
Molecular Tools and Their Applications in Developing Salt-Tolerant Soybean (Glycine max L.) Cultivars


 ,
, 

 and
and
Abstract
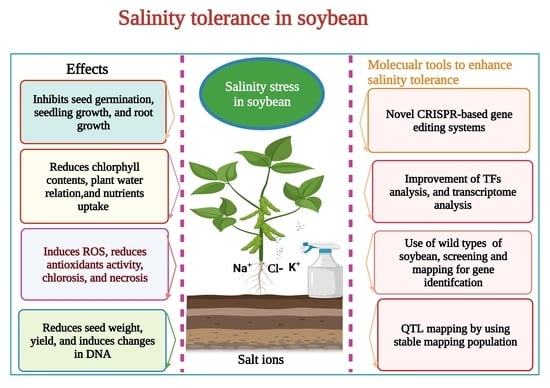
1. Introduction
2. Effects of Salinity Stress on Soybean

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Effects | References |
|---|---|
| Salinity stress affected the activities of antioxidants | [46] |
| Caused ionic imbalance, and enhanced electrolyte leakage | [3] |
| Reduced growth and biomass in soybean cultivars | [36] |
| Salinity stress affected the seed germination percentage and seedlings growth | [37] |
| Reduced the NPK contents in plants | [38] |
| Total phenol contents reduced | [47] |
| Salinity stress decreased the contents of protein and phospholipids | [44] |
| Salinity stress decreased yield by affecting number of seeds/plant | [43] |
| Salt stress decreased the level of abscisic acid and hydrogen peroxide | [40] |
| Reduced oil contents | [45] |
| Reduced stomal conductance | [42] |
| Suppressed GA and ABA levels in cell | [41] |
| Generation of ROS | [35] |
| Leaf reflectance and chlorophyll contents were decreased | [39] |
| Reduced leaf length, fresh weight, and plant height | [34] |
3. Genetic Mechanism of Salinity Tolerance in Soybean
4. Genetic Diversity
5. Mapping of QTL for Salinity Tolerance in Soybean
| Population/Parents | QTL | Chromosome | References |
|---|---|---|---|
| Cheongja 3, IT162669 | qST6 | 6 | [48] |
| RIL | qSTR3 | 3 | [71] |
| 132 F2 families (Williams 82 × Fiskeby III) | qLSC13 | 13 | [72] |
| F2:3 mapping population (Ozark × Jake) | qLCC3 | 3 | [73] |
| RA-452 and Osage | qLCC15 | 15 | [78] |
| RIL | qNcl3 | 3 | [59] |
| RIL | qST-GR-8, qST-IR-8 | 8 | [76] |
| F4:6 (RA-452 × Osage) | qLCC13 | 13 | [79] |
| RIL | qST3 | 3 | [74] |
| F6 RIL | qSTR17 | 17 | [77] |
| 96 RIL | qSTR3 | 3 | [75] |
6. GWAS for Salt Tolerance in Soybean
7. Identifying GWAS-Based Genes for Salt Tolerance in Soybean
8. Genetic Engineering for Salinity Tolerance in Soybean
| Gene | Role | Transformation Method | References |
|---|---|---|---|
| GmLecRlk | Enhanced fresh weight, proline content, as well as catalase activity | Agrobacterium rhizogenes, EHA105 | [99] |
| GmNFYA | Induced expression of salt-responsive genes | Agrobacterium tumefaciens-mediated cotyledon node transformation | [95] |
| AgGlpF | Enhanced salt tolerance | Soybean cotyledonary node transformation method | [90] |
| J (ortholog of AtELF3) | Controlled the expression of stress-related genes (GmWRKY27 and GmNAC) | Agrobacterium rhizogenes strain K599 | [23] |
| p68 | Increased photosynthetic rate, stomatal conductance, and CO2 assimilation | Agrobacterium tumefaciens strain EHA105 | [98] |
| PgTIP1 | Developed better leaf stomatal movement as well as water–gas exchange capabilities | Pollen-tube pathway method | [64] |
| GmST1 | Reduced ROS production and increased sensitivity to ABA | Agrobacterium tumefaciens GV3101 | [91] |
| GmsSOS1 | Improved seed germination and seedling growth | Agrobacterium tumefaciens | [94] |
| StP5CS | Increased the plant height, leaf area, chlorophyll contents, and number of fresh pods | Agrobacterium-mediated cotyledonary-node method | [93] |
| GmPIP1;6 | Enhanced leaf gas exchange rate | Agrobacterium tumefaciens | [92] |
| TaNHX2 | Enhanced biomass, flowers per plant, and long survival rate under salt stress conditions | Agrobacterium rhizogenes-mediated transformation | [65] |
| GmBIN2 | Increased the relative root growth and upregulated stress-responsive genes | Agrobacterium rhizogenes K599 | [97] |
9. CRISPR/Cas9-Mediated Salt Tolerance in Soybean
10. Evidence of Transcription Factors (TFs) Analysis for Salt Tolerance in Soybean
| TFs/Genes | Role | References |
|---|---|---|
| GmTGA13 | Absorption of K+, regulation of ions homeostasis, and activation of several stress-responsive genes | [115] |
| GmLecRlk | Increased proline content, fresh weight, and scavenging ability of antioxidants | [99] |
| GmbHLH3 | Increased the accumulation rate of Cl− and NO3− in roots | [111] |
| GmNAC06 | Enhanced proline and glycine betaine contents | [6] |
| GmMYB46 | Mediated salt stress by the complex regulatory network | [119] |
| GmMYB84 | GmMYB84 overexpressed, and plants witnessed high germination rate, root elongation, membrane integrity, and low K+ level | [107] |
| GmTGA17 | Enhanced chlorophyll content and proline contents under salt stress | [112] |
| GmCYP81E11 | Increased the flavonoid biosynthesis | [110] |
| GmDREB6 | Increased proline contents | [117] |
| GmNAC15 | Enhanced activation of several genes | [113] |
| GmWRKY12 | Overexpression increased proline contents under salt stress | [106] |
| MsWRKY11 | Enhanced chlorophyll contents, soluble sugar, catalase activity, plant height, and pods per plant | [66] |
| GmFDL19 | Higher shoot weight, plant height, and germination rate | [114] |
