contribution to the study of ochratoxin A in Spanish wines. Food Addit. Contam. 2002, 19, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Clouvel, P.; Bonvarlet, L.; Martínez, A.; Lagouarde, P.; Dieng, I.; Martin, P. Wine contamination by ochratoxin A in relation to vine environment. Int. J. Food Microbiol. 2008, 123, 74–80. [Google Scholar] [CrossRef] [PubMed]
- Paterson, M.; Venancio, A.; Lima, N.; Guilloux-Bénatier, M.; Rousseaux, S. Predominant mycotoxins, mycotoxigenic fungi and climate change related to wine. Food Res. Int. 2017, in press. [Google Scholar] [CrossRef]
- Gambuti, A.; Strollo, D.; Genovese, A.; Ugliano, M.; Ritieni, A.; Moio, L. Influence of enological practices on ochratoxin A concentration in wine. Am. J. Enol. Vitic. 2005, 56, 155–162. [Google Scholar]
- Food and Agriculture Organization (FAO). Manual on the Application of the HACCP System in Mycotoxin Prevention and Control; FAO Food and Nutrition Paper; Food and Agriculture Organization: Roma, Italy, 2001; Volume 73, pp. 1–124. Available online: http://www.fao.org/3/a-y1390e.pdf (accessed on 25 October 2017).
- International Organization of Vine and Wine (OIV). Resolution VITI/OENO 1/2005. Code of Sound Vitivinicultural Practices in Order to Minimise Levels of Ochratoxin A in Vine-Based Products. 2005. Available online: http://www.oiv.int/public/medias/2771/viti-oeno-1-2005-en.pdf (accessed on 25 October 2017).
- Chiodini, A.M.; Scherpenisse, P.; Bergwerff, A.A. Ochratoxin A contents in wine: Comparison of organically and conventionally produced products. J. Agric. Food Chem. 2006, 54, 7399–7404. [Google Scholar] [CrossRef] [PubMed]
- Lasram, S.; Mani, A.; Zaied, C.; Chebil, S.; Abid, S.; Bacha, H.; Mliki, A.; Ghorbel, A. Evolution of ochratoxin A content during red and rose vinification. J. Sci. Food Agric. 2008, 88, 1696–1703. [Google Scholar] [CrossRef]
- Bellver, J.; Fernández-Franzón, M.; Ruiz, M.J.; Juan-García, A. Presence of ochratoxin A (OTA) mycotoxin in alcoholic drinks from Southern European countries: Wine and beer. J. Agric. Food Chem. 2014, 62, 7643–7651. [Google Scholar] [CrossRef] [PubMed]
- Battilani, P.; Pietri, A. Ochratoxin A in grapes and wine. Eur. J. Plant Pathol. 2002, 108, 639–643. [Google Scholar] [CrossRef]
- Battilani, P.; Magan, N.; Logrieco, A. European research on ochratoxin A in grapes and wine. Int. J. Food Microbiol. 2006, 11, S2–S4. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Varelis, P.; Giannikopoulos, G.; Scott, E.S. Fate of ochratoxin A during vinification of Semillon and Shiraz grapes. J. Agric. Food Chem. 2006, 54, 6460–6464. [Google Scholar] [CrossRef] [PubMed]
- Dachery, B.; Fonseca, F.; Dal Magro, L.; Manfroi, V.; Welke, J.E. Exposure risk assessment to ochratoxin A through consumption of juice and wine considering the effect of steam extraction time and vinification stages. Food Chem. Toxicol. 2017, 109, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Csutorás, C.; Rácz, L.; Rácz, K.; Futo, P.; Forgó, P.; Kiss, A. Monitoring ochratoxin A during fermentation of different wines by applying high toxin concentrations. Microchem. J. 2013, 107, 182–184. [Google Scholar] [CrossRef]
- Fernandes, A.; Ratola, N.; Cerdeira, A.; Alves, A.; Venancio, A. Changes in ochratoxin A concentration during winemaking. Am. J. Enol. Vitic. 2007, 58, 92–96. [Google Scholar]
- Esti, M.; Benucci, I.; Liburdi, K.; Acciaro, G. Monitoring of ochratoxin A fate during alcoholic fermentation of wine-must. Food Control 2012, 27, 53–56. [Google Scholar] [CrossRef]
- Caridi, A.; Galvano, F.; Tafuri, A.; Ritieni, A. Ochratoxin A removal during winemaking. Enzyme Microb. Technol. 2006, 40, 122–126. [Google Scholar] [CrossRef]
