
Continuous Supply of Non-Combustible Gas Mixture for Safe Autotrophic Culture to Produce Polyhydroxyalkanoate by Hydrogen-Oxidizing Bacteria


Abstract
1. Introduction
2. Materials and Methods
2.1. Strains and Plasmids
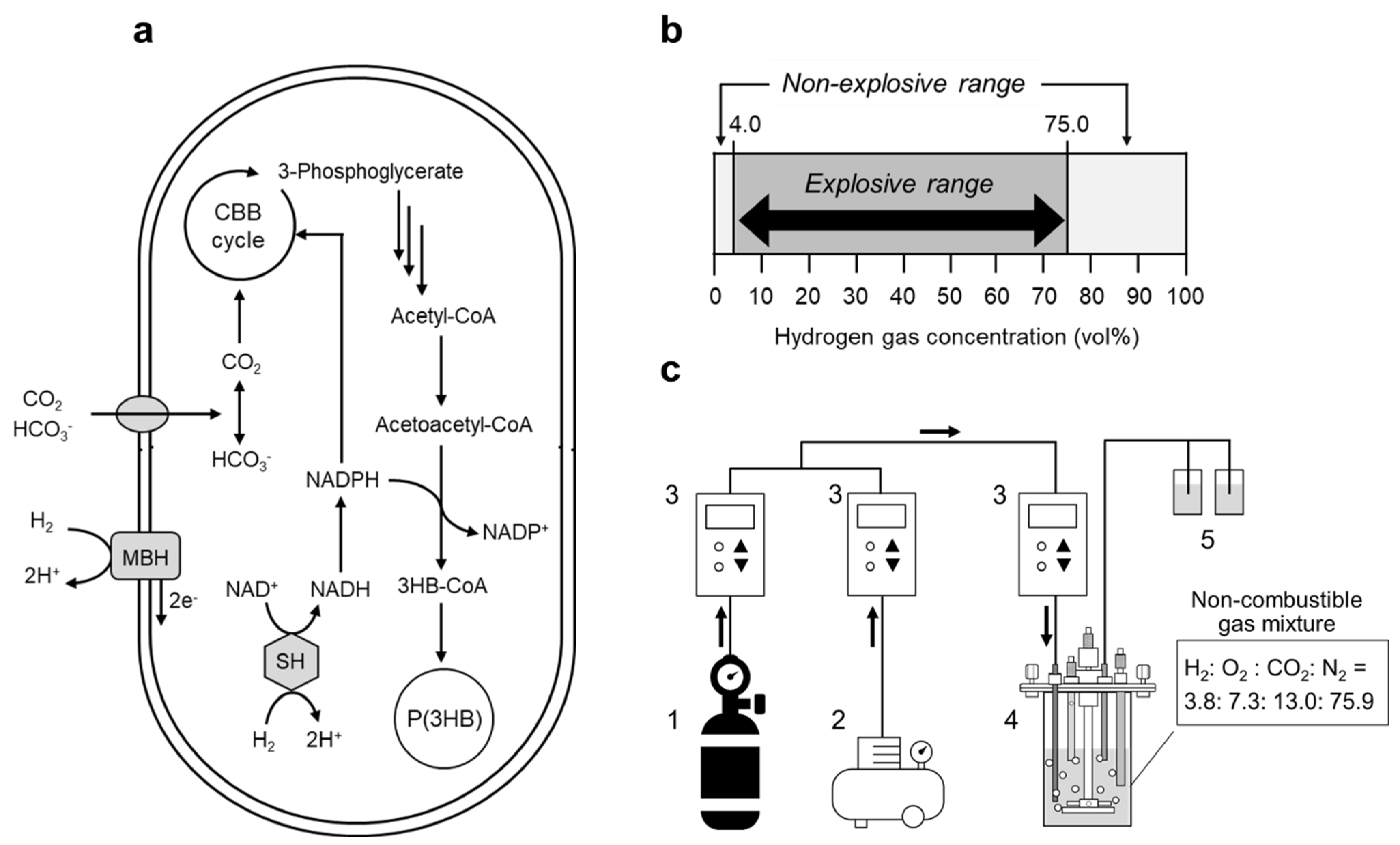
2.2. Autotrophic Cultivation and P(3HB) Production
2.3. Fluorescence Microscopy Analysis
2.4. Quantification of Intracellular P(3HB)
2.5. Extraction of Polymer and Molecular Weight Analysis
3. Results
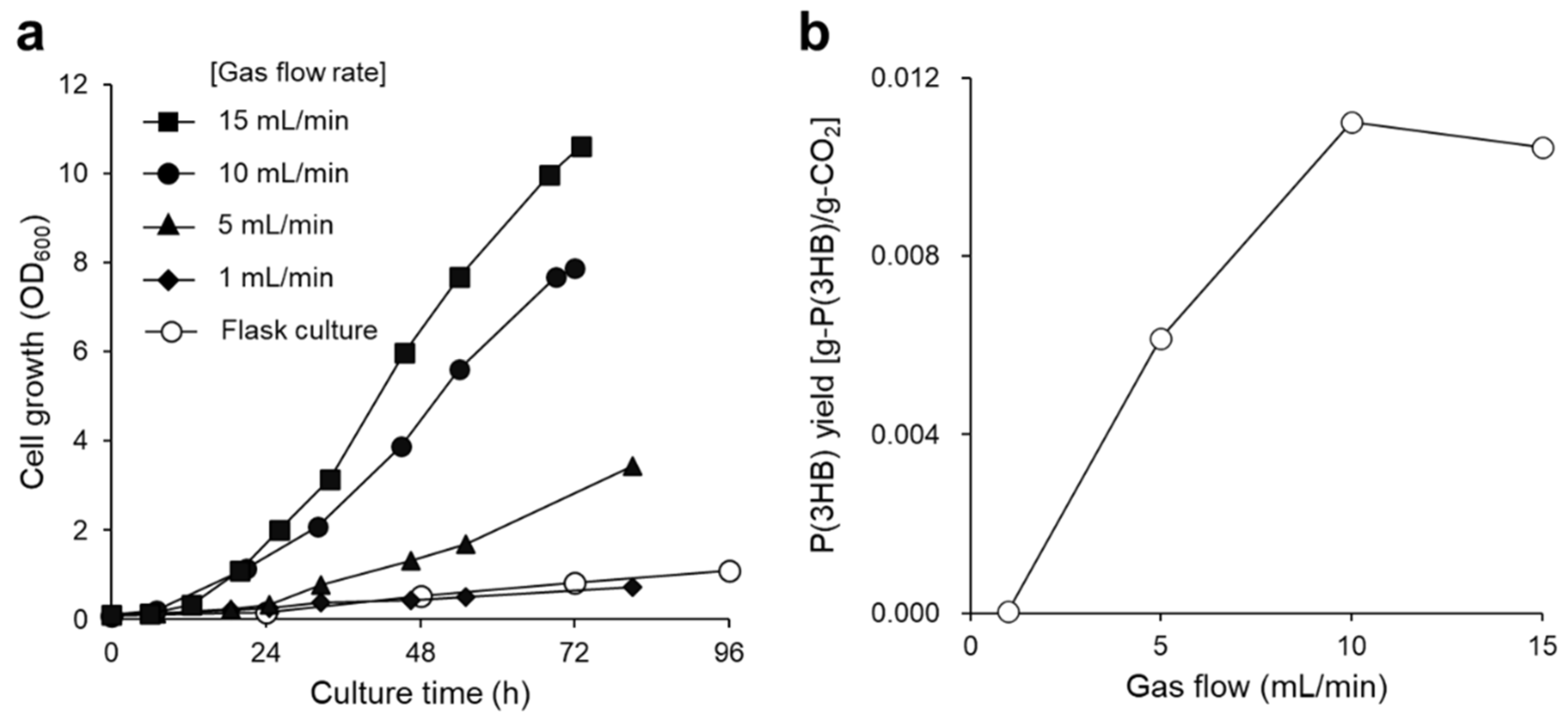
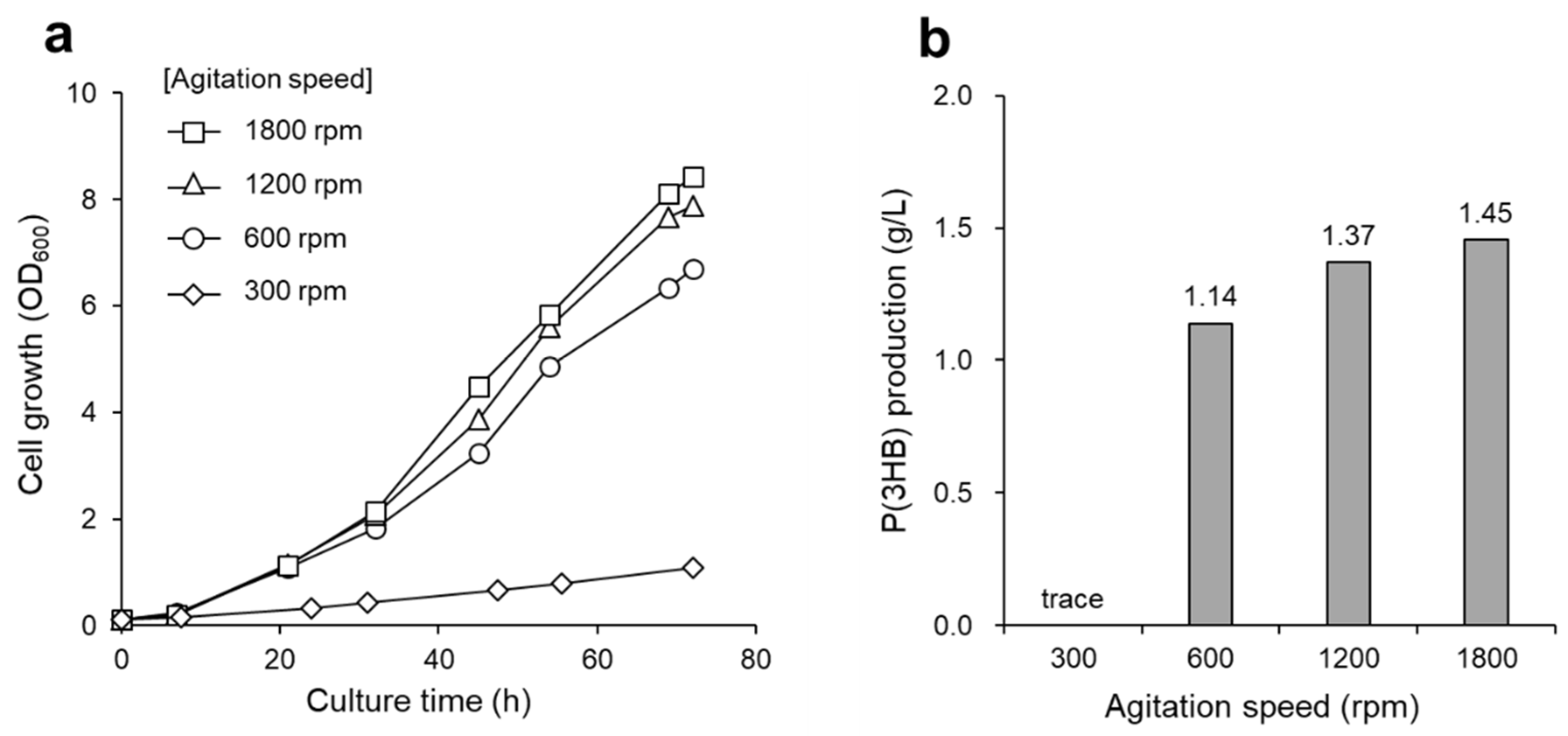
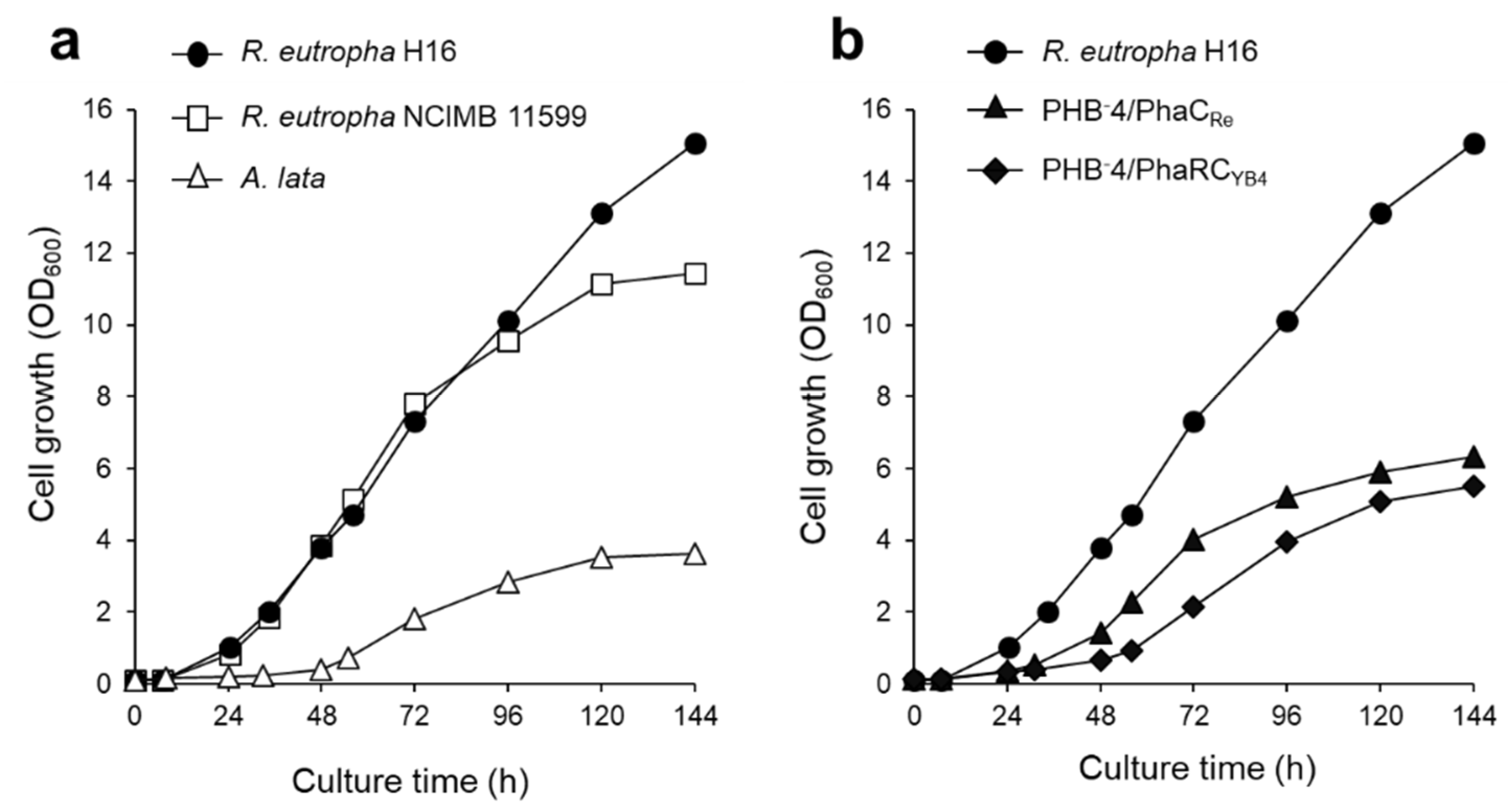
3.1. P(3HB) Production by a Continuous Supply of Non-Combustible Gas Mixture
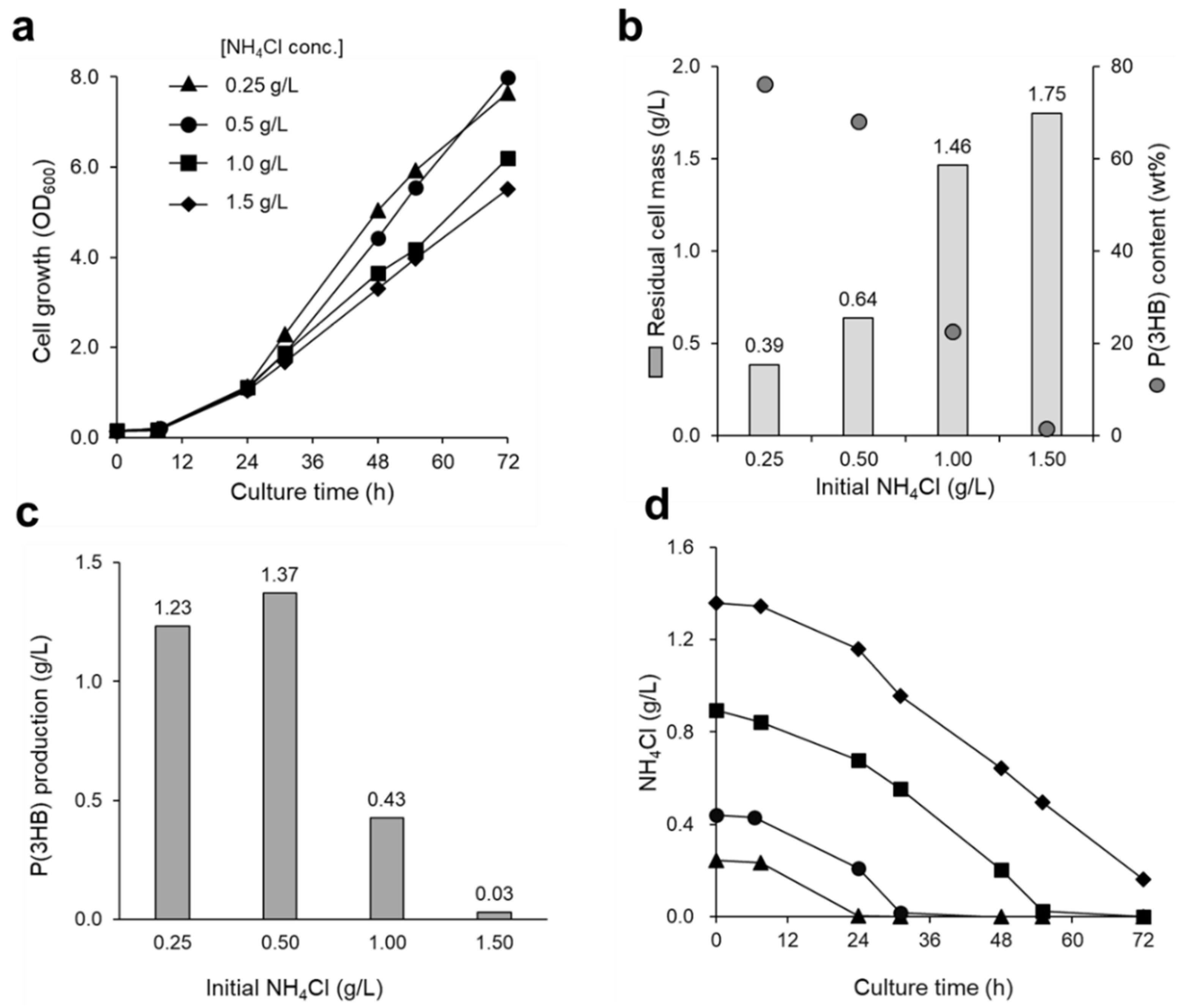
3.2. Effect of Nitrogen Source on P(3HB) Accumulation
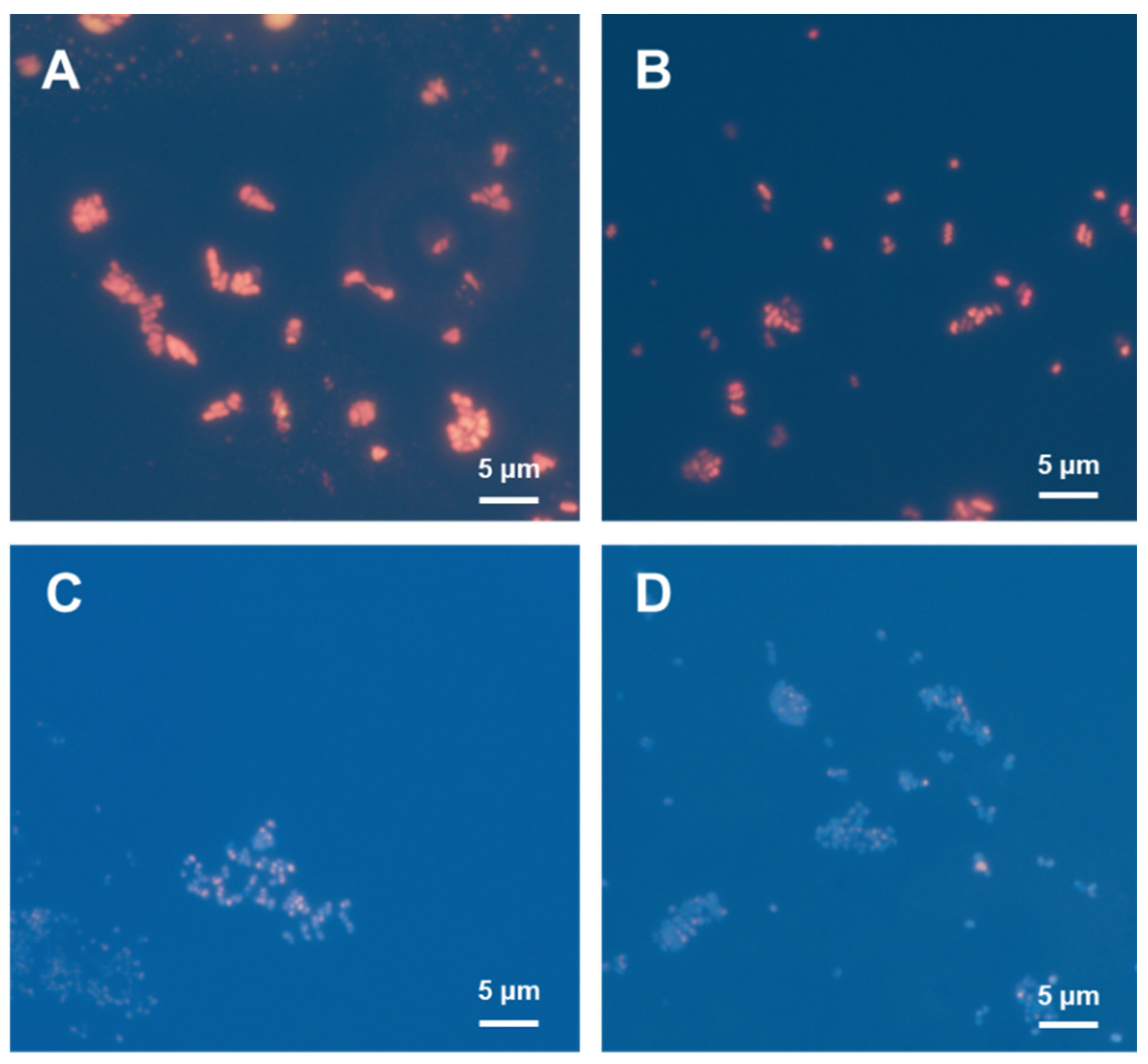
3.3. Hydrogen-Oxidizing Bacteria Grown in a Safe Culture System with a Low-Hydrogen-Content Gas Mixture
3.4. Autotrophic P(3HB) Production by Using Recombinant Strains

4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Borrelle, S.B.; Ringma, J.; Law, K.L.; Monnahan, C.C.; Lebreton, L.; McGivern, A.; Murphy, E.; Jambeck, J.; Leonard, G.H.; Hilleary, M.A.; et al. Predicted growth in plastic waste exceeds efforts to mitigate plastic pollution. Science 2020, 369, 1515–1518. [Google Scholar] [CrossRef]
- Sudesh, K.; Abe, H.; Doi, Y. Synthesis, structure and properties of polyhydroxyalkanoates: Biological polyesters. Prog. Polym. Sci. 2000, 25, 1503–1555. [Google Scholar] [CrossRef]
- Akiyama, M.; Tsuge, T.; Doi, Y. Environmental life cycle comparison of polyhydroxyalkanoates produced from renewable carbon resources by bacterial fermentation. Polym. Degrad. Stabil. 2003, 80, 183–194. [Google Scholar] [CrossRef]