| GmWRKYs | Overexpressed and enhanced salt tolerance | [108] |
| HD-Zip | Improved salt tolerance by unique pattern of expression | [118] |
| OsDREB2A | Increased soluble sugars and proline contents and increased the expression of certain stress-responsive genes | [109] |
11. Mathematical Modeling Approaches for Salinity Tolerance
12. Conclusions and Future Research Directions
Author Contributions
Funding
Conflicts of Interest
References
- Rahman, S.U.; McCoy, E.; Raza, G.; Ali, Z.; Mansoor, S.; Amin, I. Improvement of soybean, a way forward transition from genetic engineering to new plant breeding technologies. Mol. Biotechnol. 2022. online ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Sahile, A.A.; Jan, R.; Asaf, S.; Hamayun, M.; Imran, M.; Adhikari, A.; Kang, S.-M.; Kim, K.-M.; Lee, I.-J. Halotolerant bacteria mitigate the effects of salinity stress on soybean growth by regulating secondary metabolites and molecular responses. BMC Plant Biol. 2021, 21, 176. [Google Scholar] [CrossRef] [PubMed]
- Nigam, B.; Dubey, R.S.; Rathore, D. Protective role of exogenously supplied salicylic acid and PGPB (Stenotrophomonas sp.) on spinach and soybean cultivars grown under salt stress. Sci. Hortic. 2022, 293, 110654. [Google Scholar] [CrossRef]
- Battisti, I.; Ebinezer, L.B.; Lomolino, G.; Masi, A.; Arrigoni, G. Protein profile of commercial soybean milks analyzed by label-free quantitative proteomics. Food Chem. 2021, 352, 129299. [Google Scholar] [CrossRef]
- Silva, B.; Batista, B.; Lobato, A. Anatomical changes in stem and root of soybean plants submitted to salt stress. Plant Biol. 2021, 23, 57–65. [Google Scholar] [CrossRef]
- Li, M.; Chen, R.; Jiang, Q.; Sun, X.; Zhang, H.; Hu, Z. GmNAC06, a NAC domain transcription factor enhances salt stress tolerance in soybean. Plant Mol. Biol. 2021, 105, 333–345. [Google Scholar] [CrossRef]
- Afrin, S.; Akhtar, N.; Khanam, T.; Hossain, F. Alleviative effects of zinc on biomass yield and antioxidative enzymes activity in leaves of soybean (Glycine max L.) under salt stress. Am. J. Agric. For. 2021, 9, 147–155. [Google Scholar] [CrossRef]
- Rasheed, A.; Li, H.; Nawaz, M.; Mahmood, A.; Hassan, M.U.; Shah, A.N.; Hussain, F.; Azmat, S.; Gillani, S.F.A.; Majeed, Y. Molecular tools, potential frontiers for enhancing salinity tolerance in rice: A critical review and future prospective. Front. Plant Sci. 2022, 13, 966749. [Google Scholar] [CrossRef]
- Raza, A.; Tabassum, J.; Fakhar, A.Z.; Sharif, R.; Chen, H.; Zhang, C.; Ju, L.; Fotopoulos, V.; Siddique, K.H.; Singh, R.K. Smart reprograming of plants against salinity stress using modern biotechnological tools. Crit. Rev. Biotechnol. 2022, 1–28. [Google Scholar] [CrossRef]
- Tran, H.A.; Nguyen, C.T.; Lee, J.D.; Seo, H.S.; Song, J.T. Screening for Flooding Tolerance in Two Soybean Populations (EMS-Induced ‘Pungsannamul’ Population and Korean Glycine Soja Core Population). Available online: https://scholar.kyobobook.co.kr/article/detail/4010028375185 (accessed on 31 July 2022).
- Das, A.K.; Anik, T.R.; Rahman, M.; Keya, S.S.; Islam, M.; Sultana, S.; Ghosh, P.K.; Khan, S.; Ahamed, T.; Ghosh, T.K. Ethanol treatment enhances physiological and biochemical responses to mitigate saline toxicity in soybean. Plants 2022, 11, 272. [Google Scholar] [CrossRef]
- Al-Zahrani, H.S.; Nahar, K.; Alharby, H.F.; Alsamadany, H.; Hakeem, K.R.; Hasanuzzaman, M. Zinc Supplementation Enhances Glutathione-Mediated Antioxidant Defense and Glyoxalase Systems to Conferring Salt Tolerance in Soybean (Glycine max L.). Agronomy 2022, 12, 1032. [Google Scholar] [CrossRef]
- Phang, T.H.; Shao, G.; Lam, H.M. Salt tolerance in soybean. J. Integr. Plant Biol. 2008, 50, 1196–1212. [Google Scholar] [CrossRef]
- Li, H.; Wang, X.H.; Li, Q.; Xu, P.; Liu, Z.N.; Xu, M.; Cui, X.Y. GmCIPK21, a CBL-interacting protein kinase confers salt tolerance in soybean (Glycine max L.). Plant Physiol. Biochem. 2022, 184, 47–55. [Google Scholar] [CrossRef]
- Papiernik, S.K.; Grieve, C.M.; Lesch, S.M.; Yates, S.R. Effects of salinity, imazethapyr, and chlorimuron application on soybean growth and yield. Commun. Soil Sci. Plant Anal. 2005, 36, 951–967. [Google Scholar] [CrossRef]
- Pathan, M.S.; Lee, J.-D.; Shannon, J.G.; Nguyen, H.T. Recent advances in breeding for drought and salt stress tolerance in soybean. In Advances in Molecular Breeding toward Drought and Salt Tolerant Crops; Springer: Dordrecht, The Netherlands, 2007; pp. 739–773. [Google Scholar] [CrossRef]
- Chen, H.T.; Liu, X.-Q.; Zhang, H.M.; Yuan, X.X.; Gu, H.P.; Cui, X.Y.; Xin, C. Advances in salinity tolerance of soybean: Genetic diversity, heredity, and gene identification contribute to improving salinity tolerance. J. Integr. Agric. 2018, 17, 2215–2221. [Google Scholar] [CrossRef]
- Kaashyap, M.; Ford, R.; Bohra, A.; Kuvalekar, A.; Mantri, N. Improving salt tolerance of chickpea using modern genomics tools and molecular breeding. Curr. Genom. 2017, 18, 557–567. [Google Scholar] [CrossRef]
