

Fighting Fire with Fire: Exosomes and Acute Pancreatitis-Associated Acute Lung Injury

 ,
,
Abstract
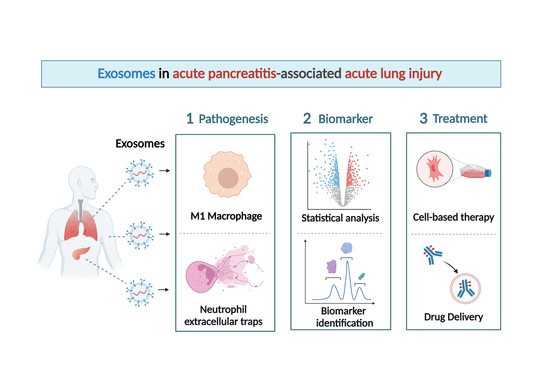
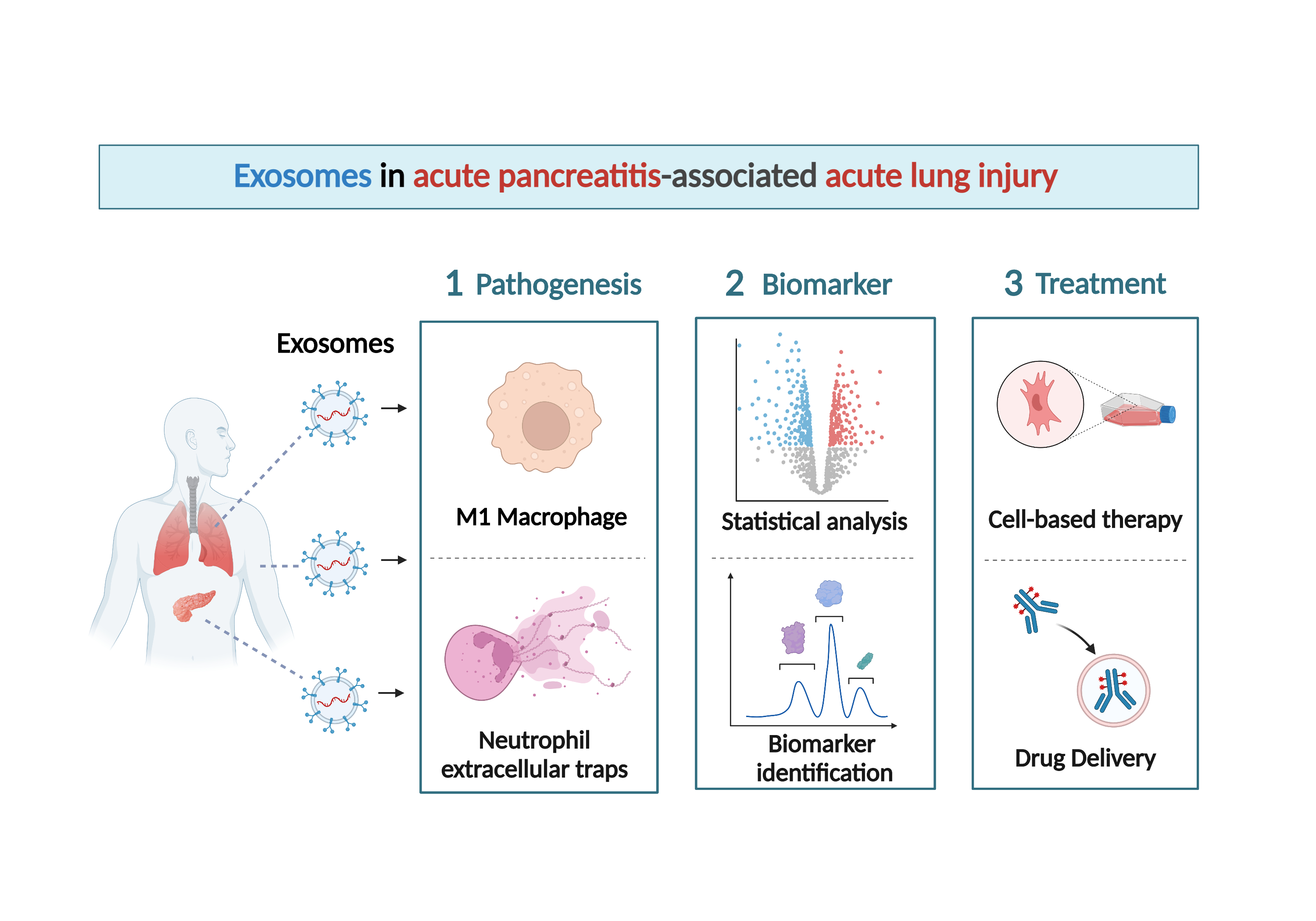
1. Introduction
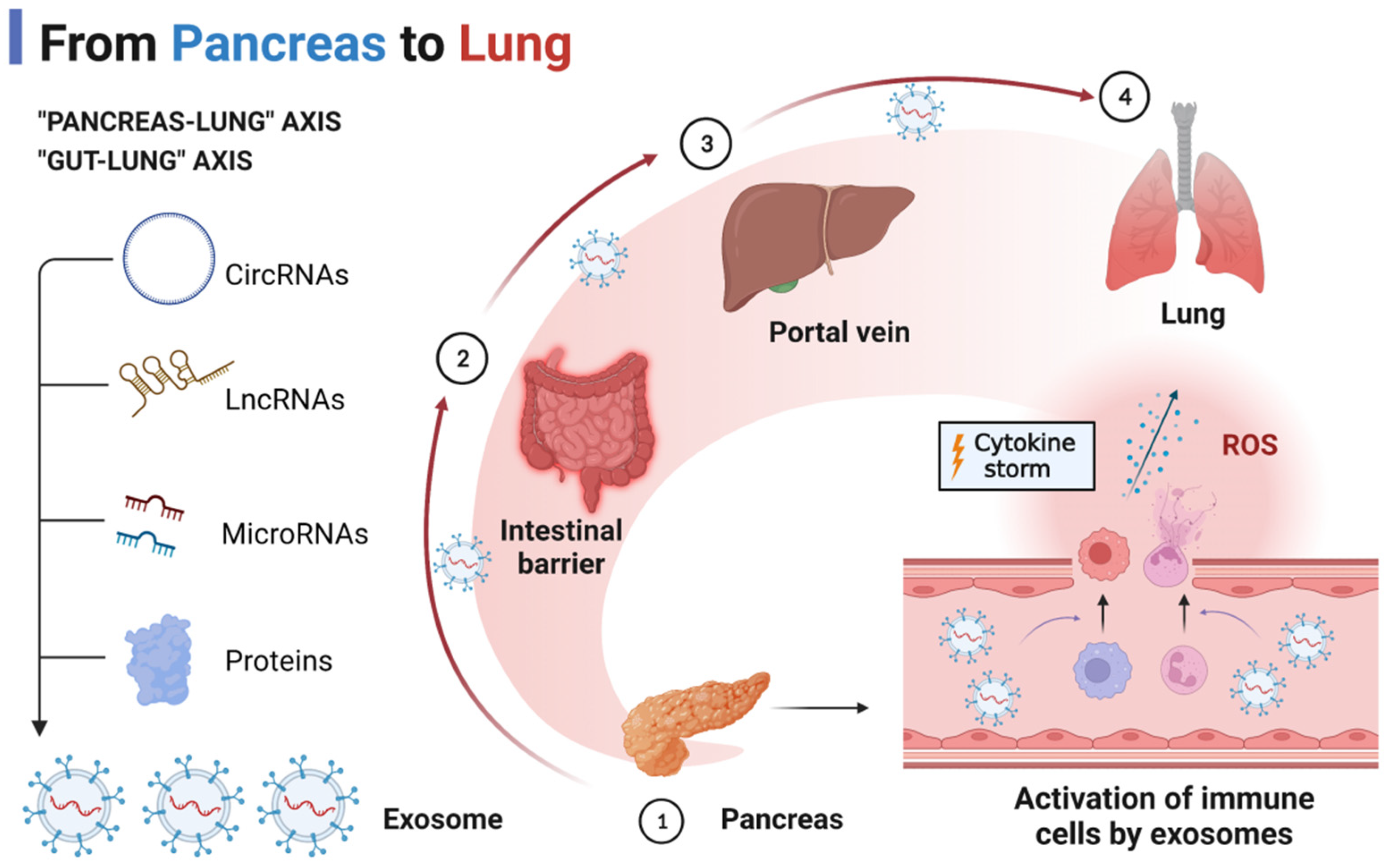
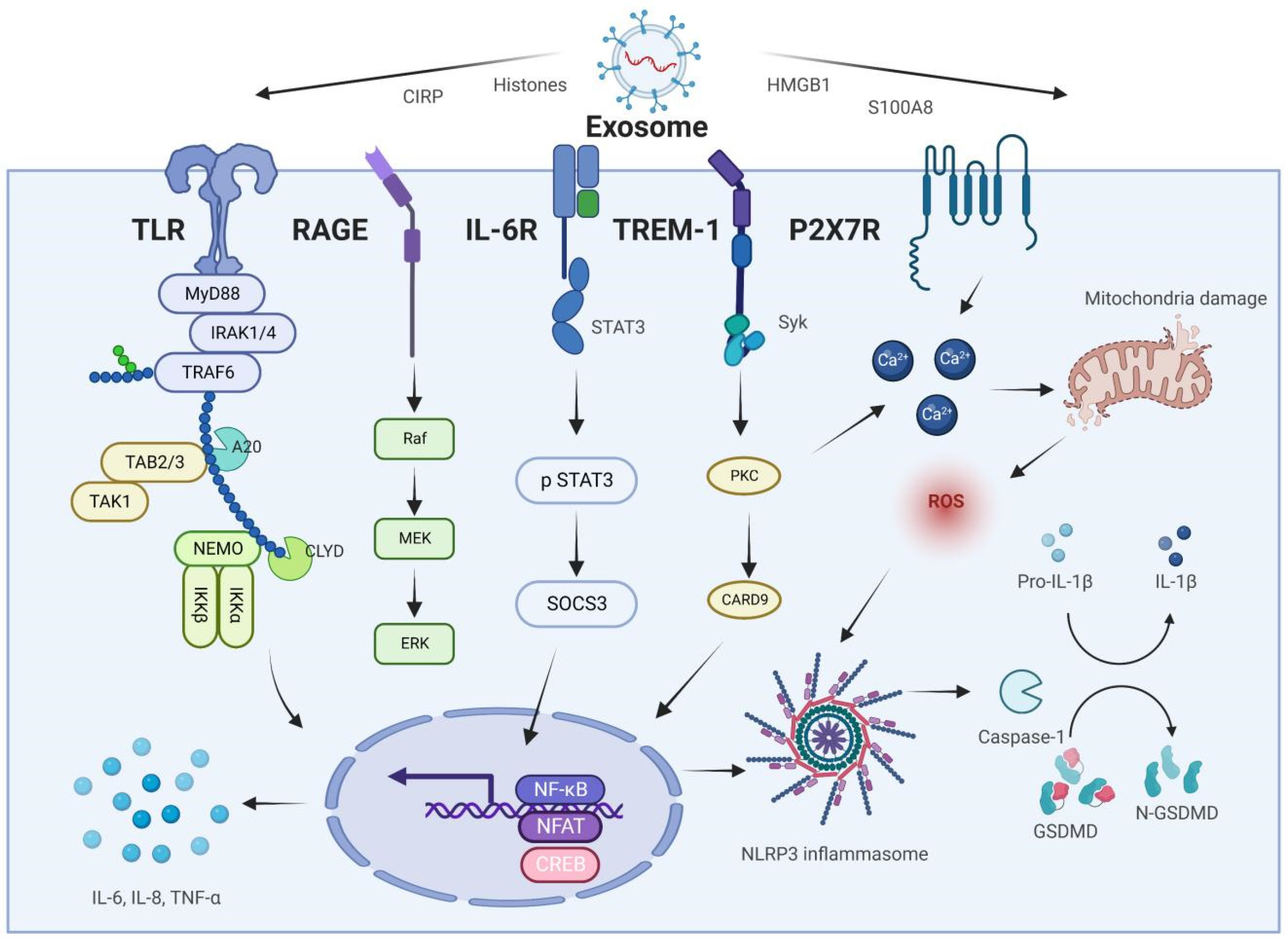
2. Exosomes and AP-Associated ALI
3. Exosome-Specific ncRNAs
3.1. LncRNAs
3.1.1. MALAT1
3.1.2. TUG1
3.1.3. Others
3.2. MiRNAs
3.2.1. MiR-155
3.2.2. MiR-21
3.2.3. MiR-216a
3.2.4. Others
4. Exosome-Specific Proteins
4.1. HMGB1
4.2. S100A8/A9
4.3. CIRP
4.4. Histones
5. The Therapeutic Potential of Exosomes in AP and Associated ALI
6. Exosome-Based Diagnostic Strategy
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AP | acute pancreatitis |
| SAP | severe acute pancreatitis |
| ALI | acute lung injury |
| PMVECs | pulmonary microvascular endothelial cells |
| AECs | alveolar epithelial cells |
| ARDS | acute respiratory distress syndrome |
| SIRS | systemic inflammatory response |
| MODS | multiple organ dysfunction syndromes |
| NcRNAs | non-coding RNAs |
| DAMPs | damage-associated molecular patterns |
| PAMPs | pathogen-associated molecular patterns |
| MVB | multivesicular body |
| ILVs | intraluminal vesicles |
| MiRNAs | microRNAs |
| CIRP | cold-inducible RNA-binding protein |
| AM | alveolar macrophage |
| IECs | intestinal epithelial cells |
| LncRNAs | long non-coding RNAs |
| MALAT1 | Metastasis-associated lung adenocarcinoma transcript 1 |
| YAP1 | yes-associated protein 1 |
| HMGB1 | high-mobility group box 1 |
| TLR4 | toll-like receptor 4 |
| NF-κB | nuclear factor kappa B |
| TUG1 | taurine upregulated gene 1 |
| LPS | lipopolysaccharide |
| SIRT1 | silent information regulator 1 |
| PVT1 | plasmacytoma variant translocation gene 1 |
| NLRP3 | NOD-like receptor protein 3 |
| JAK2 | Janus protein tyrosine kinase 2 |
| NETs | neutrophil extracellular traps |
| TLR | toll-like receptor |
| NAPDH | nicotinamide adenine dinucleotide phosphate |
| MMP | matrix metalloproteinase |
| ROS | reactive oxygen species |
| TREM-1 | triggering receptor expressed on myeloid cells-1 |
| SOFA | sequential organ failure assessment scores |
| PPAR | peroxisome proliferator-activated receptor |
| MSC | mesenchymal stem cell |
| GELN | ginger exosome-like nanoparticles |
| BALF | bronchoalveolar lavage fluid |
References
- Leppäniemi, A.; Tolonen, M.; Tarasconi, A.; Lohse, H.A.S.; Gamberini, E.; Kirkpatrick, A.W.; Ball, C.G.; Parry, N.; Sartelli, M.; Wolbrink, D.R.J.; et al. 2019 WSES guidelines for the management of severe acute pancreatitis. World J. Emerg. Surg. 2019, 14, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Iannuzzi, J.P.; King, J.A.; Leong, J.H.; Quan, J.; Windsor, J.W.; Tanyingoh, D.; Stephanie, C.; Nauzer, F.; Steven, J.H.; Abdel-Aziz, S.; et al. Global Incidence of Acute Pancreatitis Is Increasing Over Time: A Systematic Review and Meta-Analysis. Gastroenterology 2022, 162, 122–134. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Zhang, J.; Chen, H.; Wang, C.; Qiu, Y.; Liu, Y.; Wan, J.; Guo, H. Effects and mechanisms of alveolar type II epithelial cell apoptosis in severe pancreati-tis-induced acute lung injury. Exp. Ther. Med. 2014, 7, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Vrolyk, V.; Singh, B. Animal models to study the role of pulmonary intravascular macrophages in spontaneous and induced acute pancreatitis. Cell Tissue Res. 2020, 380, 207–222. [Google Scholar] [CrossRef] [PubMed]
- Luiken, I.; Eisenmann, S.; Garbe, J.; Sternby, H.; Verdonk, R.C.; Dimova, A.; Ignatavicius, P.; Ilzarbe, L.; Koiva, P.; Penttilä, A.K.; et al. Pleuropulmonary pathologies in the early phase of acute pancreatitis correlate with disease severity. PLoS ONE 2022, 17, e0263739. [Google Scholar] [CrossRef]
- Ge, P.; Luo, Y.; Okoye, C.S.; Chen, H.; Liu, J.; Zhang, G.; Xu, C.; Chen, H. Intestinal barrier damage, systemic inflammatory response syndrome, and acute lung injury: A troublesome trio for acute pancreatitis. Biomed. Pharmacother. 2020, 132, 110770. [Google Scholar] [CrossRef]
- Liu, D.; Wen, L.; Wang, Z.; Hai, Y.; Yang, D.; Zhang, Y.; Bai, M.; Song, B.; Wang, Y. The Mechanism of Lung and Intestinal Injury in Acute Pancreatitis: A Review. Front. Med. 2022, 9, 904078. [Google Scholar] [CrossRef]
- Suire, C.N.; Hade, M.D. Extracellular Vesicles in Type 1 Diabetes: A Versatile Tool. Bioengineering 2022, 9, 105. [Google Scholar] [CrossRef]
- Hade, M.; Suire, C.; Suo, Z. Mesenchymal Stem Cell-Derived Exosomes: Applications in Regenerative Medicine. Cells 2021, 10, 1959. [Google Scholar] [CrossRef]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): A position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef]
- Chargaff, E.; West, R. The Biological Significance of the Thromboplastic Protein of Blood. J. Biol. Chem. 1946, 166, 189–197. [Google Scholar] [CrossRef]
- Webbera, J.; Johnsons, A. Platelet participation in blood coagulation aspects of hemostasis. Am. J. Pathol. 1970, 60, 19–42. [Google Scholar]
- Fan, Y.; Li, Z.; He, Y. Exosomes in the Pathogenesis, Progression, and Treatment of Osteoarthritis. Bioengineering 2022, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Lei, B.; Zhang, E.; Gong, P.; Gu, J.; He, L.; Han, L.; Yuan, Z. Targeted Therapy for Inflammatory Diseases with Mesenchymal Stem Cells and Their Derived Exosomes: From Basic to Clinics. Int. J. Nanomed. 2022, 17, 1757–1781. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Bi, J.; Huang, J.; Tang, Y.; Du, S.; Li, P. Exosome: A Review of Its Classification, Isolation Techniques, Storage, Diagnostic and Targeted Therapy Applications. Int. J. Nanomed. 2020, 15, 6917–6934. [Google Scholar] [CrossRef]
- Im, H.; Lee, K.; Weissleder, R.; Lee, H.; Castro, C.M. Novel nanosensing technologies for exosome detection and profiling. Lab Chip 2017, 17, 2892–2898. [Google Scholar] [CrossRef]
- Kalra, H.; Adda, C.G.; Liem, M.; Ang, C.-S.; Mechler, A.; Simpson, R.J.; Hulett, M.D.; Mathivanan, S. Comparative proteomics evaluation of plasma exosome isolation techniques and assessment of the stability of exosomes in normal human blood plasma. Proteomics 2013, 13, 3354–3364. [Google Scholar] [CrossRef]
- Ge, Q.; Zhou, Y.; Lu, J.; Bai, Y.; Xie, X.; Lu, Z. miRNA in Plasma Exosome is Stable under Different Storage Conditions. Molecules 2014, 19, 1568–1575. [Google Scholar] [CrossRef]
- AzamAnsari, M.; Thiruvengadam, M.; Venkidasamy, B.; Alomary, M.N.; Salawi, A.; Chung, I.-M.; Shariati, M.A.; Rebezo, M. Exosome-based nanomedicine for cancer treatment by tar-geting inflammatory pathways: Current status and future perspectives. In Seminars in Cancer Biology; Academic Press: Cambridge, MA, USA, 2022. [Google Scholar]
- Jia, Y.-C.; Ding, Y.-X.; Mei, W.-T.; Wang, Y.-T.; Zheng, Z.; Qu, Y.-X.; Liang, K.; Li, J.; Cao, F.; Li, F. Extracellular vesicles and pancreatitis: Mechanisms, status and perspectives. Int. J. Biol. Sci. 2021, 17, 549–561. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, J.; Li, F.; Luo, Y.; Ge, P.; Zhang, Y.; Wen, H.; Yang, Q.; Ma, S.; Chen, H. The gut-lung axis in severe acute Pancreatitis-associated lung injury: The protection by the gut microbiota through short-chain fatty acids. Pharmacol. Res. 2022, 182, 106321. [Google Scholar] [CrossRef]
- Zhu, H.; Song, Y.; Kong, X.; Du, Y. Expression of microrNA-216A in patients with acute pancreatitis complicated with lung injury and its effect on endothelial cell permeability. J. Chin. Pancreas Dis. 2020, 20, 4. [Google Scholar]
- Xu, Q.; Wang, M.; Guo, H.; Liu, H.; Zhang, G.; Xu, C.; Chen, H. Emodin Alleviates Severe Acute Pancreatitis-Associated Acute Lung Injury by Inhibiting the Cold-Inducible RNA-Binding Protein (CIRP)-Mediated Activation of the NLRP3/IL-1β/CXCL1 Signaling. Front. Pharmacol. 2021, 12, 655372. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.Y.; Jia, T.X.; Zhang, M. Intestinal bacterial overgrowth in the early stage of severe acute pancreatitis is associated with acute respiratory distress syndrome. World J. Gastroenterol. 2021, 27, 1643–1654. [Google Scholar] [CrossRef]