- Cecchini, F.; Morassut, M.; García-Moruno, E.; Di Stefano, R. Influence of yeast strain on ochratoxin A content during fermentation of white and red must. Food Microbiol. 2006, 23, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Meca, G.; Blaiotta, G.; Ritieni, A. Reduction of ochratoxin A during the fermentation of Italian red wine Moscato. Food Control 2010, 21, 579–583. [Google Scholar] [CrossRef]
- Del Prete, V.; Rodríguez, H.; Carrascosa, A.V.; De las Rivas, B.; García-Moruno, E.; Muñoz, R. In vitro removal of ochratoxin A by wine lactic acid bacteria. J. Food Prot. 2007, 70, 2155–2160. [Google Scholar] [CrossRef] [PubMed]
- Mateo, E.M.; Medina, A.; Mateo, F.; Valle-Algarra, F.M.; Pardo, I.; Jiménez, M. Ochratoxin A removal in synthetic media by living and heat-inactivated cells of Oenococcus oeni isolated from wines. Food Control 2010, 21, 23–28. [Google Scholar] [CrossRef]
- Abrunhosa, L.; Ines, A.; Rodrigues, A.I.; Guimaraes, A.; Pereira, V.L.; Parpot, P.; Mendes-Faia, A.; Venancio, A. Biodegradation of ochratoxin A by Pediococcus parvulus isolated from Douro wines. Int. J. Food Microbiol. 2014, 188, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.L.; Hocking, A.D.; Scott, E.S. The effect of juice clarification, static or rotary fermentation and fining on ochratoxin A in wine. Aust. J. Grape Wine Res. 2006, 12, 245–251. [Google Scholar] [CrossRef]
- García-Moruno, E.; Sanlorenzo, C.; Boccaccino, B.; Di Stefano, R. Treatment with yeast to reduce the concentration of ochratoxin A in red wine. Am. J. Enol. Vitic. 2005, 56, 73–76. [Google Scholar]
- Castellari, M.; Versari, A.; Fabiani, A.; Parpinello, G.P.; Galassi, S. Removal of ochratoxin A in red wines by means of adsorption treatments with commercial fining agents. J. Agric. Food Chem. 2001, 49, 3917–3921. [Google Scholar] [CrossRef] [PubMed]
- Var, I.; Kabak, B.; Erginkaya, Z. Reduction in ochratoxin A levels in white wine, following treatment with activated carbon and sodium bentonite. Food Control 2008, 19, 592–598. [Google Scholar] [CrossRef]
- Anli, R.E.; Vural, N.; Bayram, M. Removal of ochratoxin A (OTA) from naturally contaminated wines during the vinification process. J. Inst. Brew. 2011, 117, 456–461. [Google Scholar] [CrossRef]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal from red wine by several oenological fining agents: Bentonite, egg albumin, allergen-free adsorbents, chitin and chitosan. Food Addit. Contam. A 2012, 29, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Niu, Y.; Ma, T.; Xu, P.; Huang, W.; Zhan, J. Determination, content analysis and removal efficiency of fining agents on ochratoxin A in Chinese wines. Food Control 2017, 73, 382–392. [Google Scholar] [CrossRef]
- Bornet, A.; Teissedre, P.L. Chitosan, chitin-glucan and chitin effects on minerals (iron, lead, cadmium) and organic (ochratoxin A) contaminants in wines. Eur. Food Res. Technol. 2008, 226, 681–689. [Google Scholar] [CrossRef]
- Kurtbay, H.M.; Bekçi, Z.; Merdivan, M.; Yurdakoç, K. Reduction of ochratoxin A levels in red wine by bentonite, modified bentonites and chitosan. J. Agric. Food Chem. 2008, 56, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Quintela, S.; Villarán, M.C.; López de Armentia, I.; Elejalde, E. Ochratoxin A removal in wine: A review. Food Control 2013, 30, 439–445. [Google Scholar] [CrossRef]
- Varga, J.; Kocsubé, S.; Péteri, Z.; Vágvölgyi, C.; Tóth, B. Chemical, physical and biological approaches to prevent ochratoxin induced toxicoses in humans and animals. Toxins 2010, 2, 1718–1750. [Google Scholar] [CrossRef] [PubMed]
- European Council. Council Regulation EC No 1493/1999 on the common organization of the market in wine. Off. J. Eur. Communities 1999, 179, 1–84. [Google Scholar]
- International Organization of Vine and Wine (OIV). Resolution OIV/OENO 338A/2009. Wines-Treatment Using Chitosan. 2009. Available online: http://www.oiv.int/public/medias/3551/f-code-ii-3416.pdf (accessed on 25 October 2017).
