
Establishment of an In Vitro Scab Model for Investigating Different Phases of Wound Healing

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Culture of HaCat Cells
2.3. Production and Culture of the In Vitro Scab Model
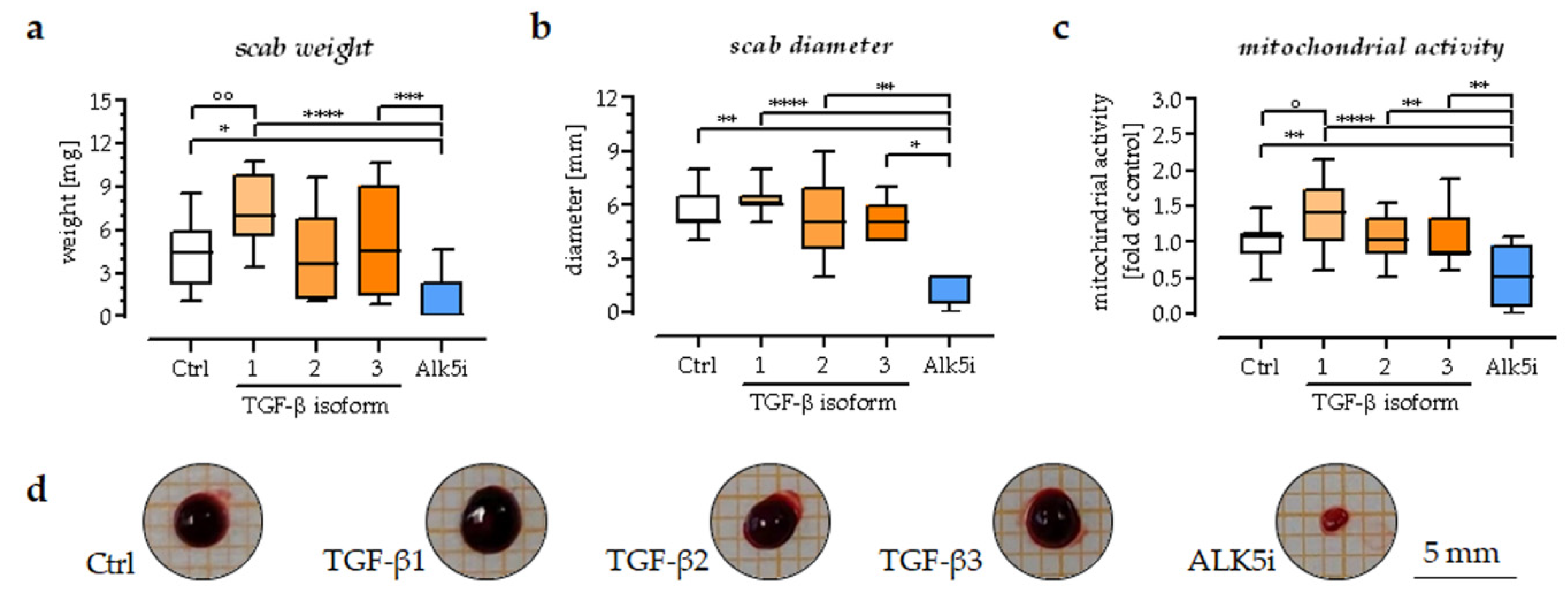
2.4. The Measurement of Size and Weight of the In Vitro Scab Model
2.5. Resazurin Conversion
2.6. Viability Staining
2.7. Cytokine Array
2.8. Gene Expression Analyses
2.9. Luciferase Reporter Assay
2.10. Statistical Analysis
3. Results
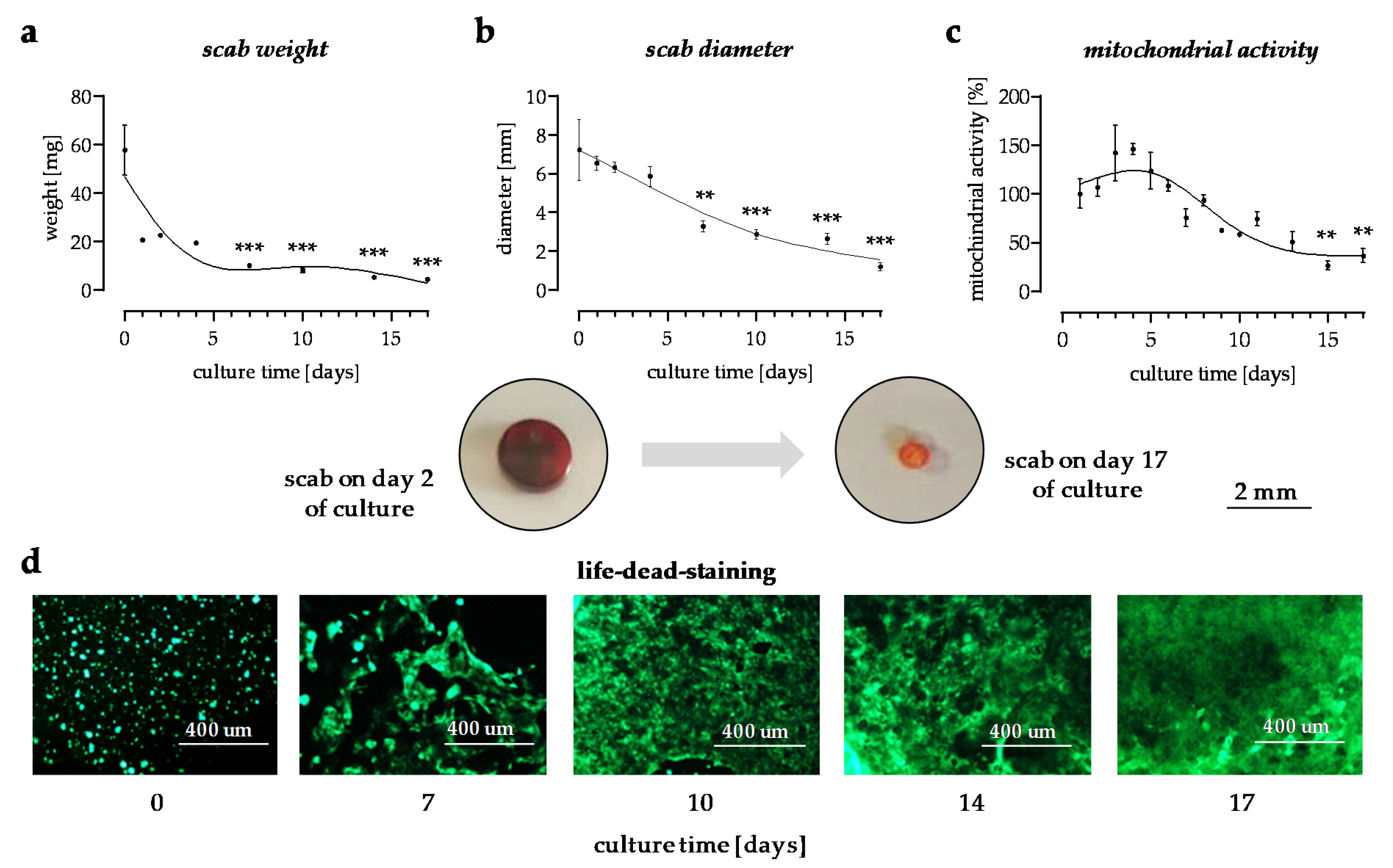
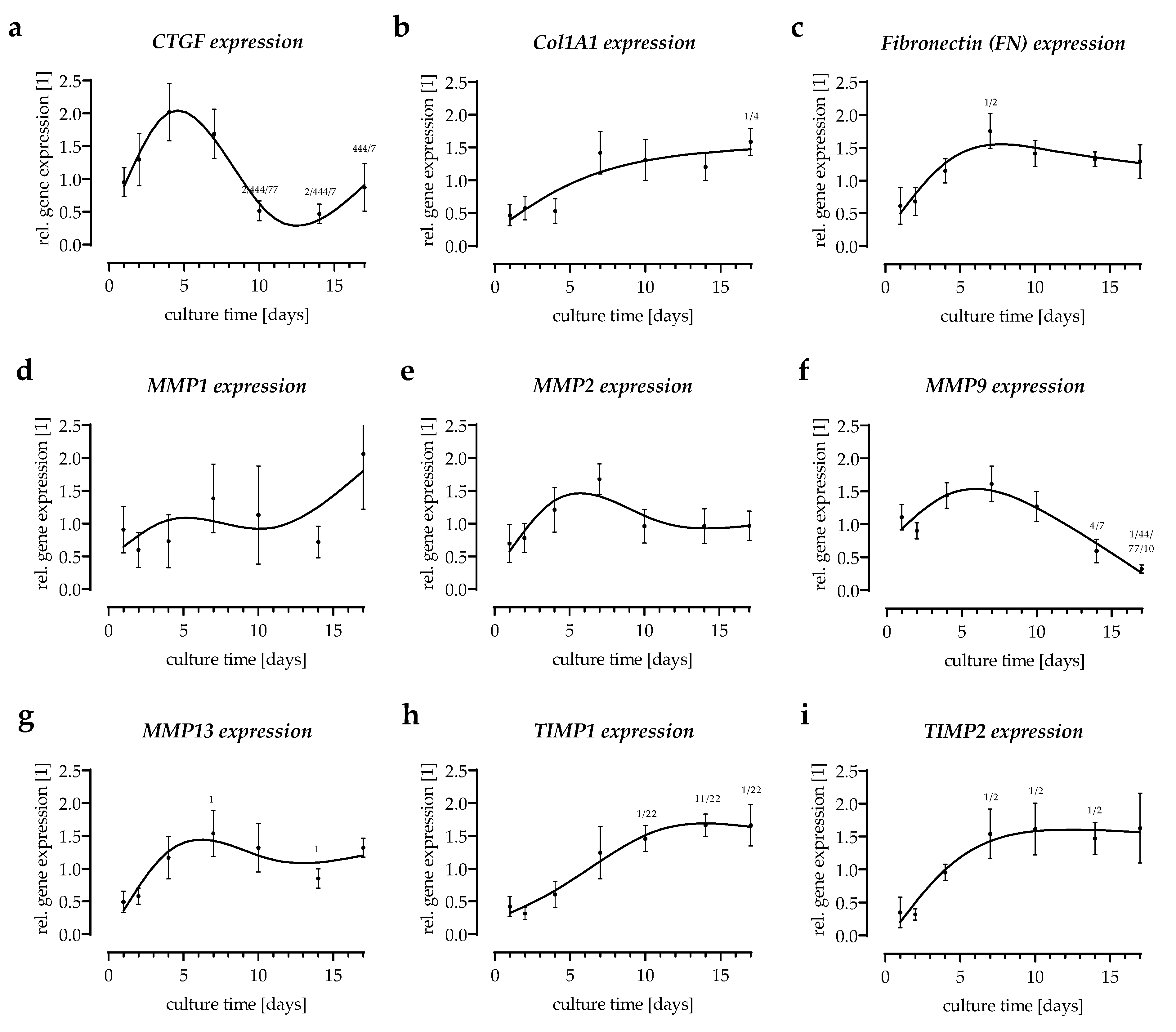
3.1. In Vitro Scabs Can Be Investigated for More than 14 Days in Culture
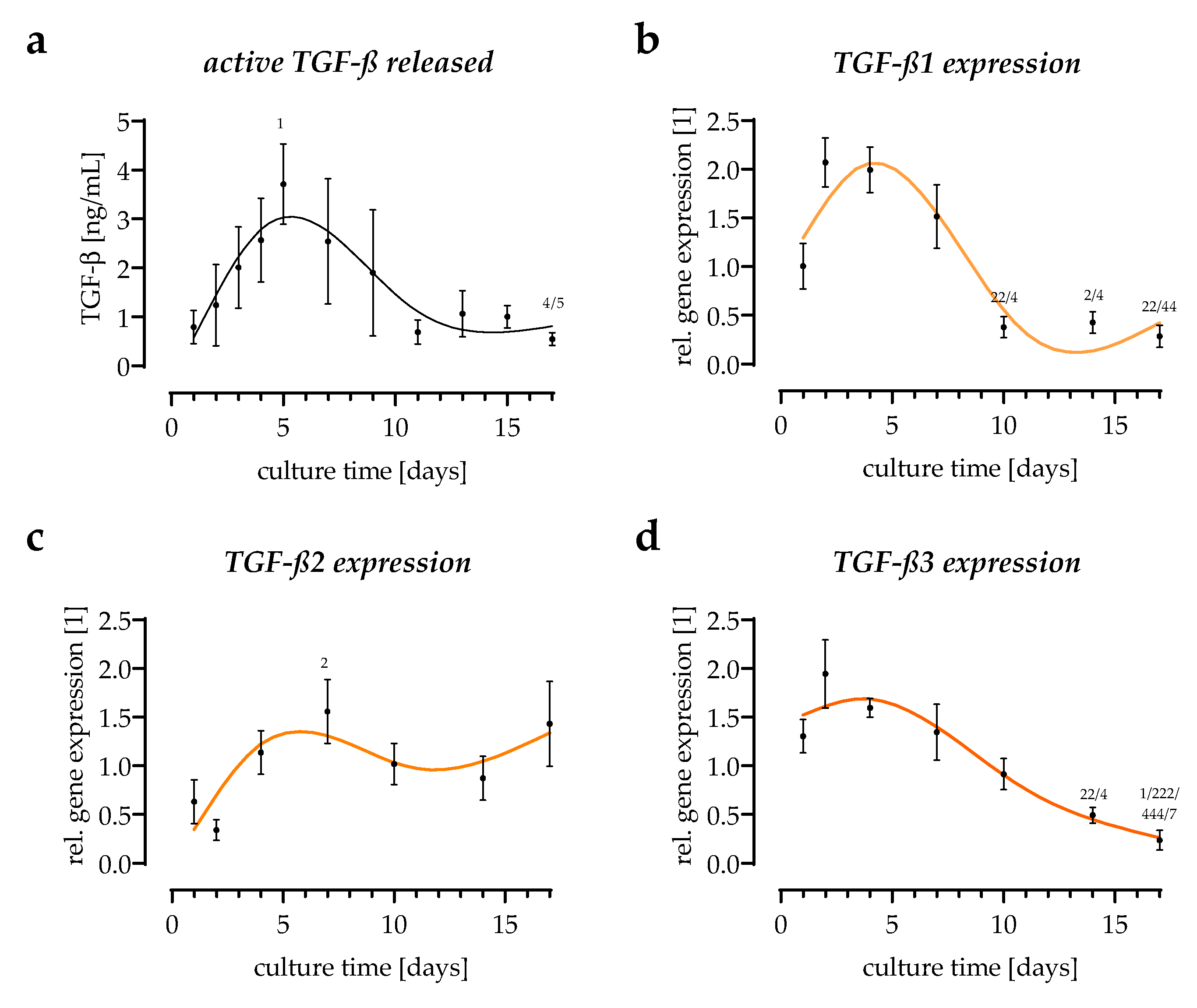
3.2. Cytokines Released from the In Vitro Scabs in the First Week of Culture
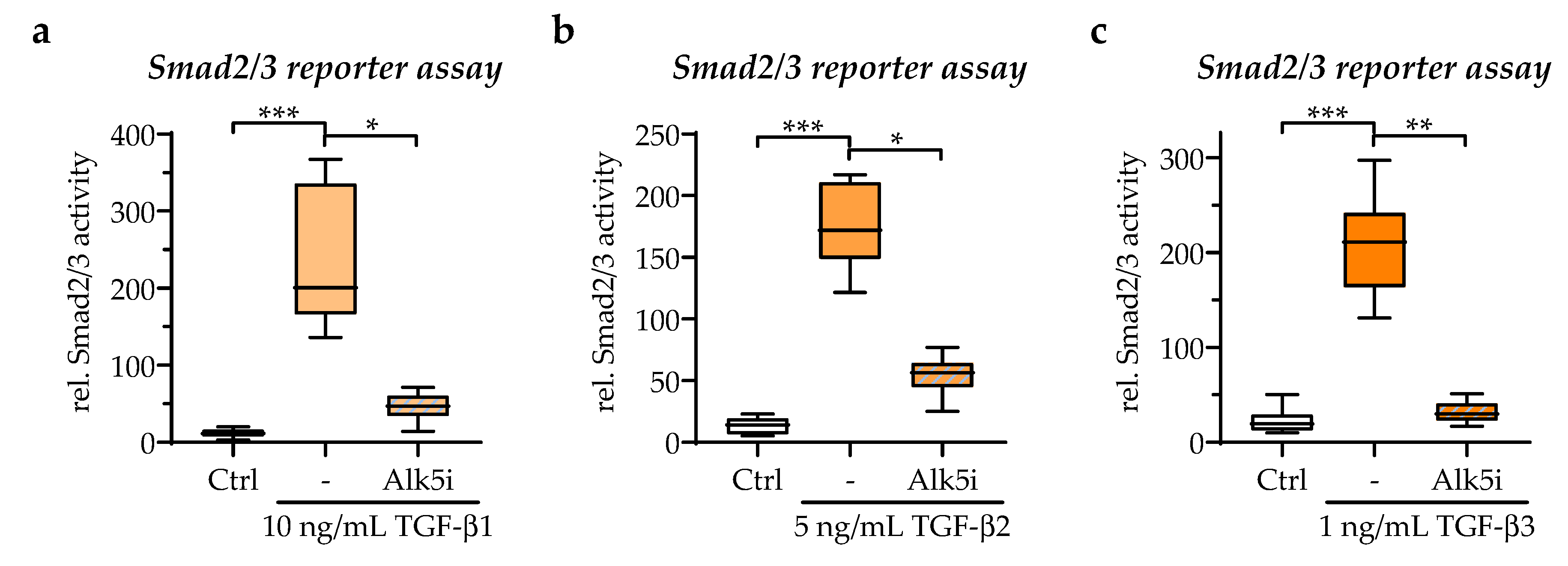
3.3. All Three TGF-β Isoforms Can Be Detected in the In Vitro Scab Model
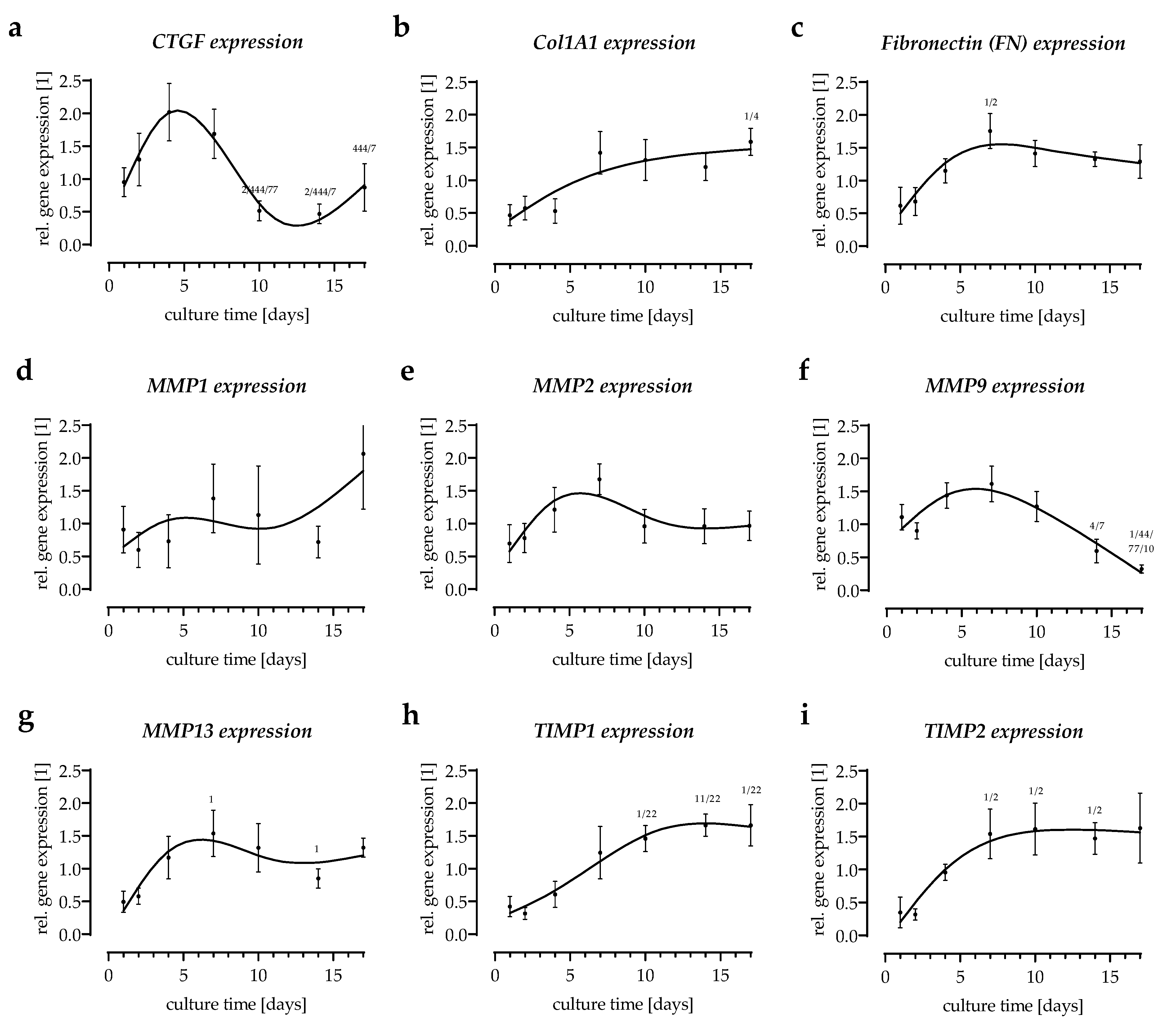
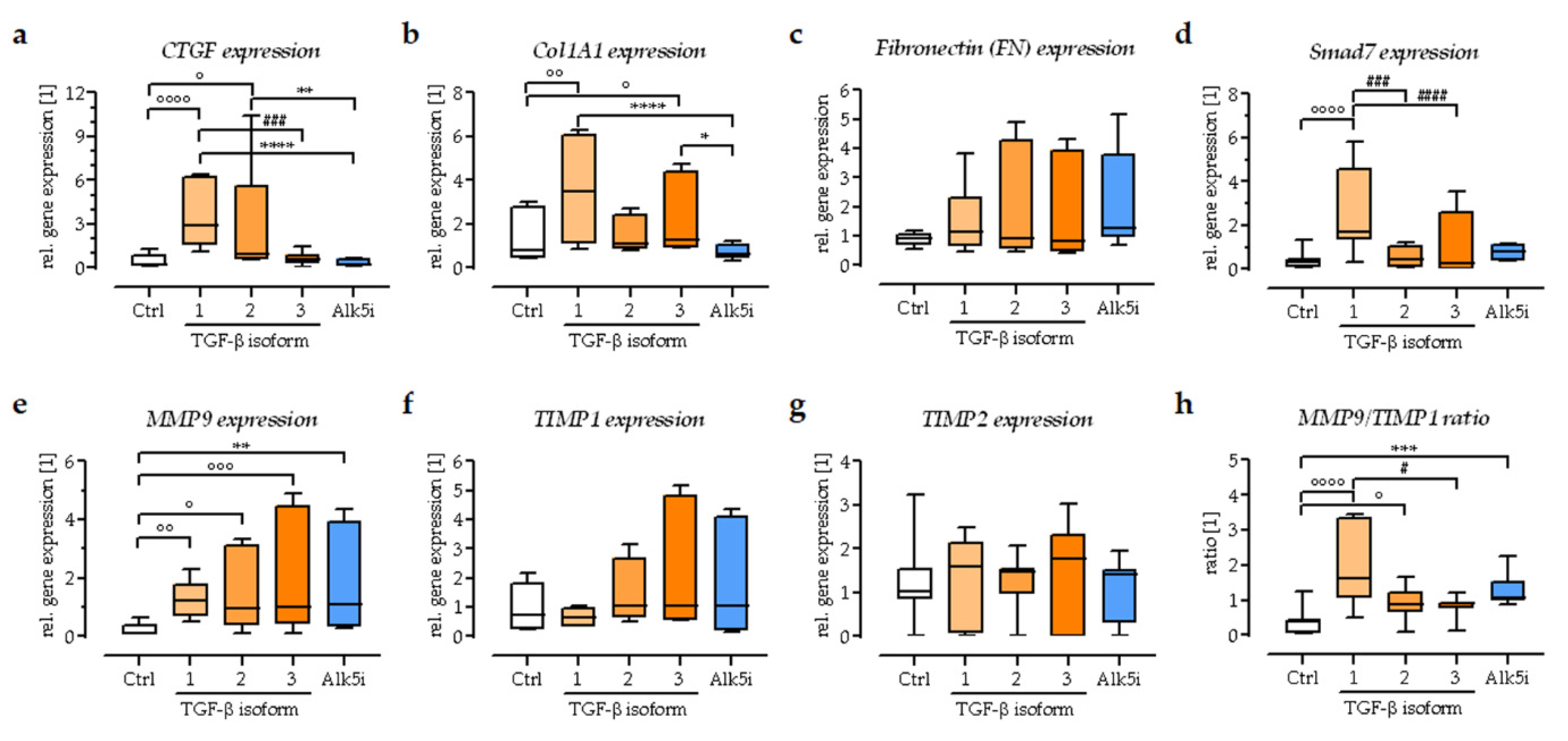
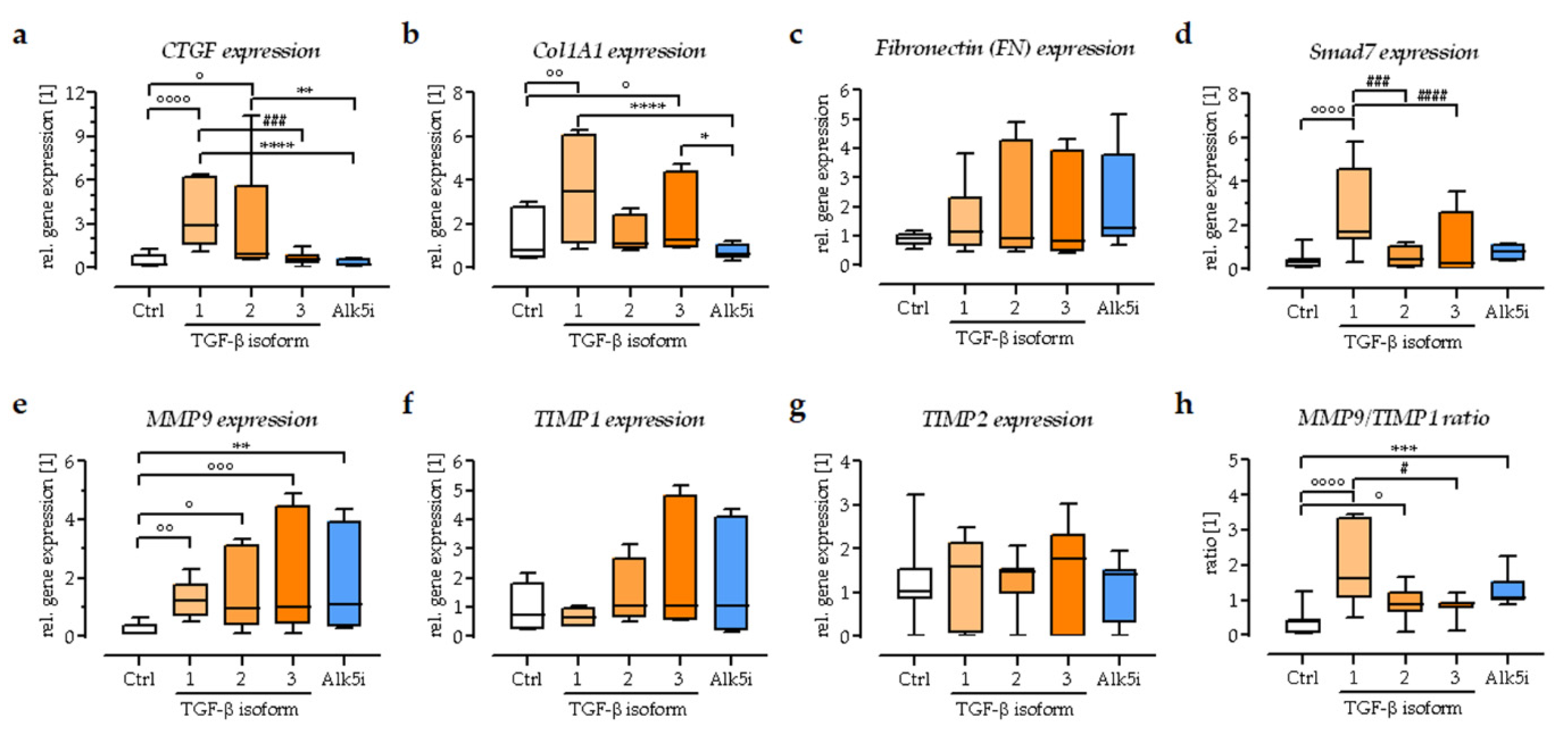
3.4. Expression of Extracellular Matrix-Related Genes in the In Vitro Scabs
3.5. TGF-β1 Could Promote the Area, Weight, and Mitochondrial Activity of the Scab Model
3.6. Exogenous Stimulation with TGF-β1 Increased the Expression of Smad7 in the In Vitro Scabs
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Karl, T.; Gussmann, A.; Storck, M. Chronic wounds—Perspective for integrated care. Zentralbl. Chir. 2007, 132, 232–235. [Google Scholar] [CrossRef]