- Troschl, C.; Meixner, K.; Drosg, B. Cyanobacterial PHA production—Review of recent advances and a summary of three years’ working experience running a pilot plant. Bioengineering 2017, 4, 26. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Sharma, L.; Mallick, N.; Mala, J. Progress and challenges in producing polyhydroxyalkanoate biopolymers from cyanobacteria. J. Appl. Phycol. 2017, 29, 1213–1232. [Google Scholar] [CrossRef]
- Ishizaki, A.; Tanaka, K. Production of poly-β-Hydroxybutyric acid from carbon dioxide by Alcaligenes eutrophus ATCC 17697T. J. Ferment. Bioeng. 1991, 71, 254–257. [Google Scholar] [CrossRef]
- Ishizaki, A.; Tanaka, K.; Takeshita, T.; Kanemaru, T.; Shimoji, T.; Kawano, T. Equipment and operation for fermentative PHB production using gaseous substrate to guarantee safety from explosion. J. Chem. Eng. Jpn. 1993, 26, 225–227. [Google Scholar] [CrossRef]
- Taga, N.; Tanaka, K.; Ishizaki, A. Effects of rheological change by addition of carboxymethylcellulose in culture media of an air-lift fermentor on poly-D-3-hydroxybutyric acid productivity in autotrophic culture of hydrogen-oxidizing bacterium, Alcaligenes eutrophus. Biotechnol. Bioeng. 1997, 53, 529–533. [Google Scholar] [CrossRef]
- Sugimoto, T.; Tsuge, T.; Tanaka, K.; Ishizaki, A. Control of acetic acid concentration by pH-stat continuous substrate feeding in heterotrophic culture phase of two-stage cultivation of Alcaligenes eutrophus for production of P(3HB) from CO2, H2, and O2 under non-explosive conditions. Biotechnol. Bioeng. 1999, 62, 625–631. [Google Scholar] [CrossRef]