- Jha, U.C.; Bohra, A.; Jha, R.; Parida, S.K. Salinity stress response and ‘omics’ approaches for improving salinity stress tolerance in major grain legumes. Plant Cell Rep. 2019, 38, 255–277. [Google Scholar] [CrossRef]
- Qi, X.; Li, M.; Xie, M.; Liu, X.; Ni, M.; Shao, G.; Song, C.; Yim, A.; Tao, Y.; Wong, F. Identification of a novel salt tolerance gene in wild soybean by whole-genome sequencing. Nat. Commun. 2014, 5, 4340. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M. Improving salinity tolerance in cereals. Crit. Rev. Plant Sci. 2013, 32, 237–249. [Google Scholar] [CrossRef]
- Huang, L. Genome-Wide Association Mapping Identifies QTLs and Candidate Genes for Salt Tolerance in Soybean. Master’s Thesis, University of Arkansas, Fayetteville, AR, USA, 20 August 2013. [Google Scholar]
- Cheng, Q.; Gan, Z.; Wang, Y.; Lu, S.; Hou, Z.; Li, H.; Xiang, H.; Liu, B.; Kong, F.; Dong, L. The soybean gene J contributes to salt stress tolerance by up-regulating salt-responsive genes. Front. Plant Sci. 2020, 11, 272. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Li, Y.; Liu, B.; Kong, F.; Tran, L.S.P. Adaptive mechanisms of soybean grown on salt-affected soils. Land Degrad. Dev. 2018, 29, 1054–1064. [Google Scholar] [CrossRef]
- Li, T.Y.; Zhang, Y.; Liu, H.; Wu, Y.; Li, W.; Zhang, H. Stable expression of Arabidopsis vacuolar Na+/H+ antiporter gene AtNHX1, and salt tolerance in transgenic soybean for over six generations. Chin. Sci. Bull. 2010, 55, 1127–1134. [Google Scholar] [CrossRef]
- Di, Y.H.; Sun, X.J.; Hu, Z.; Jiang, Q.-Y.; Song, G.H.; Zhang, B.; Zhao, S.S.; Zhang, H. Enhancing the CRISPR/Cas9 system based on multiple GmU6 promoters in soybean. Biochem. Biophy Res. Commun. 2019, 519, 819–823. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Liao, X.; Cui, Y.; Ma, W.; Zhang, X.; Du, H.; Ma, Y.; Ning, L.; Wang, H.; Huang, F. A cation diffusion facilitator, GmCDF1, negatively regulates salt tolerance in soybean. PLoS Genet. 2019, 15, e1007798. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Song, L.; Shu, Y.; Wang, S.; Niu, J.; Wang, Z.; Yu, T.; Gu, W.; Ma, H. Comparative proteomic analysis of seedling leaves of different salt tolerant soybean genotypes. J. Proteom. 2012, 75, 1529–1546. [Google Scholar] [CrossRef] [PubMed]
- Ullah, A.; Li, M.; Noor, J.; Tariq, A.; Liu, Y.; Shi, L. Effects of salinity on photosynthetic traits, ion homeostasis and nitrogen metabolism in wild and cultivated soybean. PeerJ 2019, 7, e8191. [Google Scholar] [CrossRef] [PubMed]
- Aleem, M.; Aleem, S.; Sharif, I.; Wu, Z.; Aleem, M.; Tahir, A.; Atif, R.M.; Cheema, H.M.N.; Shakeel, A.; Lei, S. Characterization of SOD and GPX gene families in the soybeans in response to drought and salinity stresses. Antioxidants 2022, 11, 460. [Google Scholar] [CrossRef]
- Teakle, N.L.; Tyerman, S.D. Mechanisms of Cl-transport contributing to salt tolerance. Plant Cell Environ. 2010, 33, 566–589. [Google Scholar] [CrossRef]
- Luo, Q.; Yu, B.; Liu, Y. Differential sensitivity to chloride and sodium ions in seedlings of Glycine max and G. soja under NaCl stress. J. Plant Physiol. 2005, 162, 1003–1012. [Google Scholar] [CrossRef]
- Chen, X.-Q.; Yu, B.-J. Ionic effects of Na+ and Cl-on photosynthesis in Glycine max seedlings under isoosmotic salt stress. Plant Physiol. Mol. Biol. 2007, 33, 294–300. [Google Scholar]
- Amirjani, M. Effect of salinity stress on growth, mineral composition, proline content, antioxidant enzymes of soybean. Am. J. Plant Physiol. 2010, 5, 350–360. [Google Scholar] [CrossRef]
- Weisany, W.; Sohrabi, Y.; Heidari, G.; Siosemardeh, A.; Ghassemi-Golezani, K. Changes in antioxidant enzymes activity and plant performance by salinity stress and zinc application in soybean (‘Glycine max’L.). Plant Omics 2012, 5, 60–67. [Google Scholar]
- Sheteiwy, M.S.; Shao, H.; Qi, W.; Daly, P.; Sharma, A.; Shaghaleh, H.; Hamoud, Y.A.; El-Esawi, M.A.; Pan, R.; Wan, Q. Seed priming and foliar application with jasmonic acid enhance salinity stress tolerance of soybean (Glycine max L.) seedlings. J. Sci. Food Agric. 2021, 101, 2027–2041. [Google Scholar] [CrossRef]
- Çirka, M.; Kaya, A.R.; Eryiğit, T. Influence of temperature and salinity stress on seed germination and seedling growth of soybean (Glycine max L.). Legume Res. Int. J. 2021, 1, 7. [Google Scholar] [CrossRef]
- Otie, V.; Udo, I.; Shao, Y.; Itam, M.O.; Okamoto, H.; An, P.; Eneji, E.A. Salinity effects on morpho-physiological and yield traits of soybean (Glycine max L.) as mediated by foliar spray with brassinolide. Plants 2021, 10, 541. [Google Scholar] [CrossRef]
- Krezhova, D.D.; Kirova, E.B.; Yanev, T.K.; Iliev, I.T. Effects of salinity on leaf spectral reflectance and biochemical parameters of nitrogen fixing soybean plants (Glycine max L.). AIP Conf. Proc. 2010, 1203, 694–696. [Google Scholar] [CrossRef]