- Gottrup, F. A specialized wound-healing center concept: Importance of a multidisciplinary department structure and surgical treatment facilities in the treatment of chronic wounds. Am. J. Surg. 2004, 187, 38S–43S. [Google Scholar] [CrossRef]
- Pfeiffenberger, M.; Bartsch, J.; Hoff, P.; Ponomarev, I.; Barnewitz, D.; Thone-Reineke, C.; Buttgereit, F.; Gaber, T.; Lang, A. Hypoxia and mesenchymal stromal cells as key drivers of initial fracture healing in an equine in vitro fracture hematoma model. PLoS ONE 2019, 14, e0214276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, A.B.; Sporn, M.B.; Assoian, R.K.; Smith, J.M.; Roche, N.S.; Wakefield, L.M.; Heine, U.I.; Liotta, L.A.; Falanga, V.; Kehrl, J.H.; et al. Transforming growth factor type beta: Rapid induction of fibrosis and angiogenesis in vivo and stimulation of collagen formation in vitro. Proc. Natl. Acad. Sci. USA 1986, 83, 4167–4171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nall, A.V.; Brownlee, R.E.; Colvin, C.P.; Schultz, G.; Fein, D.; Cassisi, N.J.; Nguyen, T.; Kalra, A. Transforming growth factor beta 1 improves wound healing and random flap survival in normal and irradiated rats. Arch. Otolaryngol. Head Neck Surg. 1996, 122, 171–177. [Google Scholar] [CrossRef]
- Pakyari, M.; Farrokhi, A.; Maharlooei, M.K.; Ghahary, A. Critical Role of Transforming Growth Factor Beta in Different Phases of Wound Healing. Adv. Wound Care 2013, 2, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lichtman, M.K.; Otero-Vinas, M.; Falanga, V. Transforming growth factor beta (TGF-beta) isoforms in wound healing and fibrosis. Wound Repair Regen. 2016, 24, 215–222. [Google Scholar] [CrossRef] [PubMed]