- Garcia-gonzalez, L.; Islam, S.; Dubreuil, M.; Volcke, E.I.P.; De Wever, H. Sustainable autotrophic production of polyhydroxybutyrate (PHB) from CO2 using a two-stage cultivation system. Catal. Today 2015, 257, 237–245. [Google Scholar] [CrossRef]
- Ghysels, S.; Mozumder, M.S.I.; De Wever, H.; Volcke, E.I.P.; Garcia-Gonzalez, L. Targeted poly(3-hydroxybutyrate-co-3-hydroxyvalerate) bioplastic production from carbon dioxide. Bioresour. Technol. 2018, 249, 858–868. [Google Scholar] [CrossRef]
- Tanaka, K.; Ishizaki, A.; Kanamaru, T.; Kawano, T. Production of poly(D-3-hydroxybutyrate) from CO2, H2, and O2 by high cell density autotrophic cultivation of Alcaligenes eutrophus. Biotechnol. Bioeng. 1995, 45, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Ishizaki, A.; Tanaka, K. Batch culture of Alcaligenes eutrophus ATCC 17697 T using recycled gas closed circuit culture system. J. Ferment. Bioeng. 1990, 69, 170–174. [Google Scholar] [CrossRef]
- Miyahara, Y.; Yamamoto, M.; Thorbecke, R.; Mizuno, S.; Tsuge, T. Autotrophic biosynthesis of polyhydroxyalkanoate by Ralstonia eutropha from non-combustible gas mixture with low hydrogen content. Biotechnol. Lett. 2020, 42, 1655–1662. [Google Scholar] [CrossRef] [PubMed]
- Thorbecke, R.; Yamamoto, M.; Miyahara, Y.; Oota, M.; Mizuno, S.; Tsuge, T. The gene dosage effect of carbonic anhydrase on the biosynthesis of poly(3-hydroxybutyrate) under autotrophic and mixotrophic culture conditions. Polym. J. 2021, 53, 209–213. [Google Scholar] [CrossRef]
- Ushimaru, K.; Motoda, Y.; Numata, K.; Tsuge, T. Phasin proteins activate Aeromonas caviase polyhydroxyalkanoate (PHA) synthase but not Ralstonia eutropha PHA synthase. Appl. Environ. Microbiol. 2014, 80, 2867–2873. [Google Scholar] [CrossRef] [PubMed]