- International Organization of Vine and Wine (OIV). Resolution OIV/OENO 338B/2009. Wines-Treatment Using Chitin Glucane. 2009. Available online: http://www.oiv.int/public/medias/3553/f-code-ii-3417.pdf (accessed on 25 October 2017).
- The Joint FAO/WHO Expert Committee on Food Additives (JECFA). Safety Evaluation of Certain Mycotoxins in Food. WHO Food Additives Series 2001, No. 47. Available online: http://www.fao.org/3/a-bc528e.pdf (accessed on 25 October 2017).
- European Commission. Commission Regulation EC No 401/2006 laying down the methods of sampling and analysis for the official control of the levels of mycotoxins in foodstuffs. Off. J. Eur. Union 2006, 70, 12–70. [Google Scholar]
- European Food Safety Authority (EFSA). Opinion of the Scientific Panel on Contaminants in the Food Chain on a request from the Commission related to ochratoxin A in food. EFSA J. 2006, 365, 1–56. [Google Scholar]
- VICAM. Global Mycotoxin Regulations. Available online: https://play.google.com/store/apps/details?id=com.vicam.Regulations (accessed on 25 October 2017).
- National Health and Family Planning Commission (NHFPC). Maximum Levels of Mycotoxins in Food (GB 2761-2017). 2017. Available online: http://www.nhfpc.gov.cn/sps/s7891/201704/b83ad058ff544ee39dea811264878981.shtml (accessed on 25 October 2017).
- Bellí, N.; Marín, S.; Sanchis, V.; Ramos, A.J. Impact of fungicides on Aspergillus carbonarius growth and ochratoxin A production on synthetic grape-like medium and on grapes. Food Addit. Contam. 2006, 23, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Mateo, R.; Valle-Algarra, F.; Mateo, E.M.; Jiménez, M. Effect of carbendazim and physicochemical factors on the growth and ochratoxin A production of Aspergillus carbonarius isolated from grapes. Int. J. Food Microbiol. 2007, 119, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Tjamos, S.E.; Antoniou, P.P.; Kazantzidou, A.; Antonopoulos, D.F.; Papageorgiou, I.; Tjamos, E.C. Aspergillus niger and Aspergillus carbonarius in Corinth raisin and wine-producing vineyards in Greece: Population composition, ochratoxin A production and chemical control. J. Phytopathol. 2004, 152, 250–255. [Google Scholar] [CrossRef]