- Ehnert, S.; Linnemann, C.; Aspera-Werz, R.H.; Bykova, D.; Biermann, S.; Fecht, L.; De Zwart, P.M.; Nussler, A.K.; Stuby, F. Immune Cell Induced Migration of Osteoprogenitor Cells Is Mediated by TGF-beta Dependent Upregulation of NOX4 and Activation of Focal Adhesion Kinase. Int. J. Mol. Sci. 2018, 19, 2239. [Google Scholar] [CrossRef] [Green Version]
- Haussling, V.; Deninger, S.; Vidoni, L.; Rinderknecht, H.; Ruoss, M.; Arnscheidt, C.; Athanasopulu, K.; Kemkemer, R.; Nussler, A.K.; Ehnert, S. Impact of Four Protein Additives in Cryogels on Osteogenic Differentiation of Adipose-Derived Mesenchymal Stem Cells. Bioengineering 2019, 6, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehnert, S.; Baur, J.; Schmitt, A.; Neumaier, M.; Lucke, M.; Dooley, S.; Vester, H.; Wildemann, B.; Stockle, U.; Nussler, A.K. TGF-beta(1) As Possible Link between Loss of Bone Mineral Density and Chronic Inflammation. PLoS ONE 2010, 5, 67. [Google Scholar] [CrossRef] [PubMed]
- Landen, N.X.; Li, D.; Stahle, M. Transition from inflammation to proliferation: A critical step during wound healing. Cell Mol. Life Sci. 2016, 73, 3861–3885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Qiu, C.X.; Ludlow, A.; Ferguson, M.W.; Brunner, G. Active transforming growth factor-beta in wound repair: Determination using a new assay. Am. J. Pathol. 1999, 154, 105–111. [Google Scholar] [CrossRef]