- Hyakutake, M.; Saito, Y.; Tomizawa, S.; Mizuno, K.; Tsuge, T. Polyhydroxyalkanoate (PHA) synthesis by Class IV PHA synthases employing Ralstonia eutropha PHB-4 as host strain. Biosci. Biotechnol. Biochem. 2011, 75, 1615–1617. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kovach, M.E.; Elzer, P.H.; Hill, D.S.; Robertson, G.T.; Farris, M.A.; Roop, R.M., II; Peterson, K.M. Four new derivatives of the broad-host-range cloning vector pBBR1MCS, carrying different antibiotic-resistance cassettes. Gene 1995, 166, 175–176. [Google Scholar] [CrossRef]
- Tanadchangsaeng, N.; Kitagawa, A.; Yamamoto, T.; Abe, H.; Tsuge, T. Identification, biosynthesis, and characterization of polyhydroxyalkanoate copolymer consisting of 3-hydroxybutyrate and 3-hydroxy-4-methylvalerate. Biomacromolecules 2009, 10, 2866–2874. [Google Scholar] [CrossRef] [PubMed]
- Miyahara, Y.; Oota, M.; Tsuge, T. NADPH supply for poly(3-Hydroxybutyrate) synthesis concomitant with enzymatic oxidation of phosphite. J. Biosci. Bioeng. 2018, 126, 764–768. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, T. Metabolic improvements and use of inexpensive carbon sources in microbial production of polyhydroxyalkanoates. J. Biosci. Bioeng. 2002, 94, 579–584. [Google Scholar] [CrossRef]
- Ostle, A.G.; Holt, J.G. Nile Blue A as a fluorescent stain for poly-β-hydroxybutyrate. Appl. Environ. Microbiol. 1982, 44, 238–241. [Google Scholar] [CrossRef] [PubMed]
- Ryu, H.W.; Hahn, S.K.; Chang, Y.K.; Chang, H.N. Production of poly(3-hydroxybutyrate) by high cell density fed-batch culture of Alcaligenes eutrophus with phosphate limitation. Biotechnol. Bioeng. 1997, 55, 28–32. [Google Scholar] [CrossRef]