- Kataria, S.; Baghel, L.; Jain, M.; Guruprasad, K. Magnetopriming regulates antioxidant defense system in soybean against salt stress. Biocatal. Agric. Biotechnol. 2019, 18, 101090. [Google Scholar] [CrossRef]
- Shu, K.; Qi, Y.; Chen, F.; Meng, Y.; Luo, X.; Shuai, H.; Zhou, W.; Ding, J.; Du, J.; Liu, J. Salt stress represses soybean seed germination by negatively regulating GA biosynthesis while positively mediating ABA biosynthesis. Front. Plant Sci. 2017, 8, 1372. [Google Scholar] [CrossRef]
- He, Y.; Chen, Y.; Yu, C.; Lu, K.; Jiang, Q.; Fu, J.; Wang, G.; Jiang, D. Photosynthesis and yield traits in different soybean lines in response to salt stress. Photosynthetica 2016, 54, 630–635. [Google Scholar] [CrossRef]
- Sadak, M.S.; El-Hameid, A.; Asmaa, R.; Zaki, F.S.; Dawood, M.G.; El-Awadi, M.E. Physiological and biochemical responses of soybean (Glycine max L.) to cysteine application under sea salt stress. Bull. Natl. Res. Cent. 2020, 44, 1. [Google Scholar] [CrossRef]
- Liu, A.; Xiao, Z.; Wang, Z.; Lam, H.-M.; Chye, M.-L. Galactolipid and phospholipid profile and proteome alterations in soybean leaves at the onset of salt stress. Front. Plant Sci. 2021, 12, 383. [Google Scholar] [CrossRef] [PubMed]
- Parveen, A.-U.-H.M.; Akhtar, J.; Basra, S.M. Interactive effect of salinity and potassium on growth, biochemical parameters, protein and oil quality of soybean genotypes. Pak. J. Agric. Sci. 2016, 53, 69–78. [Google Scholar]
- Begum, N.; Hasanuzzaman, M.; Li, Y.; Akhtar, K.; Zhang, C.; Zhao, T. Seed Germination behavior, growth, physiology and antioxidant metabolism of four contrasting cultivars under combined drought and salinity in soybean. Antioxidants 2022, 11, 498. [Google Scholar] [CrossRef]
- Osman, M.S.; Badawy, A.A.; Osman, A.I.; Abdel Latef, A.A.H. Ameliorative impact of an extract of the halophyte Arthrocnemum macrostachyum on growth and biochemical parameters of soybean under salinity stress. J. Plant Growth Regul. 2021, 40, 1245–1256. [Google Scholar] [CrossRef]
- Cho, K.-H.; Kim, M.Y.; Kwon, H.; Yang, X.; Lee, S.-H. Novel QTL identification and candidate gene analysis for enhancing salt tolerance in soybean (Glycine max (L.) Merr.). Plant Sci. 2021, 313, 111085. [Google Scholar] [CrossRef]
- Wang, K.J.; Li, X.H. Interspecific gene flow and the origin of semi-wild soybean revealed by capturing the natural occurrence of introgression between wild and cultivated soybean populations. Plant Breed. 2011, 130, 117–127. [Google Scholar] [CrossRef]
- Li, M.W.; Jiang, B.; Han, T.; Zhang, G.; Lam, H.M. Genomic research on soybean and its impact on molecular breeding. Adv. Bot. Res. 2022, 102, 1–42. [Google Scholar]
- Lee, J.-D.; Shannon, J.G.; Vuong, T.D.; Nguyen, H.T. Inheritance of salt tolerance in wild soybean (Glycine soja Sieb. and Zucc.) accession PI483463. J. Hered. 2009, 100, 798–801. [Google Scholar] [CrossRef]
- Guan, R.; Chen, J.; Jiang, J.; Liu, G.; Liu, Y.; Tian, L.; Yu, L.; Chang, R.; Qiu, L.-j. Mapping and validation of a dominant salt tolerance gene in the cultivated soybean (Glycine max) variety Tiefeng 8. Crop J. 2014, 2, 358–365. [Google Scholar] [CrossRef]
- Hua-tao, C.; Xin, C.; De-yue, Y. Inheritance analysis and mapping quantitative trait loci (QTLs) associated with salt tolerance during seedling growth in soybean. Chin. J. Oil Crop Sci. 2011, 33, 231–234. [Google Scholar]
- Moniruzzaman, M.; Saiem, R.; Emon, R.; Haque, M.; Saha, N.; Malek, M.; Khatun, K. Genetic diversity analysis of soybean genotypes using SSR markers for salinity tolerance. Prog. Agric. 2019, 30, 1–9. [Google Scholar] [CrossRef][Green Version]
- Parker, M.B.; Gascho, G.; Gaines, T. Chloride toxicity of soybeans grown on atlantic coast flatwoods soils 1. Agron. J. 1983, 75, 439–443. [Google Scholar] [CrossRef]
- Yang, J.; Blanchar, R. Differentiating chloride susceptibility in soybean cultivars. Agron. J. 1993, 85, 880–885. [Google Scholar] [CrossRef]
- Zhang, W.J.; Niu, Y.; Bu, S.H.; Li, M.; Feng, J.Y.; Zhang, J.; Yang, S.X.; Odinga, M.M.; Wei, S.-P.; Liu, X.F. Epistatic association mapping for alkaline and salinity tolerance traits in the soybean germination stage. PLoS ONE 2014, 9, e84750. [Google Scholar] [CrossRef]
- Chen, P.; Yan, K.; Shao, H.; Zhao, S. Physiological mechanisms for high salt tolerance in wild soybean (Glycine soja) from Yellow River Delta, China: Photosynthesis, osmotic regulation, ion flux and antioxidant capacity. PLoS ONE 2013, 8, e83227. [Google Scholar] [CrossRef]
- Do, T.D.; Chen, H.; Hien, V.T.T.; Hamwieh, A.; Yamada, T.; Sato, T.; Yan, Y.; Cong, H.; Shono, M.; Suenaga, K. Ncl synchronously regulates Na+, K+ and Cl− in soybean and greatly increases the grain yield in saline field conditions. Sci. Rep. 2016, 6, 19147. [Google Scholar] [CrossRef]
- Genetic Analysis of Salt Tolerance in Soybean. Available online: https://pag.confex.com/pag/xxiv/webprogram/Paper20285.html (accessed on 25 July 2022).