- Roberts, A.B.; Mccune, B.K.; Sporn, M.B. Tgf-Beta—Regulation of Extracellular-Matrix. Kidney Int. 1992, 41, 557–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishida, T.; Kawaki, H.; Baxter, R.M.; DeYoung, R.A.; Takigawa, M.; Lyons, K.M. CCN2 (Connective Tissue Growth Factor) is essential for extracellular matrix production and integrin signaling in chondrocytes. J. Cell Commun. Signal. 2007, 1, 45–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Secker, G.A.; Shortt, A.J.; Sampson, E.; Schwarz, Q.P.; Schultz, G.S.; Daniels, J.T. TGF beta stimulated re-epithelialisation is regulated by CTGF and Ras/MEK/ERK signalling. Exp. Cell Res. 2008, 314, 131–142. [Google Scholar] [CrossRef]
- Frazier, K.; Williams, S.; Kothapalli, D.; Klapper, H.; Grotendorst, G.R. Stimulation of fibroblast cell growth, matrix production, and granulation tissue formation by connective tissue growth factor. J. Investig. Dermatol. 1996, 107, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Roberts, A.B.; Heine, U.I.; Flanders, K.C.; Sporn, M.B. Transforming Growth-Factor-Beta—Major Role in Regulation of Extracellular-Matrix. Ann. N. Y. Acad. Sci. 1990, 580, 225–232. [Google Scholar] [CrossRef]
- Ignotz, R.A.; Massague, J. Transforming Growth-Factor-Beta Stimulates the Expression of Fibronectin and Collagen and Their Incorporation into the Extracellular-Matrix. J. Biol. Chem. 1986, 261, 4337–4345. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, X.F.; Wang, Z.C.; Lou, D.; Fang, Q.Q.; Hu, Y.Y.; Zhao, W.Y.; Zhang, L.Y.; Wu, L.H.; Tan, W.Q. Current potential therapeutic strategies targeting the TGF-β/Smad signaling pathway to attenuate keloid and hypertrophic scar formation. Biomed. Pharmacother. 2020, 129, 110287. [Google Scholar] [CrossRef]
- Purohit, T.; Qin, Z.; Quan, C.; Lin, Z.; Quan, T. Smad3-dependent CCN2 mediates fibronectin expression in human skin dermal fibroblasts. PLoS ONE 2017, 12, e0173191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bielefeld, K.A.; Amini-Nik, S.; Whetstone, H.; Poon, R.; Youn, A.; Wang, J.; Alman, B.A. Fibronectin and beta-catenin act in a regulatory loop in dermal fibroblasts to modulate cutaneous healing. J. Biol. Chem. 2011, 286, 27687–27697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakao, A.; Afrakhte, M.; Moren, A.; Nakayama, T.; Christian, J.L.; Heuchel, R.; Itoh, S.; Kawabata, N.; Heldin, N.E.; Heldin, C.H.; et al. Identification of Smad7, a TGF beta-inducible antagonist of TGF-beta signalling. Nature 1997, 389, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Caley, M.P.; Martins, V.L.; O’Toole, E.A. Metalloproteinases and Wound Healing. Adv. Wound Care 2015, 4, 225–234. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.E.; Parks, W.C. Metalloproteinases and their inhibitors: Regulators of wound healing. Int. J. Biochem. Cell B 2008, 40, 1334–1347. [Google Scholar] [CrossRef] [Green Version]