- Jabłońska, B.; Mrowiec, S. Nutritional Support in Patients with Severe Acute Pancreatitis-Current Standards. Nutrients 2021, 13, 1498. [Google Scholar] [CrossRef]
- Kanthasamy, K.A.; Akshintala, V.S.; Singh, V.K. Nutritional Management of Acute Pancreatitis. Gastroenterol. Clin. N. Am. 2021, 50, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Sonika, U.; Moka, P.; Sharma, B.; Sachdev, V.; Mishra, S.K.; Upadhyay, A.D.; Saraya, A. Association of endotoxaemia & gut permeability with complications of acute pancre-atitis: Secondary analysis of data. Indian J. Med. Res. 2019, 149, 763–770. [Google Scholar]
- Hu, X.; Gong, L.; Zhou, R.; Han, Z.; Ji, L.; Zhang, Y.; Zhang, S.; Wu, D. Variations in Gut Microbiome are Associated with Prognosis of Hypertriglyceridem-ia-Associated Acute Pancreatitis. Biomolecules 2021, 11, 695. [Google Scholar] [CrossRef]
- Tang, Y.; Kong, J.; Zhou, B.; Wang, X.; Liu, X.; Wang, Y.; Zhu, S. Mesenteric Lymph Duct Ligation Alleviates Acute Lung Injury Caused by Severe Acute Pancreatitis Through Inhibition of High Mobility Group Box 1-Induced Inflammation in Rats. Am. J. Dig. Dis. 2021, 66, 4344–4353. [Google Scholar] [CrossRef]
- Park, E.J.; Shimaoka, M.; Kiyono, H. Functional Flexibility of Exosomes and MicroRNAs of Intestinal Epithelial Cells in Affecting Inflammation. Front. Mol. Biosci. 2022, 9, 854487. [Google Scholar] [CrossRef]
- Dahariya, S.; Paddibhatla, I.; Kumar, S.; Raghuwanshi, S.; Pallepati, A.; Gutti, R.K. Long non-coding RNA: Classification, biogenesis and functions in blood cells. Mol. Immunol. 2019, 112, 82–92. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, X.-J.; Ji, Y.-X.; Zhang, P.; Deng, K.-Q.; Gong, J.; Ren, S.; Wang, X.; Chen, I.; Wang, H.; et al. The long noncoding RNA Chaer defines an epigenetic checkpoint in cardiac hypertrophy. Nat. Med. 2016, 22, 1131–1139. [Google Scholar] [CrossRef] [PubMed]
- Engreitz, J.M.; Haines, J.E.; Perez, E.; Munson, G.; Chen, J.; Kane, M.; McDonel, P.E.; Guttman, M.; Lander, E.S. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature 2016, 539, 452–455. [Google Scholar] [CrossRef] [PubMed]
- Dykes, I.M.; Emanueli, C. Transcriptional and Post-transcriptional Gene Regulation by Long Non-coding RNA. Genom. Proteom. Bioinform. 2017, 15, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhou, D.; Li, G.; Ming, X.; Tu, Y.F.; Tian, J.; Lu, H.; Yu, B. Long Non Coding RNA-UCA1 Contributes to Cardiomyocyte Apoptosis by Suppression of p27 Expression. Cell. Physiol. Biochem. 2015, 35, 1986–1998. [Google Scholar] [CrossRef]
- Senmatsu, S.; Hirota, K. Roles of lncRNA transcription as a novel regulator of chromosomal function. Genes Genet. Syst. 2020, 95, 213–223. [Google Scholar] [CrossRef]
- Luo, Y.; Ge, P.; Wang, M.; Chen, H.; Liu, J.; Wei, T.; Jiang, Y.; Qu, J.; Chen, H. Research progress of DLX6-AS1 in human cancers. Hum. Cell 2021, 34, 1642–1652. [Google Scholar] [CrossRef]
- Zaki, A.; Ali, M.S.; Hadda, V.; Ali, S.M.; Chopra, A.; Fatima, T. Long non-coding RNA (lncRNA): A potential therapeutic target in acute lung injury. Genes Dis. 2021, 9, 1258–1268. [Google Scholar] [CrossRef]
- Yao, Z.; Yang, Y.; Sun, M.; He, Y.; Liao, L.; Chen, K.; Li, B. New insights into the interplay between long non-coding RNAs and RNA-binding proteins in cancer. Cancer Commun. 2022, 42, 117–140. [Google Scholar] [CrossRef]
- Lai, X.; Zhong, J.; Zhang, A.; Zhang, B.; Zhu, T.; Liao, R. Focus on long non-coding RNA MALAT1: Insights into acute and chronic lung diseases. Front. Genet. 2022, 13, 2704. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Khoshbakht, T.; Hussen, B.M.; Taheri, M.; Arefian, N. Regulatory Role of Non-Coding RNAs on Immune Responses During Sepsis. Front. Immunol. 2021, 12, 798713. [Google Scholar] [CrossRef]
- Su, K.; Wang, N.; Shao, Q.; Liu, H.; Zhao, B.; Ma, S. The role of a ceRNA regulatory network based on lncRNA MALAT1 site in cancer progression. Biomed. Pharmacother. 2021, 137, 111389. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Liu, J.; Xu, D.; Lu, Y. Reciprocal Feedback Loop of the MALAT1-MicroRNA-194-YAP1 Pathway Regulates Progression of Acute Pancreatitis. Med. Sci. Monit. 2019, 25, 6894–6904. [Google Scholar] [CrossRef]
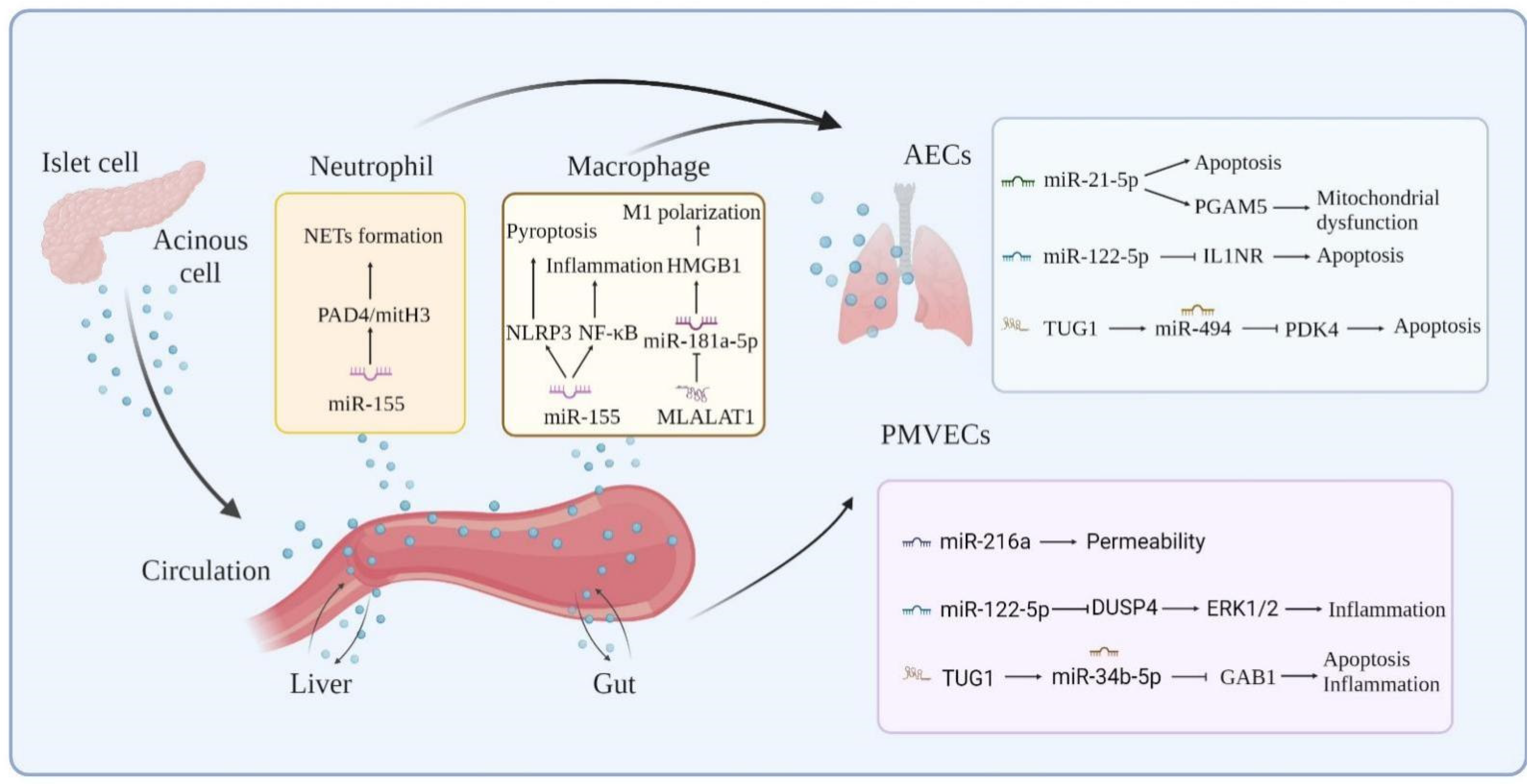
- Liu, J.; Niu, Z.; Zhang, R.; Peng, Z.; Wang, L.; Liu, Z.; Gao, Y.; Pei, H.; Pan, L. MALAT1 shuttled by extracellular vesicles promotes M1 polarization of macrophages to induce acute pancreatitis via miR-181a-5p/HMGB1 axis. J. Cell. Mol. Med. 2021, 25, 9241–9254. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Yao, W.; Liu, K.; Wang, G.; Chen, B.; Wang, Z.; Hou, S. Nonalcoholic Fatty Liver Hepatocyte-Derived lncRNA MALAT1 Aggravates Pancreatic Cell Inflammation via the Inhibition of Autophagy by Upregulating YAP. Comput. Intell. Neurosci. 2022, 2022, 2930960. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lin, Y. Long non-coding RNA plasmacytoma variant translocation 1 correlates with higher inflammation, multiple organ injury and mortality risk in acute pancreatitis, especially in severe acute pancreatitis. Clin. Res. Hepatol. Gastroenterol. 2022, 46, 101870. [Google Scholar] [CrossRef]
- Liu, W.; Geng, F.; Yu, L. Long non-coding RNA MALAT1/microRNA 125a axis presents excellent value in discriminating sepsis patients and exhibits positive association with general disease severity, organ injury, inflammation level, and mortality in sepsis patients. J. Clin. Lab. Anal. 2020, 34, e23222. [Google Scholar] [CrossRef]
- Guo, C.; Qi, Y.; Qu, J.; Gai, L.; Shi, Y.; Yuan, C. Pathophysiological Functions of the lncRNA TUGCurr. Pharm. Des. 2020, 26, 688–700. [Google Scholar] [CrossRef]
- Yang, L.; Zhao, L.; Zhang, H.; Chen, P. Up-regulation of TUG1 can regulate miR-494/PDK4 axis to inhibit LPS-induced acute lung injury caused by sepsis. Am. J. Transl. Res. 2021, 13, 12375–12385. [Google Scholar]
- Qiu, N.; Xu, X.; He, Y. LncRNA TUG1 alleviates sepsis-induced acute lung injury by targeting miR-34b-5p/GABBMC. Pulm. Med. 2020, 20, 1–12. [Google Scholar] [CrossRef]
- Ma, W.; Zhang, W.; Cui, B.; Gao, J.; Liu, Q.; Yao, M.; Ning, H.; Xing, L. Functional delivery of lncRNA TUG1 by endothelial progenitor cells derived extracellular vesicles confers anti-inflammatory macrophage polarization in sepsis via impairing miR-9-5p-targeted SIRT1 inhibition. Cell Death Dis. 2021, 12, 1056. [Google Scholar] [CrossRef]
- Wen, X.; He, B.; Tang, X.; Wang, B.; Chen, Z. Emodin inhibits the progression of acute pancreatitis via regulation of lncRNA TUG1 and exosomal lncRNA TUGMol. Med. Rep. 2021, 24, 785. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Peng, H.; Gui, F. Long noncoding plasmacytoma variant translocation 1 facilitates the surveillance of acute respiratory distress syndrome and mortality prediction in sepsis. Biomark. Med. 2021, 15, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Li, J.; Guo, M.; Bai, R.; Lei, G.; Sun, H.; Tong, S.; He, K.; He, L. Aberrant expressions of MIAT and PVT1 in serum exosomes of schizophrenia patients. Schizophr. Res. 2021, 240, 71–72. [Google Scholar] [CrossRef] [PubMed]
- Ma, Q.; Gan, G.-F.; Niu, Y.; Tong, S.-J. Analysis of associations of FBXL19-AS1 with occurrence, development and prognosis of acute pancreatitis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12763–12769. [Google Scholar] [PubMed]
- Li, J.; Bu, X.; Chen, X.; Xiong, P.; Chen, Z.; Yu, L. Predictive value of long non-coding RNA intersectin 1-2 for occurrence and in-hospital mortality of severe acute pancreatitis. J. Clin. Lab. Anal. 2019, 34, e23170. [Google Scholar] [CrossRef] [PubMed]