- Wang, T.; Xun, H.; Wang, W.; Ding, X.; Tian, H.; Hussain, S.; Dong, Q.; Ying, L.Y.; Cheng, Y.; Wang, C. Mutation of GmAITR genes by CRISPR/Cas9 genome editing results in enhanced salinity stress tolerance in soybean. Front. Plant Sci. 2021, 12, 779598. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Ma, N.; Wang, C.; Fan, H.; Wang, M.; Zhang, J.; Cao, J.; Wang, D. A golgi-localized sodium/hydrogen exchanger positively regulates salt tolerance by maintaining higher K+/Na+ ratio in soybean. Front. Plant Sci. 2021, 12, 638340. [Google Scholar] [CrossRef]
- Du, Y.-T.; Zhao, M.-J.; Wang, C.-T.; Gao, Y.; Wang, Y.-X.; Liu, Y.-W.; Chen, M.; Chen, J.; Zhou, Y.-B.; Xu, Z.-S. Identification and characterization of GmMYB118 responses to drought and salt stress. BMC Plant Biol. 2018, 18, 320. [Google Scholar] [CrossRef] [PubMed]
- An, J.; Cheng, C.; Hu, Z.; Chen, H.; Cai, W.; Yu, B. The Panax ginseng PgTIP1 gene confers enhanced salt and drought tolerance to transgenic soybean plants by maintaining homeostasis of water, salt ions and ROS. Environ. Exp. Bot. 2018, 155, 45–55. [Google Scholar] [CrossRef]
- Cao, D.; Hou, W.; Liu, W.; Yao, W.; Wu, C.; Liu, X.; Han, T. Overexpression of TaNHX2 enhances salt tolerance of ‘composite’and whole transgenic soybean plants. Plant Cell Tissue Organ Cult. PCTOC 2011, 107, 541–552. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, L.; Chen, J.; Tao, L.; An, Y.; Cai, H.; Guo, C. Overexpression of the alfalfa WRKY11 gene enhances salt tolerance in soybean. PLoS ONE 2018, 13, e0192382. [Google Scholar] [CrossRef]
- Kassem, M.A. Salt Tolerance qtl mapping in soybean: 2004–2020. In Soybean Seed Composition; Springer: Berlin/Heidelberg, Germany, 2021; pp. 385–437. [Google Scholar]
- Cho, K.H.; Kwon, H.; Kim, M.Y.; Lee, S.H. Identification of Novel Loci Conferring Salt Tolerance in a Korean Soybean Landrace [Glycine max (L.) Merr.]. Available online: https://bmcgenomics.biomedcentral.com/articles/10.1186/s12864-019-5662-9 (accessed on 31 July 2022).
- Dhungana, S.K.; Park, J.-H.; Oh, J.-H.; Kang, B.-K.; Seo, J.-H.; Sung, J.-S.; Kim, H.-S.; Shin, S.-O.; Baek, I.-Y.; Jung, C.-S. Quantitative trait locus mapping for drought tolerance in soybean recombinant inbred line population. Plants 2021, 10, 1816. [Google Scholar] [CrossRef]
- Tuyen, D.; Lal, S.; Xu, D. Identification of a major QTL allele from wild soybean (Glycine soja Sieb. & Zucc.) for increasing alkaline salt tolerance in soybean. Theor Appl. Genet. 2010, 121, 229–236. [Google Scholar]
- Shi, X.; Yan, L.; Yang, C.; Yan, W.; Moseley, D.O.; Wang, T.; Liu, B.; Di, R.; Chen, P.; Zhang, M. Identification of a major quantitative trait locus underlying salt tolerance in ‘Jidou 12′soybean cultivar. BMC Res. Notes 2018, 11, 95. [Google Scholar] [CrossRef]
- Do, T.D.; Vuong, T.D.; Dunn, D.; Smothers, S.; Patil, G.; Yungbluth, D.C.; Chen, P.; Scaboo, A.; Xu, D.; Carter, T.E. Mapping and confirmation of loci for salt tolerance in a novel soybean germplasm, Fiskeby III. Theor Appl. Genet. 2018, 131, 513–524. [Google Scholar] [CrossRef]
- Lopez, C.; Orazaly, M.; Mozzoni, L.; Korth, K.L.; Chen, P. Quantitative trait loci for salt tolerance in soybean. J. Crop Improv. 2018, 32, 766–780. [Google Scholar] [CrossRef]
- Ha, B.-K.; Vuong, T.D.; Velusamy, V.; Nguyen, H.T.; Grover Shannon, J.; Lee, J.-D. Genetic mapping of quantitative trait loci conditioning salt tolerance in wild soybean (Glycine soja) PI 483463. Euphytica 2013, 193, 79–88. [Google Scholar] [CrossRef]
- Hamwieh, A.; Tuyen, D.; Cong, H.; Benitez, E.; Takahashi, R.; Xu, D. Identification and validation of a major QTL for salt tolerance in soybean. Euphytica 2011, 179, 451–459. [Google Scholar] [CrossRef]
- Kan, G.; Ning, L.; Li, Y.; Hu, Z.; Zhang, W.; He, X.; Yu, D. Identification of novel loci for salt stress at the seed germination stage in soybean. Breed. Sci. 2016, 66, 530–541. [Google Scholar] [CrossRef] [PubMed]
- Tuyen, D.; Zhang, H.; Xu, D. Validation and high-resolution mapping of a major quantitative trait locus for alkaline salt tolerance in soybean using residual heterozygous line. Mol. Breed. 2013, 31, 79–86. [Google Scholar] [CrossRef]
- Zeng, A.; Lara, L.; Chen, P.; Luan, X.; Hancock, F.; Korth, K.; Brye, K.; Pereira, A.; Wu, C. Quantitative trait loci for chloride tolerance in ‘Osage’soybean. Crop Sci. 2017, 57, 2345–2353. [Google Scholar] [CrossRef]
- Zeng, A. Quantitative Trait Loci Mapping, Genome-Wide Association Analysis, and Gene Expression of Salt Tolerance in Soybean; University of Arkansas: Fayetteville, AR, USA, 2016. [Google Scholar]
- Zeng, A.; Chen, P.; Korth, K.; Hancock, F.; Pereira, A.; Brye, K.; Wu, C.; Shi, A. Genome-wide association study (GWAS) of salt tolerance in worldwide soybean germplasm lines. Mol. Breed. 2017, 37, 1–14. [Google Scholar] [CrossRef]
- Do, T.D.; Vuong, T.D.; Dunn, D.; Clubb, M.; Valliyodan, B.; Patil, G.; Chen, P.; Xu, D.; Nguyen, H.T.; Shannon, J.G. Identification of new loci for salt tolerance in soybean by high-resolution genome-wide association mapping. BMC Genom. 2019, 20, 318. [Google Scholar] [CrossRef]