- Bellí, N.; Marín, S.; Argilés, E.; Ramos, A.J.; Sanchis, V. Effect of chemical treatments on ochratoxigenic fungi and common mycobiota of grapes (Vitis vinifera). J. Food Prot. 2007, 70, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Medina, A.; Jiménez, M.; Mateo, R.; Magan, N. Efficacy of natamycin for control of growth and ochratoxin A production by Aspergillus carbonarius strains under different environmental conditions. J. Appl. Microbiol. 2007, 103, 2234–2239. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Wang, Y.; Zeng, H.; Li, Z.; Zhang, P.; Tessema, A.; Peng, X. Efficacy and possible mechanisms of perillaldehyde in control of Aspergillus niger causing grape decay. Int. J. Food Microbiol. 2015, 202, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Favilla, M.; Pascale, M.; Ricelli, A.; Evidente, A.; Amalfitano, C.; Altomare, C. Inhibition of species of the Aspergillus section Nigri and ochratoxin production in grapes by fusapyrone. Appl. Environ. Microbiol. 2008, 74, 2248–2253. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Apaliya, M.T.; Mahunu, G.K.; Chen, L.; Li, W. Control of ochratoxin A-producing fungi in grape berry by microbial antagonists: A review. Trends Food Sci. Technol. 2016, 51, 88–97. [Google Scholar] [CrossRef]
- Bleve, G.; Grieco, F.; Cozzi, G.; Logrieco, A.; Visconti, A. Isolation of epiphytic yeast with potential biocontrol of Aspergillus carbonarius and A. niger on grape. Int. J. Food Microbiol. 2006, 108, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Ponsone, M.L.; Chiotta, M.L.; Combina, M.; Dalcero, A.; Chulze, S. Biocontrol as a strategy to reduce the impact of ochratoxin A and Aspergillus section Nigri in grapes. Int. J. Food Microbiol. 2011, 151, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Fiori, S.; Urgeghe, P.P.; Hammami, W.; Razzu, S.; Jaoua, S.; Migueli, Q. Biocontrol activity of four non- and low-fermenting yeast strains against Aspergillus carbonarius and their ability to remove ochratoxin A from grape juice. Int. J. Food Microbiol. 2014, 189, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Pantelides, I.S.; Christou, O.; Tsolakidou, D.; Tsaltas, D.; Ioannou, N. Isolation, identification and in vitro screening of grapevine yeast for the control of black Aspergilli on grapes. Biol. Control 2015, 88, 46–53. [Google Scholar] [CrossRef]
- Dimakopoulou, M.; Tjamos, S.E.; Antoniou, P.P.; Pietri, A.; Battilani, P.; Avramidis, N.; Markakis, E.A.; Tjamos, E.C. Phyllosphere grapevine yeast Aureobasidium pullulans reduce Aspergillus carbonarius (sour rot) incidence in wine-producing vineyards in Greece. Biol. Control 2008, 46, 158–165. [Google Scholar] [CrossRef]
- Ponsone, M.L.; Nally, M.C.; Chiotta, M.L.; Combina, M.; Köhl, J.; Chulze, S.N. Evaluation of the effectiveness of potential biocontrol yeasts against black sour rot and ochratoxin A occurring under greenhouse and field grape production conditions. Biol. Control 2016, 103, 78–85. [Google Scholar] [CrossRef]