- Youn, M.; Huang, H.; Chen, C.; Kam, S.; Wilkes, M.C.; Chae, H.D.; Sridhar, K.J.; Greenberg, P.L.; Glader, B.; Narla, A.; et al. MMP9 inhibition increases erythropoiesis in RPS14-deficient del(5q) MDS models through suppression of TGF-β pathways. Blood Adv. 2019, 3, 2751–2763. [Google Scholar] [CrossRef]
- Chen, G.B.; Ge, D.D.; Zhu, B.Z.; Shi, H.X.; Ma, Q.L. Upregulation of matrix metalloproteinase 9 (MMP9)/tissue inhibitor of metalloproteinase 1 (TIMP1) and MMP2/TIMP2 ratios may be involved in lipopolysaccharide-induced acute lung injury. J. Int. Med. Res. 2020, 48, 300060520919592. [Google Scholar] [CrossRef]
- Barton, A.K.; Shety, T.; Bondzio, A.; Einspanier, R.; Gehlen, H. Metalloproteinases and Their Tissue Inhibitors in Comparison between Different Chronic Pneumopathies in the Horse. Mediat. Inflamm. 2015, 2015, 569512. [Google Scholar] [CrossRef] [Green Version]
- Vaalamo, M.; Leivo, T.; Saarialho-Kere, U. Differential expression of tissue inhibitors of metalloproteinases (TIMP-1,-2,-3, and-4) in normal and aberrant wound healing. Hum. Pathol. 1999, 30, 795–802. [Google Scholar] [CrossRef]
- Vaalamo, M.; Weckroth, M.; Puolakkainen, P.; Kere, J.; Saarinen, P.; Lauharanta, J.; SaarialhoKere, U.K. Patterns of matrix metalloproteinase and TIMP-1 expression in chronic and normally healing human cutaneous wounds. Br. J. Dermatol. 1996, 135, 52–59. [Google Scholar] [CrossRef]