- Cao, Y.; Zhang, X.; Jia, S.; Karikari, B.; Zhang, M.; Xia, Z.; Zhao, T.; Liang, F. Genome-wide association among soybean accessions for the genetic basis of salinity-alkalinity tolerance during germination. Crop Past. Sci. 2021, 72, 255–267. [Google Scholar] [CrossRef]
- Shi, M.Q.; Xi-Liang, L.; Qian, Y.; Zhang, W.; Ya-Kai, L.; Bhat, A.J.; Gui-Zhen, K.; De-Yue, Y. Linkage and association mapping of wild soybean (Glycine soja) seeds germinating under salt stress. J. Int. Agric. 2022, in press. [Google Scholar] [CrossRef]
- Leng, Z.X.; Liu, Y.; Chen, Z.Y.; Guo, J.; Chen, J.; Zhou, Y.B.; Chen, M.; Ma, Y.Z.; Xu, Z.S.; Cui, X.Y. Genome-wide analysis of the DUF4228 family in soybean and functional identification of GmDUF4228–70 in response to drought and salt stresses. Front. Plant Sci. 2021, 12, 680. [Google Scholar] [CrossRef]
- Wang, T.T.; Yu, T.F.; Fu, J.-D.; Su, H.G.; Chen, J.; Zhou, Y.B.; Chen, M.; Guo, J.; Ma, Y.Z.; Wei, W.L. Genome-wide analysis of the GRAS gene family and functional identification of GmGRAS37 in drought and salt tolerance. Front. Plant Sci. 2020, 11, 2024. [Google Scholar] [CrossRef]
- Wang, Z.Q.; Yu, T.F.; Sun, G.Z.; Zheng, J.C.; Chen, J.; Zhou, Y.B.; Chen, M.; Ma, Y.Z.; Wei, W.L.; Xu, Z.S. Genome-wide analysis of the Catharanthus roseus RLK1-Like in soybean and GmCrRLK1L20 responds to drought and salt stresses. Front. Plant Sci. 2021, 12, 614909. [Google Scholar] [CrossRef]
- Patil, G.; Do, T.; Vuong, T.D.; Valliyodan, B.; Lee, J.-D.; Chaudhary, J.; Shannon, J.G.; Nguyen, H.T. Genomic-assisted haplotype analysis and the development of high-throughput SNP markers for salinity tolerance in soybean. Sci. Rep. 2016, 6, 19199. [Google Scholar] [CrossRef]
- Dong, L.; Hou, Z.; Li, H.; Li, Z.; Fang, C.; Kong, L.; Li, Y.; Du, H.; Li, T.; Wang, L. Agronomical selection on loss-of-function of GIGANTEA simultaneously facilitates soybean salt tolerance and early maturity. J. Integr. Plant Biol 2022. online ahead of print. [Google Scholar] [CrossRef]
- Abel, G.H.; MacKenzie, A.J. Salt Tolerance of soybean varieties (Glycine max L. Merrill) during germination and later growth 1. Crop Sci. 1964, 4, 157–161. [Google Scholar] [CrossRef]
- Li, F.; Ni, H.; Yan, W.; Xie, Y.; Liu, X.; Tan, X.; Zhang, L.; Zhang, S.-H. Overexpression of an aquaporin protein from Aspergillus glaucus confers salt tolerance in transgenic soybean. Transgenic Res. 2021, 30, 727–737. [Google Scholar] [CrossRef]
- Ren, S.; Lyle, C.; Jiang, G.-l.; Penumala, A. Soybean salt tolerance 1 (GmST1) reduces ROS production, enhances ABA sensitivity, and abiotic stress tolerance in Arabidopsis thaliana. Front. Plant Sci. 2016, 7, 445. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, C.; Liu, R.; Han, Q.; Vandeleur, R.K.; Du, J.; Tyerman, S.; Shou, H. Constitutive overexpression of soybean plasma membrane intrinsic protein GmPIP1; 6 confers salt tolerance. BMC Plant Biol. 2014, 14, 181. [Google Scholar] [CrossRef]
- Zhang, G.-C.; Zhu, W.-L.; Gai, J.-Y.; Zhu, Y.-L.; Yang, L.-F. Enhanced salt tolerance of transgenic vegetable soybeans resulting from overexpression of a novel Δ1-pyrroline-5-carboxylate synthetase gene from Solanum torvum Swartz. Hortic. Environ. Biotechnol. 2015, 56, 94–104. [Google Scholar] [CrossRef]
- Nie, W.-x.; Xu, L.; Yu, B.-j. A putative soybean GmsSOS1 confers enhanced salt tolerance to transgenic Arabidopsis sos1-1 mutant. Protoplasma 2015, 252, 127–134. [Google Scholar] [CrossRef]
- Lu, L.; Wei, W.; Tao, J.J.; Lu, X.; Bian, X.H.; Hu, Y.; Cheng, T.; Yin, C.C.; Zhang, W.K.; Chen, S.Y. Nuclear factor Y subunit GmNFYA competes with GmHDA13 for interaction with GmFVE to positively regulate salt tolerance in soybean. Plant Biotechnol. J. 2021, 19, 2362–2379. [Google Scholar] [CrossRef]
- Guan, R.; Guo, X.; Jiang, J.; Liu, Y.; Yu, L.; Chang, R.; Lijuan, Q. Identification of a novel salt tolerance-related locus in wild soybean (Glycine soja Sieb. & Zucc.). Front. Plant Sci. 2021, 12, 791175. [Google Scholar]
- Wang, L.-S.; Chen, Q.S.; Xin, D.W.; Qi, Z.M.; Zhang, C.; Li, S.N.; Jin, Y.M.; Mo, L.; Mei, H.Y.; Su, A.Y. Overexpression of GmBIN2, a soybean glycogen synthase kinase 3 gene, enhances tolerance to salt and drought in transgenic Arabidopsis and soybean hairy roots. J. Integr. Agric. 2018, 17, 1959–1971. [Google Scholar] [CrossRef]
- Karthik, S.; Tuteja, N.; Ganapathi, A.; Manickavasagam, M. Pea p68, a DEAD-box helicase, enhances salt tolerance in marker-free transgenic plants of soybean [Glycine max (L.) Merrill]. 3 Biotech 2019, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fang, Q.; Zheng, J.; Li, Z.; Li, Y.; Feng, Y.; Han, Y.; Li, Y. GmLecRlk, a lectin receptor-like protein kinase, contributes to salt stress tolerance by regulating salt-responsive genes in soybean. Int. J. Mol. Sci. 2022, 23, 1030. [Google Scholar] [CrossRef]
- Ma, L.; Liang, Z. CRISPR technology for abiotic stress resistant crop breeding. Plant Growth Regul. 2021, 94, 115–129. [Google Scholar] [CrossRef]