- De Curtis, F.; de Felice, D.V.; Ianiri, G.; De Cicco, V.; Castoria, R. Environmental factors affect the activity of biocontrol agents against ochratoxigenic Aspergillus carbonarius on wine grape. Int. J. Food Microbiol. 2012, 159, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Solfrizzo, M.; Avantaggiato, G.; Panzarini, G.; Visconti, A. Removal of ochratoxin A from contaminated red wines by repassage over grape pomaces. J. Agric. Food Chem. 2010, 58, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Farbo, M.G.; Urgeghe, P.P.; Fiori, S.; Marceddu, S.; Jaoua, S.; Migueli, Q. Adsorption of ochratoxin A from grape juice by yeast cells immobilised in calcium alginate beads. Int. J. Food Microbiol. 2015, 217, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Bejaoui, H.; Mathieu, F.; Taillandier, P.; Lebrihi, A. Ochratoxin A removal in synthetic and natural grape juices by selected oenological Saccharomyces strains. J. Appl. Microbiol. 2004, 97, 1038–1044. [Google Scholar] [CrossRef] [PubMed]
- Petruzzi, L.; Baiano, A.; De Gianni, A.; Sinigaglia, M.; Corbo, M.R.; Bevilacqua, A. Differential adsorption of ochratoxin A and anthocyanins by inactivated yeasts and yeast cell walls during simulation of wine aging. Toxins 2015, 7, 4350–4365. [Google Scholar] [CrossRef] [PubMed]
- European Committee for Standardization. CSN EN 14133—Foodstuffs—Determination of Ochratoxin A in Wine and Beer—HPLC with Immunoaffinity Column Clean-Up; Czech Office for Standards, Metrology and Testing: Praha, Czech Republic, 2009. [Google Scholar]
- Brera, C.; Grossi, S.; De Santis, B.; Miraglia, M. Automated HPLC method for the determination of ochratoxin A in wine samples. J. Liq. Chromatogr. Relat. Technol. 2003, 26, 119–133. [Google Scholar] [CrossRef]
- Giraudi, G.; Ferrero, V.E.V.; Anfossi, L.; Baggiani, C.; Giovannoli, C.; Tozzi, C. Solid-phase extraction of ochratoxin A from wine based on a binding hexapeptide prepared by combinatorial synthesis. J. Chromatogr. 2007, 1175, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Kong, W.; Zhou, S.; Yin, L.; Wan, L.; Yang, M. Molecularly imprinted polymer-based solid phase clean-up for analysis of ochratoxin A in beer, red wine, and grape juice. J. Sep. Sci. 2013, 36, 1291–1297. [Google Scholar] [CrossRef] [PubMed]