- Endo, K.; Takino, T.; Miyamori, H.; Kinsen, H.; Yoshizaki, T.; Furukawa, M.; Sato, H. Cleavage of syndecan-1 by membrane type matrix metalloproteinase-1 stimulates cell migration. J. Biol. Chem. 2003, 278, 40764–40770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terasaki, K.; Kanzaki, T.; Aoki, T.; Iwata, K.; Saiki, I. Effects of recombinant human tissue inhibitor of metalloproteinases-2 (rh-TIMP-2) on migration of epidermal keratinocytes in vitro and wound healing in vivo. J. Dermatol. 2003, 30, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Liarte, S.; Bernabé-García, Á.; Nicolás, F.J. Role of TGF-β in Skin Chronic Wounds: A Keratinocyte Perspective. Cells 2020, 9, 306. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Sequence | Reverse Sequence | cDNA | TA | Cycles | Size |
|---|---|---|---|---|---|---|
| 18s | GGACAGGATTGACAGATTGAT | AGTCTCGTTCGTTATCGGAAT | 20 ng | 56 °C | 25 | 111 bp |
| RPL13a | AAGTACCAGGCAGTGACAG | CCTGTTTCCGTAGCCTCATG | 20 ng | 56 °C | 30 | 100 bp |
| TGF-β1 | TCCGGACCAGCCCTCGGGAG | CGGTCGCGGGTGCTGTTGTA | 20 ng | 58 °C | 35 | 680 bp |
| TGF-β2 | GCAGGTATTGATGGCACCTC | AGGCAGCAATTATCCTGCAC | 20 ng | 58 °C | 35 | 206 bp |
| TGF-β3 | CAGCTGCCTTGCCACCCCTC | TGCAGCCTTCCTCCCTCTCCC | 20 ng | 58 °C | 40 | 601 bp |
| CTGF | CCAATGACAACGCCTCCTG | TGGTGCAGCCAGAAAGCTC | 40 ng | 62 °C | 35 | 159 bp |
| FN1 | CCCCATTCCAGGACACTTCTG | GCCCACGGTAACAACCTCTT | 20 ng | 60 °C | 35 | 203 bp |
| COL1A1 | CAGCCGCTTCACCTACAGC | TTTTGTATTCAATCACTGTCTTGCC | 40 ng | 60 °C | 35 | 83 bp |
| MMP1 | CCCAGCGACTCTAGAAACACA | TCTTGGCAAATCTGGCGTGT | 40 ng | 60 °C | 35 | 322 bp |
| MMP2 | AACATACAAAGGGATTGCCAGGA | TTCAGCACAAACAGGTTGCAG | 40 ng | 60 °C | 35 | 117 bp |