- Shang, L.; Jiang, M.; Chang, H.N. Poly(3-Hydroxybutyrate) synthesis in fed-batch culture of Ralstonia eutropha with phosphate limitation under different glucose concentrations. Biotechnol. Lett. 2003, 25, 1415–1419. [Google Scholar] [CrossRef]
- Jo, S.Y.; Sohn, Y.J.; Park, S.Y.; Son, J.; Yoo, J.I.; Baritugo, K.A.; David, Y.; Kang, K.H.; Kim, H.; Choi, J.-I.; et al. Biosynthesis of polyhydroxyalkanoates from sugarcane molasses by recombinant Ralstonia eutropha strains. Korean J. Chem. Eng. 2021, 38, 1452–1459. [Google Scholar] [CrossRef]
- Orita, I.; Iwazawa, R.; Nakamura, S.; Fukui, T. Identification of mutation points in Cupriavidus necator NCIMB 11599 and genetic reconstitution of glucose-utilization ability in wild strain H16 for polyhydroxyalkanoate production. J. Biosci. Bioeng. 2012, 113, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Yamane, T.; Fukunaga, M.; Lee, Y.W. Increased PHB productivity by high-cell-density fed-batch culture of Alcaligenes latus, a growth-associated PHB producer. Biotechnol. Bioeng. 1996, 50, 197–202. [Google Scholar] [CrossRef]
- Wang, F.; Lee, S.Y. Poly(3-hydroxybutyrate) production with high productivity and high polymer content by a fed-batch culture of Alcaligenes latus under nitrogen limitation. Appl. Environ. Microbiol. 1997, 63, 3703–3706. [Google Scholar] [CrossRef]
- Malik, K.A.; Jung, C.; Claus, D.; Schlegel, H.G. Nitrogen fixation by the hydrogen-oxidizing bacterium Alcaligenes latus. Arch. Microbiol. 1981, 129, 254–256. [Google Scholar] [CrossRef]