- Hernández, M.J.; García-Moreno, M.V.; Durán, E.; Guillén, D.; Barroso, G. Validation of two analytical methods for the determination of ochratoxin A by reversed-phased high-performance liquid chromatography coupled to fluorescence detection in must and sweet wines from Andalusia. Anal. Chim. Acta 2006, 566, 117–121. [Google Scholar] [CrossRef]
- Longobardi, F.; Iacovelli, V.; Catucci, L.; Panzarini, G.; Pascale, M.; Visconti, A.; Agostiano, A. Determination of ochratoxin A in wine by means of immunoaffinity and aminopropyl solid-phase column cleanup and fluorometric detection. J. Agric. Food Chem. 2013, 61, 1604–1608. [Google Scholar] [CrossRef] [PubMed]
- Dall’Asta, C.; Galaverna, G.; Dossena, A.; Marchelli, R. Reversed-phase liquid chromatographic method for the determination of ochratoxin A in wine. J. Chromatogr. A 2004, 1024, 275–279. [Google Scholar] [CrossRef] [PubMed]
- García-Fonseca, S.; Ballesteros-Gómez, A.; Rubio, S.; Pérez-Bendito, D. Coacervate extraction of ochratoxin A in wines prior to liquid chromatography/fluorescence determination. Anal. Chim. Acta 2008, 617, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.C.; Bonel, L.; Ezquerra, A.; Duato, P.; Castillo, J.R. An electrochemical immunosensor for ochratoxin A determination in wines based on a monoclonal antibody and paramagnetic microbeads. Anal. Bioanal. Chem. 2012, 403, 1585–1593. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.J.; Barros, N.; Camara, J.S. A survey of the occurrence of ochratoxin A in Madeira wines based on a modified QuEChERS extraction procedure combined with liquid chromatography-triple quadrupole tandem mass spectrometry. Food Res. Int. 2013, 54, 293–301. [Google Scholar] [CrossRef]
- Zezza, F.; Longobardi, F.; Pascale, M.; Eremin, S.A.; Visconti, A. Fluorescence polarization immunoassay for rapid screening of ochratoxin A in red wine. Anal. Bioanal. Chem. 2009, 395, 1317–1323. [Google Scholar] [CrossRef] [PubMed]
- Arroyo-Manzanares, N.; Gámiz-Gracia, L.; García-Campaña, A.M. Determination of ochratoxin A in wines by capillary liquid chromatography with laser induced fluorescence detection using dispersive liquid-liquid microextraction. Food Chem. 2012, 135, 368–372. [Google Scholar] [CrossRef] [PubMed]
- Barthelmebs, L.; Jonca, J.; Hayat, A.; Prieto-Simon, B.; Marty, J.L. Enzyme-Linked Aptamer Assays (ELAAs), based on a competition format for a rapid and sensitive detection of ochratoxin A in wine. Food Control 2011, 22, 737–743. [Google Scholar] [CrossRef]