| MMP9 | TCTATGGTCCTCGCCCTGAA | CATCGTCCACCGGACTCAAA | 40 ng | 64 °C | 35 | 219 bp |
| MMP13 | CCTGCTGGCTCATGCTTTTC | AGACCTAAGGAGTGGCCGAA | 40 ng | 64 °C | 40 | 141 bp |
| TIMP1 | AGTTTTGTGGCTCCCTGGAA | AAGCCCTTTTCAGAGCCTTG | 40 ng | 60 °C | 35 | 179 bp |
| TIMP2 | ATGCAGATGTAGTGATCAGGGC | GTGATGTGCATCTTGCCGTC | 40 ng | 60 °C | 35 | 256 bp |
| Smad7 | TTCGGACAACAAGAGTCAGC | AAGCCTTGATGGAGAAACC | 20 ng | 60 °C | 40 | 200 bp |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Rinderknecht, H.; Histing, T.; Kolbenschlag, J.; Nussler, A.K.; Ehnert, S. Establishment of an In Vitro Scab Model for Investigating Different Phases of Wound Healing. Bioengineering 2022, 9, 191. https://doi.org/10.3390/bioengineering9050191
Liu C, Rinderknecht H, Histing T, Kolbenschlag J, Nussler AK, Ehnert S. Establishment of an In Vitro Scab Model for Investigating Different Phases of Wound Healing. Bioengineering. 2022; 9(5):191. https://doi.org/10.3390/bioengineering9050191
Chicago/Turabian StyleLiu, Chao, Helen Rinderknecht, Tina Histing, Jonas Kolbenschlag, Andreas K. Nussler, and Sabrina Ehnert. 2022. "Establishment of an In Vitro Scab Model for Investigating Different Phases of Wound Healing" Bioengineering 9, no. 5: 191. https://doi.org/10.3390/bioengineering9050191
APA StyleLiu, C., Rinderknecht, H., Histing, T., Kolbenschlag, J., Nussler, A. K., & Ehnert, S. (2022). Establishment of an In Vitro Scab Model for Investigating Different Phases of Wound Healing. Bioengineering, 9(5), 191. https://doi.org/10.3390/bioengineering9050191