- Maiese, A.; Scatena, A.; Costantino, A.; Chiti, E.; Occhipinti, C.; La Russa, R.; Di Paolo, M.; Turillazzi, E.; Frati, P.; Fineschi, V. Expression of MicroRNAs in Sepsis-Related Organ Dysfunction: A Systematic Review. Int. J. Mol. Sci. 2022, 23, 9354. [Google Scholar] [CrossRef] [PubMed]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA Translation and Stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from Repression to Activation: MicroRNAs Can Up-Regulate Translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed]
- Duchaine, T.F.; Fabian, M.R. Mechanistic Insights into MicroRNA-Mediated Gene Silencing. Cold Spring Harb. Perspect. Biol. 2019, 11, a032771. [Google Scholar] [CrossRef]
- Lopez, J.G.; Brieño-Enriquez, M.A.; DEL Mazo, J. MicroRNA biogenesis and variability. Biomol. Concepts 2013, 4, 367–380. [Google Scholar] [CrossRef]
- Zlotorynski, E. Insights into the kinetics of microRNA biogenesis and turnover. Nat. Rev. Mol. Cell Biol. 2019, 20, 511. [Google Scholar] [CrossRef] [PubMed]
- Treiber, T.; Treiber, N.; Meister, G. Regulation of microRNA biogenesis and its crosstalk with other cellular pathways. Nat. Rev. Mol. Cell Biol. 2019, 20, 5–20. [Google Scholar] [CrossRef]
- Zhang, J.; Li, S.; Li, L.; Li, M.; Guo, C.; Yao, J.; Mi, S. Exosome and Exosomal MicroRNA: Trafficking, Sorting, and Function. Genom. Proteom. Bioinform. 2015, 13, 17–24. [Google Scholar] [CrossRef]
- Dawson, O.; Piccinini, A.M. miR-155-3p: Processing by-product or rising star in immunity and cancer? Open Biol. 2022, 12, 220070. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Huang, S.; Liu, X.; Zhang, Y.; Wei, S.; Hu, X. miR-155: An Important Role in Inflammation Response. J. Immunol. Res. 2022, 2022, 7437281. [Google Scholar] [CrossRef] [PubMed]
- Tacke, F.; Spehlmann, M.E.; Vucur, M.; Benz, F.; Luedde, M.; Cardenas, D.V.; Roy, S.; Loosen, S.; Hippe, H.-J.; Frey, N.; et al. miR-155 Predicts Long-Term Mortality in Critically Ill Patients Younger than 65 Years. Mediat. Inflamm. 2019, 2019, 6714080. [Google Scholar] [CrossRef]
- Obradovic, D.; Rommel, K.; Blazek, S.; Klingel, K.; Gutberlet, M.; Lücke, C.; Büttner, P.; Thiele, H.; Adams, V.; Lurz, P.; et al. The potential role of plasma miR-155 and miR-206 as circulatory biomarkers in inflammatory cardiomyopathy. ESC Hear. Fail. 2021, 8, 1850–1860. [Google Scholar] [CrossRef]
- Lin, S.; Chen, X.; Wang, X. Expression and clinical significance of Mir-155 in peripheral blood of patients with acute pan-creatitis. World Chin. Dig. J. 2015, 23, 3935–3939. [Google Scholar] [CrossRef]
- Wang, D.; Tang, M.; Zong, P.; Liu, H.; Zhang, T.; Liu, Y.; Zhao, Y. MiRNA-155 Regulates the Th17/Treg Ratio by Targeting SOCS1 in Severe Acute Pancreatitis. Front. Physiol. 2018, 9, 686. [Google Scholar] [CrossRef]
- Miao, B.; Wang, J.; Zhong, J.; Pan, E.; Lin, S. Exosomal Mir-155 affects severe acute pancreatitis lung injury by regulating SOCS. Chin. Foreign Med. 2021, 40, 23–27. [Google Scholar]
- Hu, L.; Han, D.; Yu, D.; Ao, D.; Yang, Z. Circulating Blood miR-155 and miR-21 Promote the Development of Acute Pancreatitis and Can Be Used to Assess the Risk Stratification of Pancreatitis. J. Health Eng. 2021, 2021, 2064162. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Wang, R.L.; Xie, H.; Jin, W.; Yu, K.L. Overexpressed miRNA-155 dysregulates intestinal epithelial apical junctional complex in severe acute pancreatitis. World J. Gastroenterol. 2013, 19, 8282–8291. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Yang, X.; Ren, Y.; Li, X.; Zhu, Y.; Haddock, A.N.; Ji, B.; Xia, L.; Lu, N. Inhibition of miR-155 reduces impaired autophagy and improves prognosis in an experimental pancreatitis mouse model. Cell Death Dis. 2019, 10, 303. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chu, J.; Sun, H.; Zhao, D.; Ma, B.; Xue, D.; Zhang, W.; Li, Z. MiR-155 aggravates impaired autophagy of pancreatic acinar cells through targeting Rictor. Acta Biochim. Biophys. Sin. 2020, 52, 192–199. [Google Scholar] [CrossRef]
- Jiménez-Alesanco, A.; Marcuello, M.; Pastor-Jiménez, M.; López-Puerto, L.; Bonjoch, L.; Gironella, M.; Carrascal, M.; Abian, J.; de-Madaria, E.; Closa, D. Acute pancreatitis promotes the generation of two different ex-osome populations. Sci. Rep. 2019, 9, 19887. [Google Scholar] [CrossRef]
- Wu, X.-B.; Sun, H.-Y.; Luo, Z.-L.; Cheng, L.; Duan, X.-M.; Ren, J.-D. Plasma-derived exosomes contribute to pancreatitis-associated lung injury by triggering NLRP3-dependent pyroptosis in alveolar macrophages. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2020, 1866, 165685. [Google Scholar] [CrossRef]
- Jiang, K.; Yang, J.; Guo, S.; Zhao, G.; Wu, H.; Deng, G. Peripheral Circulating Exosome-Mediated Delivery of miR-155 as a Novel Mechanism for Acute Lung Inflammation. Mol. Ther. 2019, 27, 1758–1771. [Google Scholar] [CrossRef]
- Huang, Z.; Huang, H.; Shen, M.; Li, C.; Liu, C.; Zhu, H.; Zhang, W. MicroRNA-155-5p modulates the progression of acute respiratory distress syndrome by targeting interleukin receptors. Bioengineered 2022, 13, 11732–11741. [Google Scholar] [CrossRef]
- Hawez, A.; Taha, D.; Algaber, A.; Madhi, R.; Rahman, M.; Thorlacius, H. MiR-155 regulates neutrophil extracellular trap formation and lung injury in abdominal sepsis. J. Leukoc. Biol. 2021, 111, 391–400. [Google Scholar] [CrossRef]
- Wu, K.; Wang, L.; Fu, G.; Zheng, Y. Expression and clinical significance of microRNA-21-3p and microRNA-551-5p in patients with acute pancreatitis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2020, 32, 463–467. [Google Scholar]
- Nakamaru, K.; Tomiyama, T.; Kobayashi, S.; Ikemune, M.; Tsukuda, S.; Ito, T.; Tanaka, T.; Yamaguchi, T.; Ando, Y.; Ikeur, T.; et al. Extracellular vesicles microRNA analysis in type 1 autoimmune pan-creatitis: Increased expression of microRNA. Pancreatology 2020, 20, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Dixit, A.K.; Sarver, A.E.; Yuan, Z.; George, J.; Barlass, U.; Cheema, H.; Sareen, A.; Banerjee, S.; Dudeja, V.; Dawra, R.; et al. Comprehensive analysis of microRNA signature of mouse pancreatic acini: Over-expression of miR-21-3p in acute pancreatitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 311, G974–G980. [Google Scholar] [CrossRef]
- Wang, T.; Jiang, L.; Wei, X.; Liu, B.; Zhao, J.; Xie, P.; Yang, B.; Wang, L. MiR-21-3p aggravates injury in rats with acute hemorrhagic necrotizing pancreatitis by activating TRP signaling pathway. Biomed. Pharmacother. 2018, 107, 1744–1753. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Yu, J.; Gao, W.; Sun, Y.; Liu, X.; Lv, Z.; Li, L.; Xue, D. The lncRNA TCONS_00021785/miR-21-5p/Trim33 axis regulates VMP1-mediated zymophagy, reduces the activation of trypsinogen, and promotes acinar cell recovery. Cell Death Discov. 2022, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.; Chen, M.; Ji, H.; Liu, G.; Mei, H.; Li, K.; Chen, T. miR-21-5p regulates type II alveolar epithelial cell apoptosis in hyperoxic acute lung injury. Mol. Med. Rep. 2018, 17, 5796–5804. [Google Scholar] [CrossRef]
- Liu, G.; Qian, M.; Chen, M.; Chen, T.; Qin, S. miR-21-5p Suppresses Mitophagy to Alleviate Hyperoxia-Induced Acute Lung Injury by Directly Targeting PGAM5. BioMed Res. Int. 2020, 2020, 4807254. [Google Scholar] [CrossRef]
- Li, J.; Wei, L.; Han, Z.; Chen, Z. Mesenchymal stromal cells-derived exosomes alleviate ischemia/reperfusion injury in mouse lung by transporting anti-apoptotic miR-21-5p. Eur. J. Pharmacol. 2019, 852, 68–76. [Google Scholar] [CrossRef]
- Kong, X.; Du, Y.; Li, L.; Liu, J.; Wang, G.; Zhu, J.; Man, X.; Gong, Y.; Xiao, L.; Zheng, Y.; et al. Plasma miR-216a as a potential marker of pancreatic injury in a rat model of acute pancreatitis. World J. Gastroenterol. 2010, 16, 4599–4604. [Google Scholar] [CrossRef]
- Endo, K.; Weng, H.; Kito, N.; Fukushima, Y.; Iwai, N. miR-216a and miR-216b as markers for acute phased pancreatic injury. Biomed. Res. 2013, 34, 179–188. [Google Scholar] [CrossRef]