- Fukui, T.; Doi, Y. Efficient production of polyhydroxyalkanoates from plant oils by Alcaligenes eutrophus and its recombinant strain. Appl. Microbiol. Biotechnol. 1998, 49, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Kahar, P.; Tsuge, T.; Taguchi, K.; Doi, Y. High yield production of polyhydroxyalkanoates from soybean oil by Ralstonia eutropha and its recombinant strain. Polym. Degrad. Stabil. 2004, 83, 79–86. [Google Scholar] [CrossRef]
- Mifune, J.; Nakamura, S.; Fukui, T. Engineering of pha operon on Cupriavidus necator chromosome for efficient biosynthesis of poly(3-Hydroxybutyrate-co-3-hydroxyhexanoate) from vegetable oil. Polym. Degrad. Stabil. 2010, 95, 1305–1312. [Google Scholar] [CrossRef]
- Sato, S.; Maruyama, H.; Fujiki, T.; Matsumoto, K. Regulation of 3-hydroxyhexanoate composition in PHBH synthesized by recombinant Cupriavidus necator H16 from plant oil by using butyrate as a co-substrate. J. Biosci. Bioeng. 2015, 120, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Tsuge, T.; Watanabe, S.; Shimada, D.; Abe, H.; Doi, Y.; Taguchi, S. Combination of N149S and D171G mutations in Aeromonas caviae polyhydroxyalkanoate synthase and impact on polyhydroxyalkanoate biosynthesis. FEMS Microbiol. Lett. 2007, 277, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Riedel, S.L.; Bader, J.; Brigham, C.J.; Budde, C.F.; Yusof, Z.A.M.; Rha, C.; Sinskey, A.J. Production of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) by Ralstonia eutropha in high cell density palm oil fermentations. Biotechnol. Bioeng. 2012, 109, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Nangle, S.N.; Ziesack, M.; Buckley, S.; Trivedi, D.; Loh, D.M.; Nocera, D.G.; Silver, P.A. Valorization of CO2 through lithoautotrophic production of sustainable chemicals in Cupriavidus necator. Metab. Eng. 2020, 62, 207–220. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.; Ishizaki, A. Production of poly-D-3-hydroxybutyric acid from carbon dioxide by a two-stage culture method employing Alcaligenes eutrophus ATCC 17697T. J. Ferment. Bioeng. 1994, 77, 425–427. [Google Scholar] [CrossRef]
- Heinzle, E.; Lafferty, R.M. A kinetic model for growth and synthesis of poly-β-hydroxybutyric acid (PHB) in Alcaligenes eutrophus H16. Eur. J. Appl. Microbiol. Biotechnol. 1980, 11, 8–16. [Google Scholar] [CrossRef]
- Park, I.; Hea, E.; Kyoungphile, J. Optimization of carbon dioxide and valeric acid utilization for polyhydroxyalkanoates synthesis by Cupriavidus necator. J. Polym. Environ. 2014, 22, 244–251. [Google Scholar] [CrossRef]
- Tanaka, K.; Yoshida, K.; Orita, I.; Fukui, T. Biosynthesis of poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) from CO2 by a recombinant Cupriavidus necator. Bioengineering 2021, 8, 179. [Google Scholar] [CrossRef]
- Iwata, T.; Aoyagi, Y.; Fujita, M.; Yamane, H.; Doi, Y.; Suzuki, Y.; Takeuchi, A.; Uesugi, K. Processing of a strong biodegradable poly[(R)-3-hydroxybutyrate] fiber and a new fiber structure revealed by micro-beam X-ray diffraction with synchrotron radiation. Macromol. Rapid Commun. 2004, 25, 1100–1104. [Google Scholar] [CrossRef]
- Iwata, T.; Aoyagi, Y.; Tanaka, T.; Fujita, M.; Takeuchi, A.; Suzuki, Y.; Uesugi, K. Microbeam X-ray diffraction and enzymatic degradation of poly [(R)-3-hydroxybutyrate] fibers with two kinds of molecular conformations. Macromolecules 2006, 39, 5789–5795. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Dry Cell wt. (g/L) | Residual Cell Mass (g/L) | PHA Content (wt%) | PHA (g/L) | Molecular Weight a | |