- Dongre, N.; Kumari, D.; Mahto, B.K.; Arya, S.S.; Lenka, S.K. Mutagenomics for Functional Analysis of Plant Genome using CRISPR Library Screen. In RNA-Based Technologies for Functional Genomics in Plants; Springer: Berlin/Heidelberg, Germany, 2021; pp. 339–367. [Google Scholar]
- Nazir, R.; Mandal, S.; Mitra, S.; Ghorai, M.; Das, N.; Jha, N.K.; Majumder, M.; Pandey, D.K.; Dey, A. Clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated genome-editing toolkit to enhance salt stress tolerance in rice and wheat. Physiol. Plant. 2022, 174, e13642. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.T.; Choi, M.; Bae, S.J.; Kim, J.S. The functional association of ACQOS/VICTR with salt stress resistance in Arabidopsis thaliana was confirmed by CRISPR-mediated mutagenesis. Int. J. Mol. Sci. 2021, 22, 11389. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.H.; Tariq, H.; Farooq, I.; Tasleeem, H.; Ghouri, M.Z.; Mubarik, M.S.; Khan, Z. Applications of CRISPR/Cas System in Plants. In The CRISPR/Cas Tool Kit for Genome Editing; Springer: Berlin/Heidelberg, Germany, 2022; pp. 285–309. [Google Scholar]
- Niu, F.; Jiang, Q.; Cheng, R.; Sun, X.; Hu, Z.; Wang, L.; Zhang, H. CRISPR/Cas9-mediated targeted mutagenesis of wild soybean (Glycine soja) hairy roots altered the transcription profile of the mutant. J. Agric. Sci. 2020, 12, 14–33. [Google Scholar] [CrossRef]
- Shi, W.Y.; Du, Y.T.; Ma, J.; Min, D.-H.; Jin, L.G.; Chen, J.; Chen, M.; Zhou, Y.B.; Ma, Y.Z.; Xu, Z.-S. The WRKY transcription factor GmWRKY12 confers drought and salt tolerance in soybean. Int. J. Mol Sci. 2018, 19, 4087. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, N.; Yang, J.; Guo, H.; Liu, Z.; Zheng, X.; Li, S.; Xiang, F. The salt-induced transcription factor GmMYB84 confers salinity tolerance in soybean. Plant Sci. 2020, 291, 110326. [Google Scholar] [CrossRef]
- Yu, Y.; Wang, N.; Hu, R.; Xiang, F. Genome-wide identification of soybean WRKY transcription factors in response to salt stress. Springerplus 2016, 5, 920. [Google Scholar] [CrossRef]
- Zhang, X.X.; Tang, Y.J.; Ma, Q.B.; Yang, C.Y.; Mu, Y.H.; Suo, H.C.; Luo, L.H.; Nian, H. OsDREB2A, a rice transcription factor, significantly affects salt tolerance in transgenic soybean. PLoS ONE 2013, 8, e83011. [Google Scholar] [CrossRef]
- Pi, E.; Xu, J.; Li, H.; Fan, W.; Zhu, C.; Zhang, T.; Jiang, J.; He, L.; Lu, H.; Wang, H. Enhanced salt tolerance of rhizobia-inoculated soybean correlates with decreased phosphorylation of the transcription factor GmMYB183 and altered flavonoid biosynthesis. Mol. Cell. Proteom. 2019, 18, 2225–2243. [Google Scholar] [CrossRef]
- Liu, X.; Pi, B.; Du, Z.; Yang, T.; Gu, M.; Sun, S.; Yu, B. The transcription factor GmbHLH3 confers Cl−/salt tolerance to soybean by upregulating GmCLC1 expression for maintenance of anion homeostasis. Environ. Exp. Bot. 2022, 194, 104755. [Google Scholar] [CrossRef]
- Li, B.; Liu, Y.; Cui, X.-Y.; Fu, J.D.; Zhou, Y.B.; Zheng, W.J.; Lan, J.-H.; Jin, L.G.; Chen, M.; Ma, Y.Z. Genome-wide characterization and expression analysis of soybean TGA transcription factors identified a novel TGA gene involved in drought and salt tolerance. Front Plant Sci. 2019, 10, 549. [Google Scholar] [CrossRef]
- Ming, L.; Zheng, H.; Jiang, Q.-y.; SUN, X.-j.; Yuan, G.; Qi, J.C.; Zhang, H. GmNAC15 overexpression in hairy roots enhances salt tolerance in soybean. J. Integr. Agric. 2018, 17, 530–538. [Google Scholar]
- Li, Y.; Chen, Q.; Nan, H.; Li, X.; Lu, S.; Zhao, X.; Liu, B.; Guo, C.; Kong, F.; Cao, D. Overexpression of GmFDL19 enhances tolerance to drought and salt stresses in soybean. PLoS ONE 2017, 12, e0179554. [Google Scholar] [CrossRef]
- Ke, D.; He, Y.; Fan, L.; Niu, R.; Cheng, L.; Wang, L.; Zhang, Z. The soybean TGA transcription factor GmTGA13 plays important roles in the response to salinity stress. Plant Biol. 2022, 24, 313–322. [Google Scholar] [CrossRef]
- Bian, X.H.; Li, W.; Niu, C.F.; Wei, W.; Hu, Y.; Han, J.Q.; Lu, X.; Tao, J.J.; Jin, M.; Qin, H. A class B heat shock factor selected for during soybean domestication contributes to salt tolerance by promoting flavonoid biosynthesis. New Phytol. 2020, 225, 268–283. [Google Scholar] [CrossRef]
- Nguyen, Q.H.; Vu, L.T.K.; Nguyen, L.T.N.; Pham, N.T.T.; Nguyen, Y.T.H.; Le, S.V.; Chu, M.H. Overexpression of the GmDREB6 gene enhances proline accumulation and salt tolerance in genetically modified soybean plants. Sci. Rep. 2019, 9, 19663. [Google Scholar] [CrossRef]
- Belamkar, V.; Weeks, N.T.; Bharti, A.K.; Farmer, A.D.; Graham, M.A.; Cannon, S.B. Comprehensive characterization and RNA-Seq profiling of the HD-Zip transcription factor family in soybean (Glycine max) during dehydration and salt stress. BMC Genom. 2014, 15, 950. [Google Scholar] [CrossRef]
- Liu, X.; Yang, X.; Zhang, B. Transcriptome analysis and functional identification of GmMYB46 in soybean seedlings under salt stress. PeerJ 2021, 9, e12492. [Google Scholar] [CrossRef] [PubMed]