- Goodwin, D.; Rosenzweig, B.; Zhang, J.; Xu, L.; Stewart, S.; Thompson, K.; Rouse, R. Evaluation of miR-216a and miR-217 as potential biomarkers of acute pan-creatic injury in rats and mice. Biomarkers 2014, 19, 517–529. [Google Scholar] [CrossRef]
- Blenkiron, C.; Askelund, K.J.; Shanbhag, S.T.; Chakraborty, M.; Petrov, M.; Delahunt, B.; Windsor, J.A.; Phillips, A.R. MicroRNAs in mesenteric lymph and plasma during acute pancreatitis. Ann. Surg. 2014, 260, 341–347. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Park, H.; Choi, H.; Lee, S.; Lee, S.; Lee, J.; Cho, E.; Han, H.; Seok, J.; Son, W. Evaluation of Circulating MicroRNA Biomarkers in the Acute Pancreatic Injury Dog Model. Int. J. Mol. Sci. 2018, 19, 3048. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Deng, L.; Chen, W.; Shi, N.; Jin, T.; Lin, Z.; Ma, Y.; Jiang, K.; Yang, X.; Xia, Q. Circulating microRNA 216 as a Marker for the Early Identification of Severe Acute Pancreatitis. Am. J. Med. Sci 2017, 353, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Wang, C.; Yang, L.; Zhao, G.; Liu, H.; Li, J.; Chen, P.; Cui, K. Significance of microRNA 216a, 324–325p and 29a expression in peripheral blood in patients with acute pancreatitis and their correlation with liver injury. Zhonghua Yi Xue Za Zhi 2020, 100, 2126–2131. [Google Scholar]
- Kong, F.; Sun, Y.; Song, W.; Zhou, Y.; Zhu, S. MiR-216a alleviates LPS-induced acute lung injury via regulating JAK2/STAT3 and NF-κB signaling. Hum. Cell 2019, 33, 67–78. [Google Scholar] [CrossRef]
- Qu, Y.; Ding, Y.; Lu, J.; Jia, Y.; Bian, C.; Guo, Y.; Zheng, Z.; Mei, W.; Cao, F.; Li, F. Identification of key microRNAs in exosomes derived from patients with the severe acute pancreatitis. Asian J. Surg. 2022; in press. [Google Scholar] [CrossRef]
- Wang, F.; Li, J.; Li, L.; Chen, Z.; Wang, N.; Zhu, M.; Mi, H.; Xiong, Y.; Guo, G.; Gu, Y. Circular RNA circ_IRAK3 contributes to tumor growth through upregulating KIF2A via adsorbing miR-603 in breast cancer. Cancer Cell Int. 2022, 22, 81. [Google Scholar] [CrossRef]
- Lin, Y.; Wu, X.; Zheng, C.; Zhang, Q.; Zhang, G.; Chen, K.; Zhan, Q.; An, F. Mechanistic Investigation on the Regulation of FABP1 by the IL-6/miR-603 Signaling in the Pathogenesis of Hepatocellular Carcinoma. Biomed. Res. Int. 2021, 2021, 8579658. [Google Scholar] [CrossRef]
- Gao, X.; Yin, J.; Yao, Y. hsa_circ_0001955 Promotes Colorectal Cancer Progression by Regulating miR-583/FGF21 Axis. J. Oncol. 2022, 2022, 4288474. [Google Scholar] [CrossRef]
- Hu, Y.; Yu, Y. Dysregulation of miR-192-5p in acute pancreatitis patients with nonalcoholic fatty liver and its functional role in acute pancreatitis progression. Biosci. Rep. 2020, 40, BSR20194345. [Google Scholar] [CrossRef]
- Lu, Z.; Feng, H.; Shen, X.; He, R.; Meng, H.; Lin, W.; Geng, Q. MiR-122-5p protects against acute lung injury via regulation of DUSP4/ERK signaling in pulmonary microvascular endothelial cells. Life Sci. 2020, 256, 117851. [Google Scholar] [CrossRef]
- Li, J.; Zeng, X.; Wang, W. miR-122-5p downregulation attenuates lipopolysaccharide-induced acute lung injury by targeting, I.L.1.R.N. Exp. Ther. Med. 2021, 22, 1278. [Google Scholar] [CrossRef] [PubMed]
- Caserta, S.; Ghezzi, P. Release of redox enzymes and micro-RNAs in extracellular vesicles, during infection and inflammation. Free Radic. Biol. Med. 2021, 169, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Murao, A.; Aziz, M.; Wang, H.; Brenner, M.; Wang, P. Release mechanisms of major DAMPs. Apoptosis 2021, 26, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Block, H.; Rossaint, J.; Zarbock, A. The Fatal Circle of NETs and NET-Associated DAMPs Contributing to Organ Dys-function. Cells 2022, 11, 1919. [Google Scholar] [CrossRef] [PubMed]
- Deng, C.; Zhao, L.; Yang, Z.; Shang, J.-J.; Wang, C.-Y.; Shen, M.-Z.; Jiang, S.; Li, T.; Di, W.-C.; Chen, Y.; et al. Targeting HMGB1 for the treatment of sepsis and sepsis-induced organ injury. Acta Pharmacol. Sin. 2021, 43, 520–528. [Google Scholar] [CrossRef]
- Yasuda, T.; Ueda, T.; Takeyama, Y.; Shinzeki, M.; Sawa, H.; Nakajima, T.; Ajiki, T.; Fujino, Y.; Suzuki, Y.; Kuroda, Y. Significant Increase of Serum High-Mobility Group Box Chromosomal Protein 1 Levels in Patients With Severe Acute Pancreatitis. Pancreas 2006, 33, 359–363. [Google Scholar] [CrossRef]
- Xu, G.-F.; Guo, M.; Tian, Z.-Q.; Wu, G.-Z.; Zou, X.-P.; Zhang, W.-J. Increased of serum high-mobility group box chromosomal protein 1 correlated with intestinal mucosal barrier injury in patients with severe acute pancreatitis. World J. Emerg. Surg. 2014, 9, 61. [Google Scholar] [CrossRef]
- Arriaga-Pizano, L.; Boscó-Gárate, I.; Martínez-Ordaz, J.L.; Wong-Baeza, I.; Gutiérrez-Mendoza, M.; Sánchez-Fernandez, P.; Macías, C.I.R.L.; Isibasi, A.; Pelaez-Luna, M.; Cérbulo-Vázquez, A.; et al. High Serum Levels of High-Mobility Group Box 1 (HMGB1) and Low Levels of Heat Shock Protein 70 (Hsp70) are Associated with Poor Prognosis in Patients with Acute Pancreatitis. Arch. Med. Res. 2018, 49, 504–511. [Google Scholar] [CrossRef]
- Yuan, H.; Jin, X.; Sun, J.; Li, F.; Feng, Q.; Zhang, C.; Cao, Y.; Wang, Y. Protective Effect of HMGB1 A Box on Organ Injury of Acute Pancreatitis in Mice. Pancreas 2009, 38, 143–148. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.; Dui, D.; Ren, H.; Liu, J. High mobility group box 1 induces the activation of the Janus kinase 2 and signal transducer and activator of transcription 3 (JAK2/STAT3) signaling pathway in pancreatic acinar cells in rats, while AG490 and rapamycin inhibit their activation. Bosn. J. Basic Med. Sci. 2016, 16, 307–312. [Google Scholar] [CrossRef] [PubMed]
- Gao, E.; Jiang, Y.; Li, Z.; Xue, D.; Zhang, W. Association between high mobility group box-1 protein expression and cell death in acute pancreatitis. Mol. Med. Rep. 2017, 15, 4021–4026. [Google Scholar] [CrossRef] [PubMed]
- Irie, Y.; Tsubota, M.; Ishikura, H.; Sekiguchi, F.; Terada, Y.; Tsujiuchi, T.; Liu, K.; Nishibori, M.; Kawabata, A. Macrophage-derived HMGB1 as a Pain Mediator in the Early Stage of Acute Pancreatitis in Mice: Targeting RAGE and CXCL12/CXCR4 Axis. J. Neuroimmune Pharmacol. 2017, 12, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Yang, Z.; Wang, H.; Zhao, Y.; Gao, X.; Zang, B. High-mobility group box protein-1 induces acute pancreatitis through ac-tivation of neutrophil extracellular trap and subsequent production of IL-1β. Life Sci. 2021, 286, 119231. [Google Scholar] [CrossRef]
- Luan, Z.-G.; Zhang, X.-J.; Yin, X.-H.; Ma, X.-C.; Zhang, H.; Zhang, C.; Guo, R.-X. Downregulation of HMGB1 protects against the development of acute lung injury after severe acute pancreatitis. Immunobiology 2013, 218, 1261–1270. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, H.-X.; Bai, C.; Zhou, X.-Y. Blockade of high-mobility group box 1 attenuates intestinal mucosal barrier dysfunction in experimental acute pancreatitis. Sci. Rep. 2017, 7, 6799. [Google Scholar] [CrossRef]
- Huang, L.; Zhang, D.; Han, W.; Guo, C. High-mobility group box-1 inhibition stabilizes intestinal permeability through tight junctions in experimental acute necrotizing pancreatitis. Agents Actions 2019, 68, 677–689. [Google Scholar] [CrossRef]
- Kang, R.; Zhang, Q.; Hou, W.; Yan, Z.; Chen, R.; Bonaroti, J.; Bansal, P.; Billiar, T.R.; Tsung, A.; Wang, Q.; et al. Intracellular Hmgb1 Inhibits Inflammatory Nucleosome Release and Limits Acute Pancreatitis in Mice. Gastroenterology 2014, 146, 1097–1107.e8. [Google Scholar] [CrossRef]
- Zhu, C.-J.; Yang, W.-G.; Li, D.-J.; Song, Y.-D.; Chen, S.-Y.; Wang, Q.-F.; Liu, Y.-N.; Zhang, Y.; Cheng, B.; Wu, Z.-W.; et al. Calycosin attenuates severe acute pancreatitis-associated acute lung injury by curtailing high mobility group box 1—Induced inflammation. World J. Gastroenterol. 2021, 27, 7669–7686. [Google Scholar] [CrossRef]