|---|---|---|---|---|---|---|
| Mw (×105) | PDI | |||||
| R. eutropha H16 | 3.31 | 0.37 | 89 | 2.94 | 13.5 | 1.87 |
| R. eutropha NCIMB 11599 | 2.46 | 0.67 | 73 | 1.79 | - | - |
| A. lata | 0.83 | 0.61 | 27 | 0.22 | - | - |
| R. eutropha PHB-4/PhaCRe | 1.55 | 0.51 | 67 | 1.04 | 1.54 | 1.39 |
| R. eutropha PHB-4/PhaRCYB4 | 1.17 | 0.48 | 59 | 0.69 | 16.9 | 1.57 |
| R. eutropha H16/pBBR1MCS-2 b (Flask culture) | 0.39 | 0.12 | 70 | 0.27 | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyahara, Y.; Wang, C.-T.; Ishii-Hyakutake, M.; Tsuge, T. Continuous Supply of Non-Combustible Gas Mixture for Safe Autotrophic Culture to Produce Polyhydroxyalkanoate by Hydrogen-Oxidizing Bacteria. Bioengineering 2022, 9, 586. https://doi.org/10.3390/bioengineering9100586
Miyahara Y, Wang C-T, Ishii-Hyakutake M, Tsuge T. Continuous Supply of Non-Combustible Gas Mixture for Safe Autotrophic Culture to Produce Polyhydroxyalkanoate by Hydrogen-Oxidizing Bacteria. Bioengineering. 2022; 9(10):586. https://doi.org/10.3390/bioengineering9100586
Chicago/Turabian StyleMiyahara, Yuki, Chih-Ting Wang, Manami Ishii-Hyakutake, and Takeharu Tsuge. 2022. "Continuous Supply of Non-Combustible Gas Mixture for Safe Autotrophic Culture to Produce Polyhydroxyalkanoate by Hydrogen-Oxidizing Bacteria" Bioengineering 9, no. 10: 586. https://doi.org/10.3390/bioengineering9100586
APA StyleMiyahara, Y., Wang, C.-T., Ishii-Hyakutake, M., & Tsuge, T. (2022). Continuous Supply of Non-Combustible Gas Mixture for Safe Autotrophic Culture to Produce Polyhydroxyalkanoate by Hydrogen-Oxidizing Bacteria. Bioengineering, 9(10), 586. https://doi.org/10.3390/bioengineering9100586