- Price, A.L.; Patterson, N.J.; Plenge, R.M.; Weinblatt, M.E.; Shadick, N.A.; Reich, D. Principal components analysis corrects for stratification in genome-wide association studies. Nat. Genet. 2006, 38, 904–909. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Ersoz, E.; Lai, C.-Q.; Todhunter, R.J.; Tiwari, H.K.; Gore, M.A.; Bradbury, P.J.; Yu, J.; Arnett, D.K.; Ordovas, J.M. Mixed linear model approach adapted for genome-wide association studies. Nat. Genet. 2010, 42, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Segura, V.; Vilhjálmsson, B.J.; Platt, A.; Korte, A.; Seren, Ü.; Long, Q.; Nordborg, M. An efficient multi-locus mixed-model approach for genome-wide association studies in structured populations. Nat. Gen. 2012, 44, 825–830. [Google Scholar] [CrossRef]
- Wen, Z.; Boyse, J.F.; Song, Q.; Cregan, P.B.; Wang, D. Genomic consequences of selection and genome-wide association mapping in soybean. BMC Genom. 2015, 16, 671. [Google Scholar] [CrossRef]
- Zhang, J.; Song, Q.; Cregan, P.B.; Jiang, G.-L. Genome-wide association study, genomic prediction and marker-assisted selection for seed weight in soybean (Glycine max). Theor. Appl. Genet. 2016, 129, 117–130. [Google Scholar] [CrossRef]
- Wu, W.; Zucca, C.; Muhaimeed, A.S.; Al-Shafie, W.M.; Fadhil Al-Quraishi, A.M.; Nangia, V.; Zhu, M.; Liu, G. Soil salinity prediction and mapping by machine learning regression in C entral M esopotamia, I raq. Land Degrad. Dev. 2018, 29, 4005–4014. [Google Scholar] [CrossRef]
- Teodoro, P.E.; Teodoro, L.P.R.; Baio, F.H.R.; da Silva Junior, C.A.; dos Santos, R.G.; Ramos, A.P.M.; Pinheiro, M.M.F.; Osco, L.P.; Gonçalves, W.N.; Carneiro, A.M. Predicting days to maturity, plant height, and grain yield in soybean: A machine and deep learning approach using multispectral data. Remote Sens. 2021, 13, 4632. [Google Scholar] [CrossRef]
- Niazian, M.; Shariatpanahi, M.E.; Abdipour, M.; Oroojloo, M. Modeling callus induction and regeneration in an anther culture of tomato (Lycopersicon esculentum L.) using image processing and artificial neural network method. Protoplasma 2019, 256, 1317–1332. [Google Scholar] [CrossRef]
- Hesami, M.; Naderi, R.; Yoosefzadeh-Najafabadi, M.; Rahmati, M. Data-driven modeling in plant tissue culture. J. Appl. Environ. Biol. Sci 2017, 7, 37–44. [Google Scholar]
- Ravari, S.; Dehghani, H.; Naghavi, H. Assessment of salinity indices to identify Iranian wheat varieties using an artificial neural network. Ann. Appl. Biol. 2016, 168, 185–194. [Google Scholar] [CrossRef]




| Varieties/Mutants/Transgenic Lines | Breeding Tools | References |
|---|---|---|
| gmaitr36, gmaitr23456 mutants (Wm82 wild type background) | CRISPR/Cas9 | [61] |
| GmNHX5 mutants (Jidou-7 variety) | CRISPR/Cas9 | [62] |
| GmMYB118 mutants | CRISPR/Cas9 | [63] |
| NIL-J transgenic soybean line | Genetic engineering | [23] |
| PgTIP1-transgenic lines (hybrid cultivar) 4076 | Genetic engineering | [64] |
| cv. Liaodou 15 | Genetic engineering | [25] |
| Transgenic lines (T3) with TaNHX2 overexpression | Genetic engineering | [65] |
| Jackson (Ncl gene) | Genetic engineering | [59] |
| OX1, OX2, OX4 transgenic lines | Genetic engineering | [66] |
| Nannong 1138-2 | Conventional breeding | [32] |
| Tiefeng 8 | Conventional breeding | [52] |
| Baiqiu 1 | Conventional breeding | [57] |
| Fengzitianandou | Conventional breeding | [57] |
| Gene | Functions | References |
|---|---|---|
| GIGANTEA | Reduced the flowering time and increased salt tolerance | [88] |
| GsAKR1 | Enhanced salt tolerance during germination stage | [83] |
| GmDUF4228-70 | Promoted the expression of marker genes during salt stress | [84] |
| GmCHX | Exhibited large allelic variation and improved salt tolerance | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rasheed, A.; Raza, A.; Jie, H.; Mahmood, A.; Ma, Y.; Zhao, L.; Xing, H.; Li, L.; Hassan, M.U.; Qari, S.H.; et al. Molecular Tools and Their Applications in Developing Salt-Tolerant Soybean (Glycine max L.) Cultivars. Bioengineering 2022, 9, 495. https://doi.org/10.3390/bioengineering9100495
Rasheed A, Raza A, Jie H, Mahmood A, Ma Y, Zhao L, Xing H, Li L, Hassan MU, Qari SH, et al. Molecular Tools and Their Applications in Developing Salt-Tolerant Soybean (Glycine max L.) Cultivars. Bioengineering. 2022; 9(10):495. https://doi.org/10.3390/bioengineering9100495
Chicago/Turabian StyleRasheed, Adnan, Ali Raza, Hongdong Jie, Athar Mahmood, Yushen Ma, Long Zhao, Hucheng Xing, Linlin Li, Muhammad Umair Hassan, Sameer H. Qari, and et al. 2022. "Molecular Tools and Their Applications in Developing Salt-Tolerant Soybean (Glycine max L.) Cultivars" Bioengineering 9, no. 10: 495. https://doi.org/10.3390/bioengineering9100495
APA StyleRasheed, A., Raza, A., Jie, H., Mahmood, A., Ma, Y., Zhao, L., Xing, H., Li, L., Hassan, M. U., Qari, S. H., & Jie, Y. (2022). Molecular Tools and Their Applications in Developing Salt-Tolerant Soybean (Glycine max L.) Cultivars. Bioengineering, 9(10), 495. https://doi.org/10.3390/bioengineering9100495