- Yin, X.; Wang, S.; Liu, X.; He, C.; Tang, Y.; Li, Q.; Liu, Q.; Su, H.; Tan, T.; Dong, Y. Aptamer-based colorimetric biosensing of ochratoxin A in fortified white grape wine sample using unmodified gold nanoparticles. Anal. Sci. 2017, 33, 659–664. [Google Scholar] [CrossRef] [PubMed]
- Remiro, R.; González-Peñas, E.; Lizarraga, E.; López de Cerain, A. Quantification of ochratoxin A and five analogs in Navarra red wines. Food Control 2012, 27, 139–145. [Google Scholar] [CrossRef]
- Di Stefano, V.; Avellone, G.; Pitonzo, R.; Capocchiano, V.G.; Mazza, A.; Cicero, N.; Dugo, G. Natural co-occurrence of ochratoxin A, ochratoxin B and aflatoxins in Sicilian red wines. Food Addit. Contam. Part A 2015, 32, 1343–1351. [Google Scholar] [CrossRef] [PubMed]



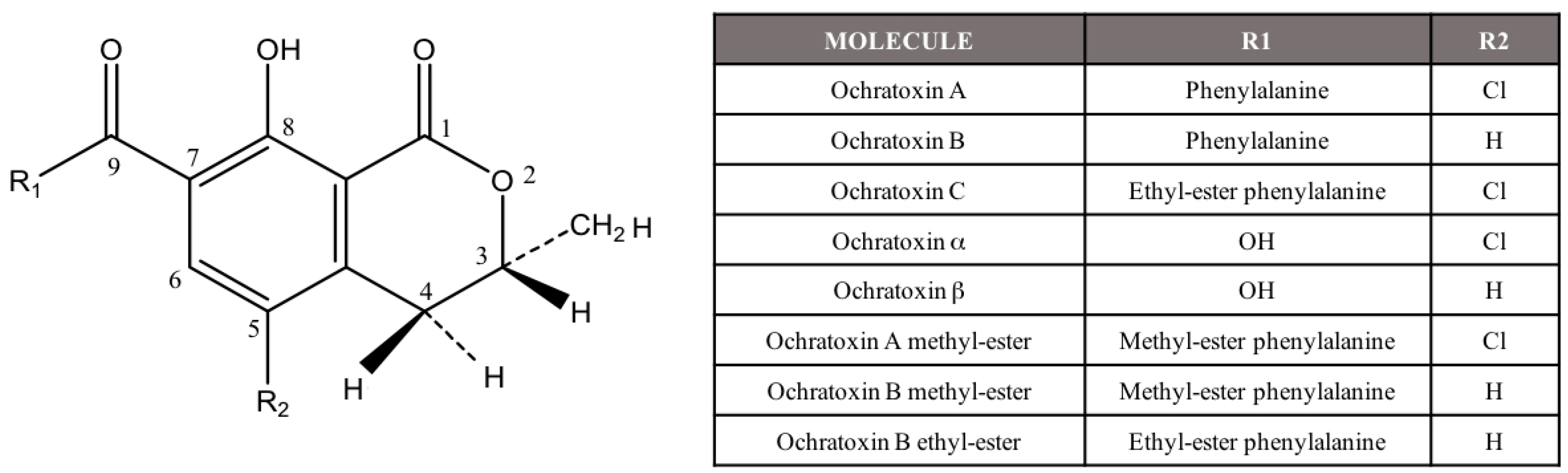{kind=link}
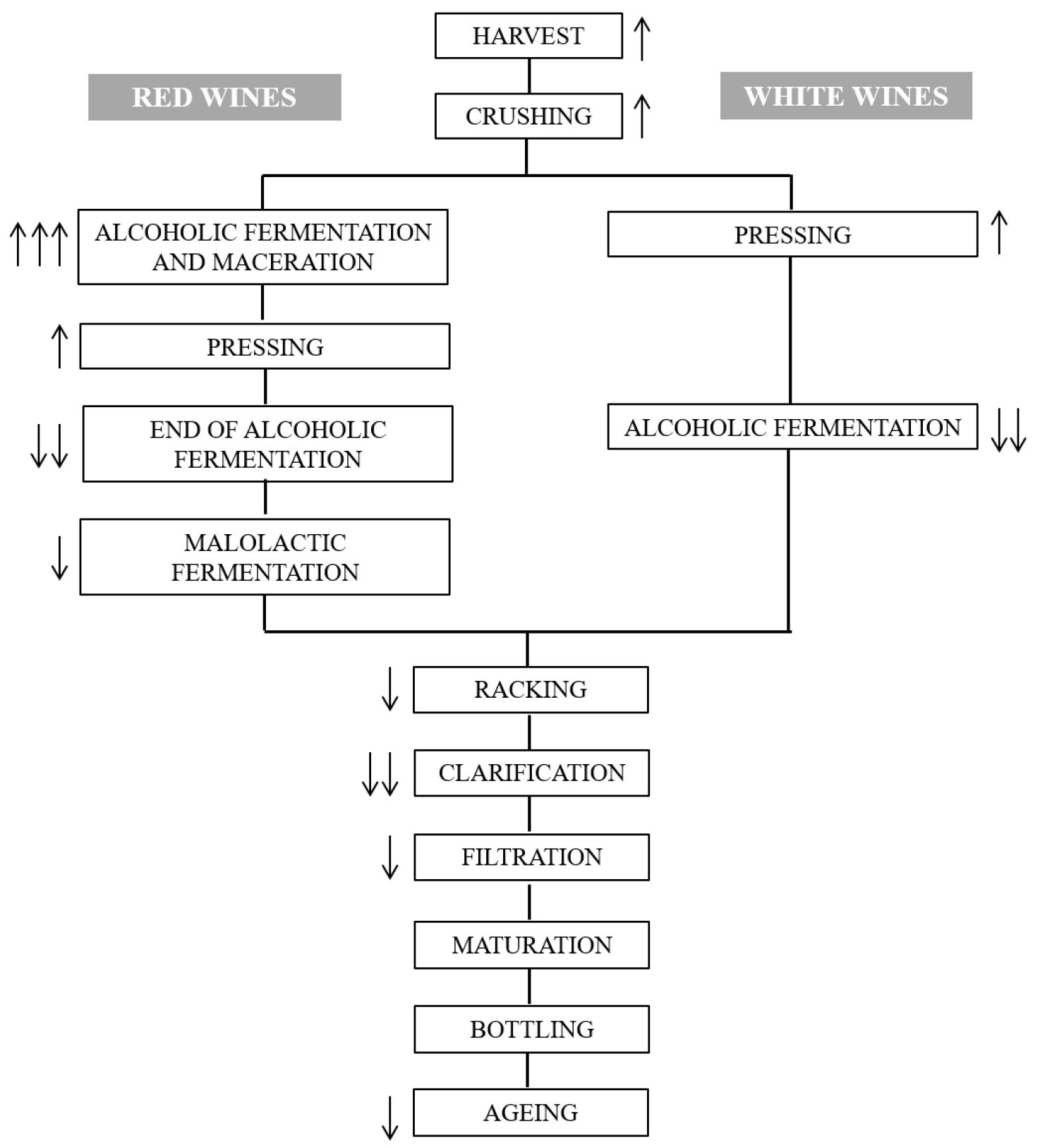{kind=link}
| COUNTRY | RED | WHITE | DESSERT | ANALYSIS METHOD | MAXIMUM OTA (µg/L) | SAMPLES ABOVE LIMITS | REFERENCE |
|---|---|---|---|---|---|---|---|
| Argentina | 4/47 (9%) | - | - | HPLC | 4.82 | 2 (4%) | [42] |
| Australia | 49/344 (14%) | 41/257 (16%) | 3/13 (23%) | HPLC | 0.62 | 0 (0%) | [43] |
| Canada | 10/43 (23%) | 5/36 (14%) | - | LC | 0.39 | 0 (0%) | [44] |
| Chile | 28/841 (3%) | 6/313 (2%) | - | HPLC | 0.35 | 0 (0%) | [45] |
| China | 18/63 (29%) | 4/42 (10%) | - | LC/MS | 1.18 | 0 (0%) | [46] |
| 44/77 (57%) | 1/34 (3%) | - | HPLC | 5.65 | 3 (3%) | [47] | |
| Czech Republic | 2/13 (15%) | 3/33 (9%) | - | UPLC | 0.07 | 0 (0%) | [48] |
| France | - | - | 47/49 (96%) | LC/MS | 1.22 | 0 (0%) | [49] |
| Greece | 71/104 (68%) | 63/118 (53%) | 15/18 (83%) | HPLC | 2.82 | n.a. | [50] |
| 9/14 (65%) | 7/13 (54%) | 6/7 (86%) | HPLC | 3.20 | 3 (9%) | [51] | |
| 45/64 (70%) | 31/49 (63%) | 22/27 (81%) | HPLC | 2.00 | 0 (0%) | [52] | |
| Hungary | 33/33 (100%) | 31/32 (97%) | - | HPLC | 0.53 | 0 (0%) | [53] |
| Italy | 86/92 (93%) | - | 9/15 (60%) | HPLC | 3.86 | n.a. | [54] |
| 695/1002 (69%) | 125/204 (61%) | - | HPLC | 2.63 | 29 (2%) | [55] | |
| 37/38 (97%) | 4/9 (57%) | - | HPLC | 7.63 | 6 (13%) | [56] | |
| 535/773 (69%) | 128/290 (44%) | 18/28 (64%) | HPLC | 7.50 | 22 (2%) | [57] | |
| - | - | 29/30 (97%) | HPLC | 1.56 | 0 (0%) | [58] | |
| Romania | 17/44 (39%) | 21/55 (38%) | - | HPLC | 1.89 | 0 (0%) | [59] |
| Spain | 24/130 (19%) | 4/50 (8%) | - | HPLC | 4.24 | 2 (1%) | [60] |
| 108/188 (57%) | 4/6 (67%) | - | HPLC | 0.18 | 0 (0%) | [61] | |
| - | - | 32/40 (80%) | LC | 0.64 | 0 (0%) | [62] | |
| - | - | 186/188 (99%) | LC/MS | 4.63 | 18 (1%) | [49] | |
| South Africa | 9/9 (100%) | 15/15 (100%) | - | HPLC | 0.39 | 0 (0%) | [63] |