- Wen, Y.; Sun, H.Y.; Tan, Z.; Liu, R.H.; Huang, S.Q.; Chen, G.Y.; Qi, H.; Tang, L.J. Abdominal paracentesis drainage ameliorates myocardial injury in severe experimental pan-creatitis rats through suppressing oxidative stress. World J. Gastroenterol. 2020, 26, 35–54. [Google Scholar] [CrossRef]
- Liu, S.; Stolz, D.B.; Sappington, P.L.; Macias, C.A.; Killeen, M.E.; Tenhunen, J.J.; Delude, R.L.; Fink, M.P. HMGB1 is secreted by immunostimulated enterocytes and contributes to cytomix-induced hyperpermeability of Caco-2 monolayers. Am. J. Physiol. Physiol. 2006, 290, C990–C999. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Sun, H.; Bai, Y.; Zhi, F. Gut dysbiosis-derived exosomes trigger hepatic steatosis by transiting HMGB1 from intestinal to liver in mice. Biochem. Biophys. Res. Commun. 2019, 509, 767–772. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Deng, M.; Loughran, P.A.; Yang, M.; Lin, M.; Yang, C.; Gao, W.; Jin, S.; Li, S.; Scott, M.J.; et al. LPS Induces Active HMGB1 Release From Hepatocytes Into Exosomes Through the Co-ordinated Activities of TLR4 and Caspase-11/GSDMD Signaling. Front. Immunol. 2020, 11, 229. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Ali, S.A. Multifunctional Role of S100 Protein Family in the Immune System: An Update. Cells 2022, 11, 2274. [Google Scholar] [CrossRef]
- Kerkhoff, C.; Klempt, M.; Sorg, C. Novel insights into structure and function of MRP8 (S100A8) and MRP14 (S100A9). Biochim. Biophys. Acta 1998, 1448, 200–211. [Google Scholar] [CrossRef]
- Schiopu, A.; Cotoi, O.S. S100A8 and S100A9: DAMPs at the Crossroads between Innate Immunity, Traditional Risk Factors, and Cardiovascular Disease. Mediat. Inflamm. 2013, 2013, 828354. [Google Scholar] [CrossRef]
- Norman, G.L.; Navaz, S.A.; Kanthi, Y.; Albesa, R.; Mahler, M.; Knight, J.S.; Zuo, Y. Circulating Calprotectin as a Predictive and Severity Biomarker in Patients with COVID-19. Diagnostics 2022, 12, 1324. [Google Scholar] [CrossRef]
- Mortensen, J.H.; Sinkeviciute, D.; Manon-Jensen, T.; Domislović, V.; McCall, K.; Thudium, C.S.; Brinar, M.; Önnerfjord, P.; Goodyear, C.S.; Krznarić, Ž.; et al. A specific calprotectin neo-epitope (CPa9-HNE) in serum from in-flammatory bowel disease patients is associated with neutrophil activity and endoscopic severity. J. Crohns Colitis 2022, 16, 1447–1460. [Google Scholar] [CrossRef]
- Liao, W.-C.; Huang, B.-S.; Yu, Y.-H.; Yang, H.-H.; Chen, P.-R.; Huang, C.-C.; Huang, H.-Y.; Wu, M.-S.; Chow, L.-P. Galectin-3 and S100A9: Novel Diabetogenic Factors Mediating Pancreatic Cancer–Associated Diabetes. Diabetes Care 2019, 42, 1752–1759. [Google Scholar] [CrossRef]
- Farkas, G., Jr.; Tiszlavicz, Z.; Takács, T.; Szabolcs, A.; Farkas, G.; Somogyvári, F.; Mándi, Y. Analysis of plasma levels and polymorphisms of S100A8/9 and S100A12 in patients with acute pancreatitis. Pancreas 2014, 43, 485–487. [Google Scholar] [CrossRef]
- Waldron, R.T.; Lugea, A.; Gulla, A.; Pandol, S.J. Proteomic Identification of Novel Plasma Biomarkers and Pathobiologic Pathways in Alcoholic Acute Pancreatitis. Front. Physiol. 2018, 9, 1215. [Google Scholar] [CrossRef] [PubMed]
- Carrascal, M.; Areny-Balagueró, A.; De-Madaria, E.; Cárdenas-Jaén, K.; García-Rayado, G.; Rivera, R.; Mateos, R.M.M.; Pascual-Moreno, I.; Gironella, M.; Abian, J.; et al. Inflammatory capacity of exosomes released in the early stages of acute pancreatitis predicts the severity of the disease. J. Pathol. 2021, 256, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Nesvaderani, M.; Dhillon, B.K.; Chew, T.; Tang, B.; Baghela, A.; Hancock, R.; Eslick, G.; Cox, M. Gene Expression Profiling: Identification of Novel Pathways and Potential Bi-omarkers in Severe Acute Pancreatitis. J. Am. Coll. Surg. 2022, 234, 803–815. [Google Scholar] [CrossRef]
- Sprenkeler, E.G.G.; Zandstra, J.; van Kleef, N.D.; Goetschalckx, I.; Verstegen, B.; Aarts, C.E.M.; Janssen, H.; Tool, A.T.J.; van Mierlo, G.; van Bruggen, R.; et al. S100A8/A9 Is a Marker for the Release of Neutrophil Extracellular Traps and Induces Neutrophil Activation. Cells 2022, 11, 236. [Google Scholar] [CrossRef]
- Schnekenburger, J.; Schick, V.; Krüger, B.; Manitz, M.P.; Sorg, C.; Nacken, W.; Kerkhoff, C.; Kahlert, A.; Mayerle, J.; Domschke, W.; et al. The calcium binding protein S100A9 is essential for pancreatic leukocyte infil-tration and induces disruption of cell-cell contacts. J. Cell Physiol. 2008, 216, 558–567. [Google Scholar] [CrossRef]
- Xiang, H.; Guo, F.; Tao, X.; Zhou, Q.; Xia, S.; Deng, D.; Li, L.; Shang, D. Pancreatic ductal deletion of S100A9 alleviates acute pancreatitis by targeting VNN1-mediated ROS release to inhibit NLRP3 activation. Theranostics 2021, 11, 4467–4482. [Google Scholar] [CrossRef]
- Ding, Z.; Du, F.; Averitt, V.R.G.; Jakobsson, G.; Rönnow, C.-F.; Rahman, M.; Schiopu, A.; Thorlacius, H. Targeting S100A9 Reduces Neutrophil Recruitment, Inflammation and Lung Damage in Abdominal Sepsis. Int. J. Mol. Sci. 2021, 22, 12923. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Lu, R.; Chen, J.; Xie, M.; Zhao, X.; Kong, L. S100A9 blockade prevents lipopolysaccharide-induced lung injury via sup-pressing the NLRP3 pathway. Respir. Res. 2021, 22, 45. [Google Scholar] [CrossRef] [PubMed]
- Al-Kuraishy, H.M.; Al-Gareeb, A.I.; Al-Niemi, M.S.; Alexiou, A.; Batiha, G.E.-S. Calprotectin: The link between acute lung injury and gastrointestinal injury in Covid-19: Ban or boon. Curr. Protein Pept. Sci. 2022, 23, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Aziz, M.; Wang, P. Damage-Associated Molecular Patterns As Double-Edged Swords in Sepsis. Antioxid. Redox Signal. 2021, 35, 1308–1323. [Google Scholar] [CrossRef]
- Aziz, M.; Brenner, M.; Wang, P. Extracellular CIRP (eCIRP) and inflammation. J. Leukoc. Biol. 2019, 106, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Tong, L.; Tang, L.; Wu, S. The role of cold-inducible RNA binding protein in cell stress response. Int. J. Cancer 2017, 141, 2164–2173. [Google Scholar] [CrossRef] [PubMed]
- Zhong, P.; Peng, J.; Yuan, M.; Kong, B.; Huang, H. Cold-inducible RNA-binding protein (CIRP) in inflammatory diseases: Molecular insights of its associated signalling pathways. Scand. J. Immunol. 2020, 93, e12949. [Google Scholar] [CrossRef]
- Gong, J.-D.; Qi, X.-F.; Zhang, Y.; Li, H.-L. Increased admission serum cold-inducible RNA-binding protein concentration is associated with prognosis of severe acute pancreatitis. Clin. Chim. Acta 2017, 471, 135–142. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, Y.; Li, C.; Xu, Y.; Wang, X.; Wu, D.; Gao, Z.; Qian, H.; You, Z.; Zhang, Z.; et al. Extracellular CIRP-Impaired Rab26 Restrains EPOR-Mediated Macrophage Polarization in Acute Lung Injury. Front. Immunol. 2021, 12, 768435. [Google Scholar] [CrossRef] [PubMed]
- Herre, M.; Cedervall, J.; Mackman, N.; Olsson, A.K. Neutrophil extracellular traps in the pathology of cancer and other in-flammatory diseases. Physiol. Rev. 2022, 103, 277–312. [Google Scholar] [CrossRef]
- Scozzi, D.; Liao, F.; Krupnick, A.S.; Kreisel, D.; Gelman, A.E. The role of neutrophil extracellular traps in acute lung injury. Front. Immunol. 2022, 13, 953195. [Google Scholar] [CrossRef]
- Wan, J.; Ren, Y.; Yang, X.; Li, X.; Xia, L.; Lu, N. The Role of Neutrophils and Neutrophil Extracellular Traps in Acute Pancreatitis. Front. Cell Dev. Biol. 2021, 8, 565758. [Google Scholar] [CrossRef]
- Linders, J.; Madhi, R.; Rahman, M.; Mörgelin, M.; Regner, S.; Brenner, M.; Wang, P.; Thorlacius, H. Extracellular cold-inducible RNA-binding protein regulates neutrophil extracellular trap formation and tissue damage in acute pancreatitis. Lab. Investig. 2020, 100, 1618–1630. [Google Scholar] [CrossRef]
- Tan, C.; Gurien, S.D.; Royster, W.; Aziz, M.; Wang, P. Extracellular CIRP Induces Inflammation in Alveolar Type II Cells via TREM-1. Front. Cell Dev. Biol. 2020, 8, 579157. [Google Scholar] [CrossRef]
