
3D Hepatic Organoid-Based Advancements in LIVER Tissue Engineering



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
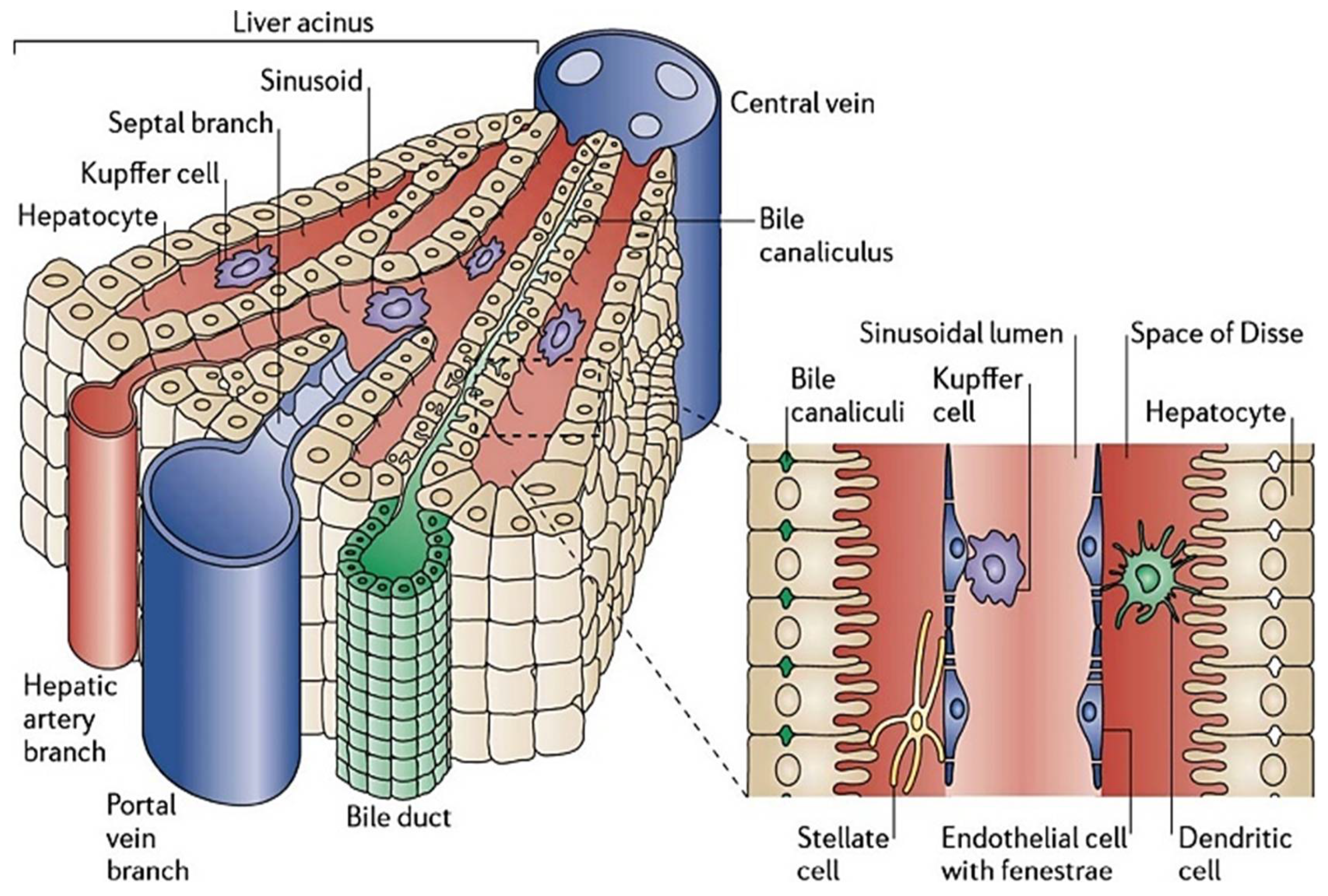
2. Physiological Microenvironment of the Liver
2.1. Liver Physiological Units
2.1.1. Liver Lobule
2.1.2. Liver Sinusoid (LSs)
2.2. Cell Types and Composition