| 19/65 (29%) | 20/42 (48%) | 11/15 (73%) | HPLC | 2.67 | n.a. | [64] | |
| Turkey | 44/51 (86%) | 29/34 (85%) | - | HPLC | 0.82 | 0 (0%) | [65] |
| Type of Wine | Extraction | Clean-up | Detection | Recovery | Reproducibility (%RSD) | Detection Limits | Special Remarks | Reference |
|---|---|---|---|---|---|---|---|---|
| White and red | PBS | IAC | HPLC-FD | 84–95% | <5% | 0.01 µg/L | Automated protocol | [136] |
| White, rosé, red, dessert | PEG8000 | Peptide SPE | HPLC | 75–113% | n.a | 0.10 µg/L | Efficiency similar to IAC | [137] |
| Red and must | - | MIP SPE | HPLC | 91–103% | <5% | 0.025 µg/L | MIP columns are reusable | [138] |
| White, rosé, red, dessert | PEG8000 + NaHCO3 | IAC | HPLC-MS | 88–104% | <10% | 0.01 µg/L | Reference method | [56] |
| Red and must | - | C18 SPE | HPLC-FD | 94–114% | <2% | 0.22–0.77 µg/L | Must do not require SPE | [139] |
| Red | PEG8000 + NaHCO3 | IAC + Aminopropyl SPE | HPLC-FD | 95–105% | <15% | 0.2 µg/L | Double clean up to reduce matrix interference | [140] |
| White, rosé, red, dessert | - | - | HPLC-FD | 93–96% | <5% | 0.05 µg/L | Without extraction and clean up | [141] |
| White, rosé, red | - | Coacervate of decanoic acid in THF | HPLC | 85–100% | <7% | 0.004–0.015 µg/L | - | [142] |
| Red | PVP + PEG3350 | Magnetic beads linked to anti-OTA antibodies | Electrochemical | 105–109% | <9% | 0.11 µg/L | Competitive electrochemical immunosensor | [143] |
| Dessert | QuEChERS | Dispersive SPE | HPLC-MS | 87–103% | <9% | 0.1 µg/L | - | [144] |
| Red | - | Aminopropyl SPE | FP immunoassay | 75–83% | <12% | 0.7 µg/L | Rapid procedure (less than 10 min) | [145] |
| White, rosé, red | - | DLLME | HPLC-LIF | 92–98% | <5% | 0.006 µg/L | Green analytical chemistry alternative | [146] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gil-Serna, J.; Vázquez, C.; González-Jaén, M.T.; Patiño, B. Wine Contamination with Ochratoxins: A Review. Beverages 2018, 4, 6. https://doi.org/10.3390/beverages4010006
Gil-Serna J, Vázquez C, González-Jaén MT, Patiño B. Wine Contamination with Ochratoxins: A Review. Beverages. 2018; 4(1):6. https://doi.org/10.3390/beverages4010006
Chicago/Turabian StyleGil-Serna, Jessica, Covadonga Vázquez, María Teresa González-Jaén, and Belén Patiño. 2018. "Wine Contamination with Ochratoxins: A Review" Beverages 4, no. 1: 6. https://doi.org/10.3390/beverages4010006
APA StyleGil-Serna, J., Vázquez, C., González-Jaén, M. T., & Patiño, B. (2018). Wine Contamination with Ochratoxins: A Review. Beverages, 4(1), 6. https://doi.org/10.3390/beverages4010006