- Murao, A.; Tan, C.; Jha, A.; Wang, P.; Aziz, M. Exosome-Mediated eCIRP Release From Macrophages to Induce Inflammation in Sepsis. Front. Pharmacol. 2021, 12, 791648. [Google Scholar] [CrossRef] [PubMed]
- Szatmary, P.; Huang, W.; Criddle, D.; Tepikin, A.; Sutton, R. Biology, role and therapeutic potential of circulating histones in acute in-flammatory disorders. J. Cell Mol. Med. 2018, 22, 4617–4629. [Google Scholar] [CrossRef] [PubMed]
- Ligi, D.; Giglio, R.V.; Henry, B.M.; Lippi, G.; Ciaccio, M.; Plebani, M.; Mannello, F. What is the impact of circulating histones in COVID-19: A systematic review. Clin. Chem. Lab. Med. 2022, 60, 1506–1517. [Google Scholar] [CrossRef]
- Li, Y.; Wan, D.; Luo, X.; Song, T.; Wang, Y.; Yu, Q.; Jiang, L.; Liao, R.; Zhao, W.; Su, B. Circulating Histones in Sepsis: Potential Outcome Predictors and Therapeutic Targets. Front. Immunol. 2021, 12, 650184. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.R.; Mazza, D.; Brambilla, F.; Gorzanelli, A.; Agresti, A.; Bianchi, M.E. LPS-Challenged Macrophages Release Microvesicles Coated With Histones. Front. Immunol. 2018, 9, 1463. [Google Scholar] [CrossRef] [PubMed]
- Keerthikumar, S.; Chisanga, D.; Ariyaratne, D.; Al Saffar, H.; Anand, S.; Zhao, K.; Samuel, M.; Pathan, M.; Jois, M.; Chilamkurti, N.; et al. ExoCarta: A Web-Based Compendium of Exosomal Cargo. J. Mol. Biol. 2016, 428, 688–692. [Google Scholar] [CrossRef]
- Pathan, M.; Fonseka, P.; Chitti, S.V.; Kang, T.; Sanwlani, R.; Van Deun, J.; Hendrix, A.; Mathivanan, S. Vesiclepedia 2019: A compendium of RNA, proteins, lipids and metabolites in extracellular vesicles. Nucleic Acids Res. 2018, 47, D516–D519. [Google Scholar] [CrossRef]
- Pan, B.; Li, Y.; Liu, Y.; Wang, W.; Huang, G.; Ouyang, Y. Circulating CitH3 Is a Reliable Diagnostic and Prognostic Biomarker of Septic Patients in Acute Pancreatitis. Front. Immunol. 2021, 12, 766391. [Google Scholar] [CrossRef]
- Penttilä, A.K.; Rouhiainen, A.; Kylänpää, L.; Mustonen, H.; Puolakkainen, P.; Rauvala, H.; Repo, H. Circulating nucleosomes as predictive markers of severe acute pancreatitis. J. Intensive Care 2016, 4, 14. [Google Scholar] [CrossRef]
- Pérez, S.; Finamor, I.; Martí-Andrés, P.; Pereda, J.; Campos, A.; Domingues, R.; Haj, F.; Sabater, L.; De-Madaria, E.; Sastre, J. Role of obesity in the release of extracellular nucleosomes in acute pancreatitis: A clinical and experimental study. Int. J. Obes. 2018, 43, 158–168. [Google Scholar] [CrossRef]
- Liu, T.; Huang, W.; Szatmary, P.; Abrams, S.T.; Alhamdi, Y.; Lin, Z.; Greenhalf, W.; Wang, G.; Sutton, R.; Toh, C.H. Accuracy of circulating histones in predicting persistent organ failure and mortality in patients with acute pancreatitis. Br. J. Surg. 2017, 104, 1215–1225. [Google Scholar] [CrossRef] [PubMed]
- Biberci Keskin, E.; İnce, A.T.; Sümbül Gültepe, B.; Köker, İ.H.; Şentürk, H. The relationship between serum histon levels and the severity of acute pancreatitis. Turk. J. Gastroenterol. 2019, 30, 807–810. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.; Abrams, S.T.; Alhamdi, Y.; Toh, J.; Yu, W.; Wang, G.; Toh, C.-H. Circulating Histones Are Major Mediators of Multiple Organ Dysfunction Syndrome in Acute Critical Illnesses. Crit. Care Med. 2019, 47, e677–e684. [Google Scholar] [CrossRef]
- Guo, H.Y.; Cui, Z.J. Extracellular Histones Activate Plasma Membrane Toll-Like Receptor 9 to Trigger Calcium Oscillations in Rat Pancreatic Acinar Tumor Cell AR4-2J. Cells 2018, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Cao, F.; Ding, Y.; Lu, J.; Liu, S.; Li, F. Acinar Cells Derived Exosomes Alleviate the Severity of Acute Pancreatitis. Discov. Med. 2021, 31, 95–105. [Google Scholar] [PubMed]
- Zhou, Q.; Xiang, H.; Liu, H.; Qi, B.; Shi, X.; Guo, W.; Zou, J.; Wan, X.; Wu, W.; Wang, Z.; et al. Emodin Alleviates Intestinal Barrier Dysfunction by Inhibiting Apoptosis and Regulating the Immune Response in Severe Acute Pancreatitis. Pancreas 2021, 50, 1202–1211. [Google Scholar] [CrossRef] [PubMed]
- Hu, Q.; Yao, J.; Wu, X.; Li, J.; Li, G.; Tang, W.; Liu, J.; Wan, M. Emodin attenuates severe acute pancreatitis-associated acute lung injury by suppressing pancreatic exosome-mediated alveolar macrophage activation. Acta Pharm. Sin. B 2021, 12, 3986–4003. [Google Scholar] [CrossRef]
- Feng, Z.; Zhou, J.; Liu, Y.; Xia, R.; Li, Q.; Yan, L.; Chen, Q.; Chen, X.; Jiang, Y.; Chao, G.; et al. Epithelium- and endothelium-derived exosomes regulate the alveolar macrophages by targeting RGS1 mediated calcium signaling-dependent immune response. Cell Death Differ. 2021, 28, 2238–2256. [Google Scholar] [CrossRef]
- Zheng, L.; Su, J.; Zhang, Z.; Jiang, L.; Wei, J.; Xu, X.; Lv, S. Salidroside regulates inflammatory pathway of alveolar macrophages by influencing the se-cretion of miRNA-146a exosomes by lung epithelial cells. Sci. Rep. 2020, 10, 20750. [Google Scholar] [CrossRef]
- Ye, C.; Li, H.; Bao, M.; Zhuo, R.; Jiang, G.; Wang, W. Alveolar macrophage—Derived exosomes modulate severity and outcome of acute lung injury. Aging 2020, 12, 6120–6128. [Google Scholar] [CrossRef]
- Qin, X.; Zhou, Y.; Jia, C.; Chao, Z.; Qin, H.; Liang, J.; Liu, X.; Liu, Z.; Sun, T.; Yuan, Y.; et al. Caspase-1-mediated extracellular vesicles derived from pyroptotic alveolar macrophages promote inflammation in acute lung injury. Int. J. Biol. Sci. 2022, 18, 1521–1538. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhu, L.; Li, H.; Ren, W.; Zhuo, R.; Feng, C.; He, Y.; Hu, Y.; Ye, C. Alveolar macrophage-derived exosomal tRF-22-8BWS7K092 activates Hippo signaling pathway to induce ferroptosis in acute lung injury. Int. Immunopharmacol. 2022, 107, 108690. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Cao, X.; Qin, L. Mesenchymal Stem Cell-Derived Extracellular Vesicles: A Novel Cell-Free Therapy for Sepsis. Front. Immunol. 2020, 11, 647. [Google Scholar] [CrossRef] [PubMed]
- Goodman, R.R.; Jong, M.K.; Davies, J.E. Concise review: The challenges and opportunities of employing mesenchymal stromal cells in the treatment of acute pancreatitis. Biotechnol. Adv. 2019, 42, 107338. [Google Scholar] [CrossRef]
- Pu, Q.; Xiu, G.; Sun, J.; Liu, P.; Ling, B. Progress on the effect of mesenchymal stem cell derived exosomes on multiple organ dysfunction in sepsis. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2021, 33, 757–760. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Li, X.; Su, X.; Xiao, H.; Yang, J. Hypoxic Preconditioning of Human Umbilical Cord Mesenchymal Stem Cells Is an Effective Strategy for Treating Acute Lung Injury. Stem Cells Dev. 2021, 30, 128–134. [Google Scholar] [CrossRef]
- Shaikh, M.S.; Shahzad, Z.; Tash, E.A.; Janjua, O.S.; Khan, M.I.; Zafar, M.S. Human Umbilical Cord Mesenchymal Stem Cells: Current Literature and Role in Periodontal Regeneration. Cells 2022, 11, 1168. [Google Scholar] [CrossRef]
- Han, L.; Zhao, Z.; Chen, X.; Yang, K.; Tan, Z.; Huang, Z.; Zhou, L.; Dai, R. Human umbilical cord mesenchymal stem cells-derived exosomes for treating traumatic pan-creatitis in rats. Stem Cell Res. Ther. 2022, 13, 221. [Google Scholar] [CrossRef]
- Luo, Y.; Li, Z.; Ge, P.; Guo, H.; Li, L.; Zhang, G.; Xu, C.; Chen, H. Comprehensive Mechanism, Novel Markers and Multidisciplinary Treatment of Severe Acute Pan-creatitis-Associated Cardiac Injury—A Narrative Review. J. Inflamm. Res. 2021, 14, 3145–3169. [Google Scholar] [CrossRef]
- Chen, M.; Chen, J.; Huang, W.; Li, C.; Luo, H.; Xue, Z.; Xiao, Y.; Wu, Q.; Chen, C. Exosomes from human induced pluripotent stem cells derived mesenchymal stem cells improved myocardial injury caused by severe acute pancreatitis through activating Akt/Nrf2/HO-1 axis. Cell Cycle 2022, 21, 1578–1589. [Google Scholar] [CrossRef]
- Xia, L.; Zhang, C.; Lv, N.; Liang, Z.; Ma, T.; Cheng, H.; Xia, Y.; Shi, L. AdMSC-derived exosomes alleviate acute lung injury via transferring mitochondrial component to improve homeostasis of alveolar macrophages. Theranostics 2022, 12, 2928–2947. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Q.; Kim, K.-H. Emergence of Edible Plant-Derived Nanovesicles as Functional Food Components and Nanocarriers for Therapeutics Delivery: Potentials in Human Health and Disease. Cells 2022, 11, 2232. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Zhang, L.; Zhang, Y.; Lu, R. Plant-Derived Exosomes as a Drug-Delivery Approach for the Treatment of Inflammatory Bowel Disease and Colitis-Associated Cancer. Pharmaceutics 2022, 14, 822. [Google Scholar] [CrossRef]
- Zhang, Z.; Yu, Y.; Zhu, G.; Zeng, L.; Xu, S.; Cheng, H.; Ouyang, Z.; Chen, J.; Pathak, J.L.; Wu, L.; et al. The Emerging Role of Plant-Derived Exosomes-Like Nanoparticles in Immune Regulation and Periodontitis Treatment. Front. Immunol. 2022, 13, 896745. [Google Scholar] [CrossRef]
- Nemati, M.; Singh, B.; Mir, R.A.; Nemati, M.; Babaei, A.; Ahmadi, M.; Rasmi, Y.; Golezani, A.G.; Rezaie, J. Plant-derived extracellular vesicles: A novel nanomedicine approach with advantages and challenges. Cell Commun. Signal. 2022, 20, 69. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Xu, F.; Zhang, X.; Mu, J.; Sayed, M.; Hu, X.; Lei, C.; Sriwastva, M.; Kumar, A.; Sundaram, K.; et al. Plant-derived exosomal microRNAs inhibit lung inflammation induced by exosomes SARS-CoV-2 Nsp12. Mol. Ther. 2021, 29, 2424–2440. [Google Scholar] [CrossRef]
- Yang, D.; Zhang, W.; Zhang, H.; Zhang, F.; Chen, L.; Ma, L.; Larcher, L.M.; Chen, S.; Liu, N.; Zhao, Q.; et al. Progress, opportunity, and perspective on exosome isolation—Efforts for efficient exosome-based theranostics. Theranostics 2020, 10, 3684–3707. [Google Scholar] [CrossRef]
- Liu, W.-Z.; Ma, Z.-J.; Kang, X.-W. Current status and outlook of advances in exosome isolation. Anal. Bioanal. Chem. 2022, 414, 7123–7141. [Google Scholar] [CrossRef] [PubMed]
- Desai, C.S.; Khan, A.; Bellio, M.A.; Willis, M.L.; Mahung, C.; Ma, X.; Baldwin, X.; Williams, B.M.; Baron, T.H.; Coleman, L.G.; et al. Characterization of extracellular vesicle miRNA identified in peripheral blood of chronic pancreatitis patients. Mol. Cell. Biochem. 2021, 476, 4331–4341. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wu, J.; Ye, N.; Li, F.; Zhan, H.; Chen, S.; Xu, J. Plasma-Derived Exosome MiR-19b Acts as a Diagnostic Marker for Pancreatic Cancer. Front. Oncol. 2021, 11, 739111. [Google Scholar] [CrossRef]
- Wu, Y.; Zeng, H.; Yu, Q.; Huang, H.; Fervers, B.; Chen, Z.-S.; Lu, L. A Circulating Exosome RNA Signature Is a Potential Diagnostic Marker for Pancreatic Cancer, a Systematic Study. Cancers 2021, 13, 2565. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Zhang, Y.; Gao, X.; Yuan, Y.; Zhao, J.; Zhou, S.; Wang, H.; Wang, L.; Xu, G.; Li, X.; et al. Plasma-Derived Exosomal ALIX as a Novel Biomarker for Diagnosis and Classification of Pancreatic Cancer. Front. Oncol. 2021, 11, 628346. [Google Scholar] [CrossRef] [PubMed]
- Yan, Z.; Zang, B.; Gong, X.; Ren, J.; Wang, R. MiR-214-3p exacerbates kidney damages and inflammation induced by hyperlipidemic pancreatitis complicated with acute renal injury. Life Sci. 2019, 241, 117118. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Liu, S.; Wang, F. MicroRNA MiR-27a-5p Alleviates the Cerulein-Induced Cell Apoptosis and Inflammatory Injury of AR42J Cells by Targeting Traf3 in Acute Pancreatitis. Inflammation 2020, 43, 1988–1998. [Google Scholar] [CrossRef]
- Erdos, Z.; Barnum, J.E.; Wang, E.; DeMaula, C.; Dey, P.M.; Forest, T.; Bailey, W.J.; Glaab, W.E. Evaluation of the Relative Performance of Pancreas-Specific MicroRNAs in Rat Plasma as Biomarkers of Pancreas Injury. Toxicol. Sci. 2019, 173, 5–18. [Google Scholar] [CrossRef]
- Yu, W.; Zhang, M.; Li, X.; Pan, N.; Bian, X.; Wu, W. Protective Effect of miR-193a-5p and miR-320-5p on Caerulein-Induced Injury in AR42J Cells. Am. J. Dig. Dis. 2021, 66, 4333–4343. [Google Scholar] [CrossRef]
- Usborne, A.L.; Smith, A.T.; Engle, S.K.; Watson, D.E.; Sullivan, J.M.; Walgren, J.L. Biomarkers of exocrine pancreatic injury in 2 rat acute pancreatitis models. Toxicol. Pathol. 2014, 42, 195–203. [Google Scholar] [CrossRef]
- Miao, B.; Qi, W.; Zhang, S.; Wang, H.; Wang, C.; Hu, L.; Huang, G.; Li, S.; Wang, H. miR-148a suppresses autophagy by down-regulation of IL-6/STAT3 signaling in ceru-lein-induced acute pancreatitis. Pancreatology 2019, 19, 557–565. [Google Scholar] [CrossRef]
- Song, G.; Zhou, J.; Song, R.; Liu, D.; Yu, W.; Xie, W.; Ma, Z.; Gong, J.; Meng, H.; Yang, T.; et al. Long noncoding RNA H19 regulates the therapeutic efficacy of mesenchymal stem cells in rats with severe acute pancreatitis by sponging miR-138-5p and miR-141-3p. Stem Cell Res. Ther. 2020, 11, 420. [Google Scholar] [CrossRef]
- Liu, P.; Xia, L.; Zhang, W.-L.; Ke, H.-J.; Su, T.; Deng, L.-B.; Chen, Y.-X.; Lv, N.-H. Identification of serum microRNAs as diagnostic and prognostic biomarkers for acute pancreatitis. Pancreatology 2014, 14, 159–166. [Google Scholar] [CrossRef]
- Lu, P.; Wang, F.; Wu, J.; Wang, C.; Yan, J.; Li, Z.-L.; Song, J.-X.; Wang, J.-J. Elevated Serum miR-7, miR-9, miR-122, and miR-141 Are Noninvasive Biomarkers of Acute Pancreatitis. Dis. Markers 2017, 2017, 7293459. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Huang, L.; Zhu, S.; Li, X.; Li, Z.; Yu, C.; Yu, X. Regulation of autophagy by systemic admission of microRNA-141 to target HMGB1 in l-arginine-induced acute pancreatitis in vivo. Pancreatology 2016, 16, 337–346. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yan, L.; Han, W. Elevated Level of miR-551b-5p is Associated With Inflammation and Disease Progression in Patients With Severe Acute Pancreatitis. Ther. Apher. Dial. 2018, 22, 649–655. [Google Scholar] [CrossRef] [PubMed]
- Kuśnierz-Cabala, B.; Nowak, E.; Sporek, M.; Kowalik, A.; Kuźniewski, M.; Enguita, F.J.; Stępień, E. Serum levels of unique miR-551-5p and endothelial-specific miR-126a-5p allow discrimination of patients in the early phase of acute pancreatitis. Pancreatology 2015, 15, 344–351. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Wang, H.; Xue, D.; Zhang, W. Screening and validation of differentially expressed extracellular miRNAs in acute pan-creatitis. Mol. Med. Rep. 2017, 16, 6412–6418. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.G.; Kang, X.; Zhan, L.B.; Kang, L.M.; Fan, Z.W.; Bai, L.Z. Circulating miRNAs as biomarkers for severe acute pancreatitis associated with acute lung injury. World J. Gastroenterol. 2017, 23, 7440–7449. [Google Scholar] [CrossRef]
- Shi, N.; Deng, L.; Chen, W.; Zhang, X.; Luo, R.; Jin, T.; Ma, Y.; Du, C.; Lin, Z.; Jiang, K.; et al. Is MicroRNA-127 a Novel Biomarker for Acute Pancreatitis with Lung Injury? Dis. Markers 2017, 2017, 1204295. [Google Scholar] [CrossRef]
- Shan, Y.; Kong, W.; Zhu, A.; Zhang, J.; Ying, R.; Zhu, W. Increased levels of miR-372 correlate with disease progression in patients with hyper-lipidemic acute pancreatitis. Exp. Ther. Med. 2020, 19, 3845–3850. [Google Scholar]
- Chen, Y.J.; Lin, T.L.; Cai, Z.; Yan, C.H.; Gou, S.R.; Zhuang, Y.D. Assessment of acute pancreatitis severity via determination of serum levels of hsa-miR-126-5p and IL-6. Exp. Ther. Med. 2021, 21, 26. [Google Scholar] [CrossRef]
- Li, X.Y.; Wang, Y.F.; Li, N. Circulating microRNA-146a and microRNA-146b exhibit potential to serve as markers for acute pancreatitis management and prognosis. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 12770–12780. [Google Scholar]
- Dai, J.; Jiang, M.; Hu, Y.; Xiao, J.; Hu, B.; Xu, J.; Han, X.; Shen, S.; Li, B.; Wu, Z.; et al. Dysregulated SREBP1c/miR-153 signaling induced by hypertriglyceridemia worsens acute pancreatitis and delays tissue repair. JCI Insight 2021, 6, e138584. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Chen, W.; Lu, J.; Zhang, S.; Xiang, X.; Wang, X.; Tang, G. Circ_0000284 Promoted Acute Pancreatitis Progression through the Regulation of miR-10a-5p/Wnt/β-Catenin Pathway. Chem. Biodivers. 2022, 19, e202101006. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.; Pan, L.; Yang, L.; Niu, Z.; Wang, L.; Gao, Y.; Liu, J.; Liu, Z.; Pei, H. Interfering hsa_circ_0073748 alleviates caerulein-induced ductal cell injury in acute pancreatitis by inhibiting miR-132-3p/TRAF3/NF-κB pathway. Cell Cycle 2022, 21, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.; Chen, Y.; Sun, W.W.; Chen, W.W.; Ma, L.; Yang, Z.T.; Huang, J.; Chen, E.Z.; Fei, J.; Mao, E.Q. Effect of S100A12 and soluble receptor for advanced glycation end products on the occurrence of severe acute pancreatitis. J. Dig. Dis. 2016, 17, 475–482. [Google Scholar] [CrossRef]
- Wu, L.; Xu, W.; Wang, F.; Lv, T.; Yin, Z.; Song, Y. Plasma mtDNA Analysis Aids in Predicting Pancreatic Necrosis in Acute Pancreatitis Patients: A Pilot Study. Am. J. Dig. Dis. 2018, 63, 2975–2982. [Google Scholar] [CrossRef]
- Li, N.; Wang, B.; Cai, S.; Liu, P. The Role of Serum High Mobility Group Box 1 and Interleukin-6 Levels in Acute Pancreatitis: A Meta-Analysis. J. Cell. Biochem. 2017, 119, 616–624. [Google Scholar] [CrossRef]
- Owusu, L.; Xu, C.; Chen, H.; Liu, G.; Zhang, G.; Zhang, J.; Tang, Z.; Sun, Z.; Yi, X. Gamma-enolase predicts lung damage in severe acute pancreatitis-induced acute lung injury. Histochem. J. 2018, 49, 347–356. [Google Scholar] [CrossRef]
- Moutinho-Ribeiro, P.; Macedo, G.; Melo, S.A. Pancreatic Cancer Diagnosis and Management: Has the Time Come to Prick the Bubble? Front. Endocrinol. 2019, 9, 779. [Google Scholar] [CrossRef]
- Yoshizawa, N.; Sugimoto, K.; Tameda, M.; Inagaki, Y.; Ikejiri, M.; Inoue, H.; Usui, M.; Ito, M.; Takei, Y. miR-3940-5p/miR-8069 ratio in urine exosomes is a novel diagnostic biomarker for pancreatic ductal adenocarcinoma. Oncol. Lett. 2020, 19, 2677–2684. [Google Scholar] [CrossRef]
- Yang, J.; Xu, R.; Wang, C.; Qiu, J.; Ren, B.; You, L. Early screening and diagnosis strategies of pancreatic cancer: A comprehensive review. Cancer Commun. 2021, 41, 1257–1274. [Google Scholar] [CrossRef]
- Villaseñor, A.; Kinross, J.M.; Li, J.V.; Penney, N.; Barton, R.H.; Nicholson, J.K.; Darzi, A.; Barbas, C.; Holmes, E. 1H NMR Global Metabolic Phenotyping of Acute Pancreatitis in the Emergency Unit. J. Proteome Res. 2014, 13, 5362–5375. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-T.; Liao, H.-Y.; Huang, W.-H.; Lin, S.-Y.; Tsai, T.-Y.; Yang, C.-Y.; Tsai, F.-J.; Chen, C.-J. Early prediction of severe acute pancreatitis by urinary β-2 microglobulin/saposin B peak ratios on MALDI-TOF. Clin. Chim. Acta 2015, 440, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Pallagi, P.; Hegyi, P.; Rakonczay, Z., Jr. The Physiology and Pathophysiology of Pancreatic Ductal Secretion: The Back-ground for Clinicians. Pancreas 2015, 44, 1211–1233. [Google Scholar] [CrossRef] [PubMed]
- Osteikoetxea, X.; Benke, M.; Rodriguez, M.; Pálóczi, K.; Sódar, B.W.; Szvicsek, Z.; Szabó-Taylor, K.; Vukman, K.V.; Kittel, Á.; Wiener, Z.; et al. Detection and proteomic characterization of extracellular vesicles in human pancreatic juice. Biochem. Biophys. Res. Commun. 2018, 499, 37–43. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Sadakari, Y.; Ohtsuka, T.; Okayama, T.; Nakashima, Y.; Gotoh, Y.; Saeki, K.; Mori, Y.; Nakata, K.; Miyasaka, Y.; et al. Pancreatic Juice Exosomal MicroRNAs as Biomarkers for Detection of Pancreatic Ductal Adenocarcinoma. Ann. Surg. Oncol. 2019, 26, 2104–2111. [Google Scholar] [CrossRef]
- Ge, P.; Zhang, J.; Zhang, G.; Chen, H. Research progress on application of metabolomics in acute lung injury. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2021, 33, 1266–1271. [Google Scholar]
- Papadopoulos, S.; Kazepidou, E.; Antonelou, M.H.; Leondaritis, G.; Tsapinou, A.; Koulouras, V.P.; Avgeropoulos, A.; Nakos, G.; Lekka, M.E. Secretory Phospholipase A(2)-IIA Protein and mRNA Pools in Extra-cellular Vesicles of Bronchoalveolar Lavage Fluid from Patients with Early Acute Respiratory Distress Syndrome: A New Perception in the Dissemination of Inflammation. Pharmaceuticals 2020, 13, 415. [Google Scholar] [CrossRef]
- Kaur, G.; Maremanda, K.P.; Campos, M.; Chand, H.S.; Li, F.; Hirani, N.; Haseeb, M.A.; Li, D.; Rahman, I. Distinct Exosomal miRNA Profiles from BALF and Lung Tissue of COPD and IPF Patients. Int. J. Mol. Sci. 2021, 22, 11830. [Google Scholar] [CrossRef]
- Kishore, A.; Navratilova, Z.; Kolek, V.; Novosadova, E.; Čépe, K.; Du Bois, R.M.; Petrek, M. Expression analysis of extracellular microRNA in bronchoalveolar lavage fluid from patients with pulmonary sarcoidosis. Respirology 2018, 23, 1166–1172. [Google Scholar] [CrossRef]
- Domagala-Kulawik, J. The relevance of bronchoalveolar lavage fluid analysis for lung cancer patients. Expert Rev. Respir. Med. 2019, 14, 329–337. [Google Scholar] [CrossRef]
- Zhou, B.; Guo, M.; Hao, X.; Lou, B.; Liu, J.; She, J. Altered exosomal microRNA profiles in bronchoalveolar lavage fluid can mediate metabolism in patients with Acinetobacter baumannii ventilator-associated pneumonia. Ann. Transl. Med. 2020, 8, 1561. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Li, Y.-P.; Geng, X.-R.; Zhao, M.; Ma, S.-B.; Yang, Y.-H.; Deng, Z.-H.; Luo, L.-M.; Pan, X.-Q. Expression Level of MiRNA-126 in Serum Exosomes of Allergic Asthma Patients and Lung Tissues of Asthmatic Mice. Curr. Drug Metab. 2019, 20, 799–803. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NcRNAs | Species | Targets | Expression and Role in AP | Expression and Role in ALI | Reference |
|---|---|---|---|---|---|
| MALAT1 | Human, mouse, and rat | miR-194, miR-181a-5p | Up, Aggravated | Up, Aggravated | [43,44,45,47] |
| TUG1 | Human, mouse, and rat | miR-494, miR-34b-5p, miR-9-5p | Up, Aggravated | Down, Suppressive | [49,50,51,52] |
| MiR-155 | Human, mouse, and rat | Rictor, TAB2, SOCS1, RhoA, IL17RB, IL18R1, IL22RA2 | Up/Down, Aggravated | Up, Aggravated | [70,71,72,73,74,75,76,77,78,79,80] |
| miR-21-3p | Human, mouse, and rat | TRPs, | Up/Down, Aggravated | Up, Aggravated | [81,72,83,84,85] |
| miR-21-5p | Human, mouse, and rat | Trim33, SKP2, PGAM5 | Up/Down, Aggravated | Up, Suppressive | [72,82,86,87] |
| miR-216a | Human, mouse, and rat | JAK2 | Up, Aggravated | Up, Aggravated/Suppressive | [22,92,96] |
| Proteins | Species | Expression and Role in AP | Expression and Role in ALI | Reference |
|---|---|---|---|---|
| HMGB1 | Human, mouse, and rat | Up, Aggravated | Up, Aggravated | [109,112,113,114,115,119,120] |
| S100A8/A9 | Human, mouse, and rat | Up, Aggravated | Up, Aggravated | [131,132,133,134,135,136,137,138,139,140] |
| CIRP | Human, mouse, and rat | Up, Aggravated | Up, Aggravated | [23,145,146,150,151,152] |
| Histones | Human, mouse, and rat | Up/No statistical difference, Aggravated | Up, Aggravated | [159,160,161,162,163,164,165] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Q.; Luo, Y.; Lan, B.; Dong, X.; Wang, Z.; Ge, P.; Zhang, G.; Chen, H. Fighting Fire with Fire: Exosomes and Acute Pancreatitis-Associated Acute Lung Injury. Bioengineering 2022, 9, 615. https://doi.org/10.3390/bioengineering9110615
Yang Q, Luo Y, Lan B, Dong X, Wang Z, Ge P, Zhang G, Chen H. Fighting Fire with Fire: Exosomes and Acute Pancreatitis-Associated Acute Lung Injury. Bioengineering. 2022; 9(11):615. https://doi.org/10.3390/bioengineering9110615
Chicago/Turabian StyleYang, Qi, Yalan Luo, Bowen Lan, Xuanchi Dong, Zhengjian Wang, Peng Ge, Guixin Zhang, and Hailong Chen. 2022. "Fighting Fire with Fire: Exosomes and Acute Pancreatitis-Associated Acute Lung Injury" Bioengineering 9, no. 11: 615. https://doi.org/10.3390/bioengineering9110615
APA StyleYang, Q., Luo, Y., Lan, B., Dong, X., Wang, Z., Ge, P., Zhang, G., & Chen, H. (2022). Fighting Fire with Fire: Exosomes and Acute Pancreatitis-Associated Acute Lung Injury. Bioengineering, 9(11), 615. https://doi.org/10.3390/bioengineering9110615