2.2.1. Parenchymal Cells/Hepatocytes (HEPs)
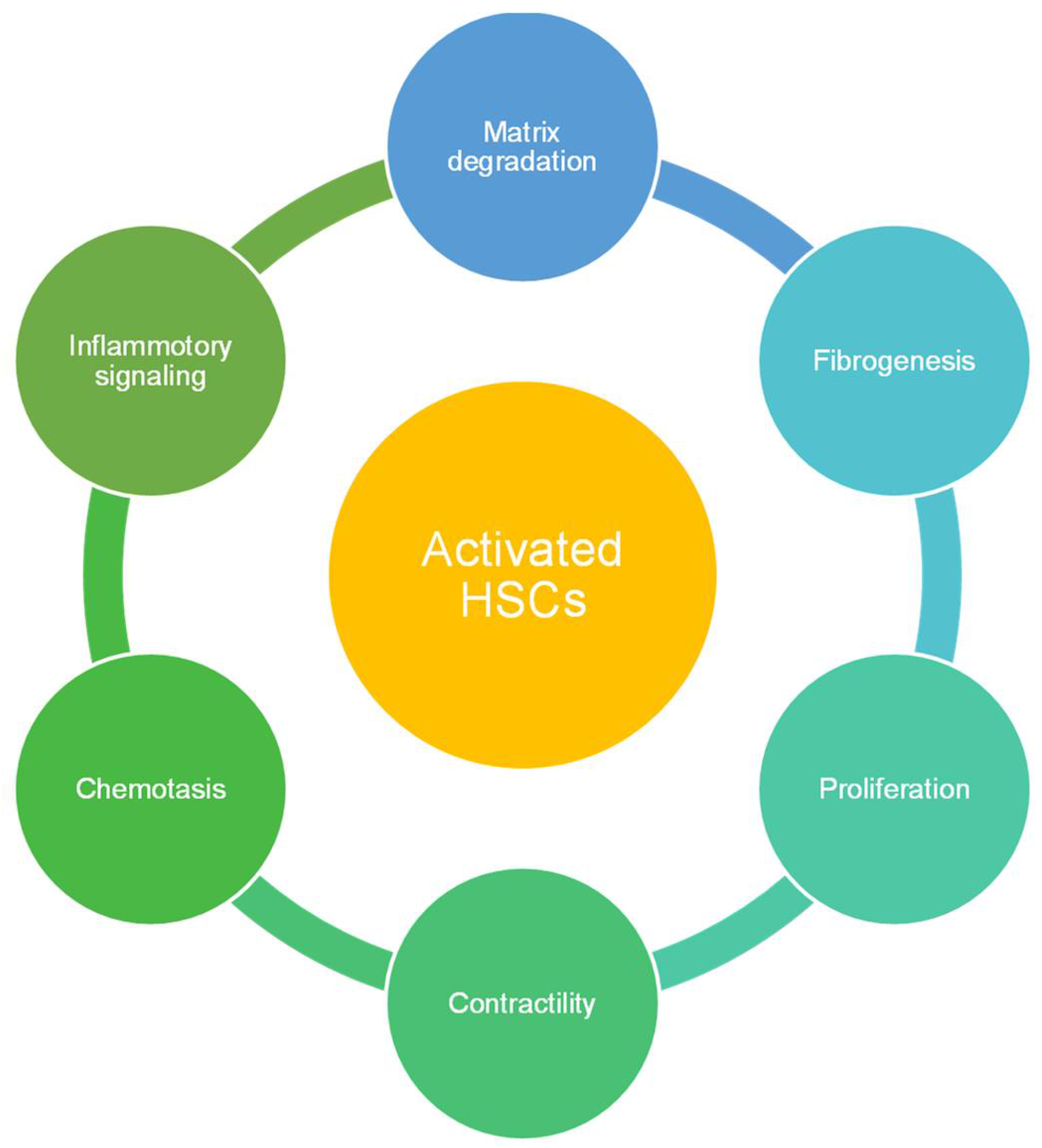
2.2.2. Hepatic Stellate Cells (HSCs)
2.2.3. Hepatic/Liver Sinusoidal Endothelial Cells (LSECs)
2.2.4. Kupffer Cells (KCs)
2.2.5. Biliary Epithelial Cells/Cholangiocytes (BECs)
3. Development of Hepatic Tissues: Strategies and Challenges
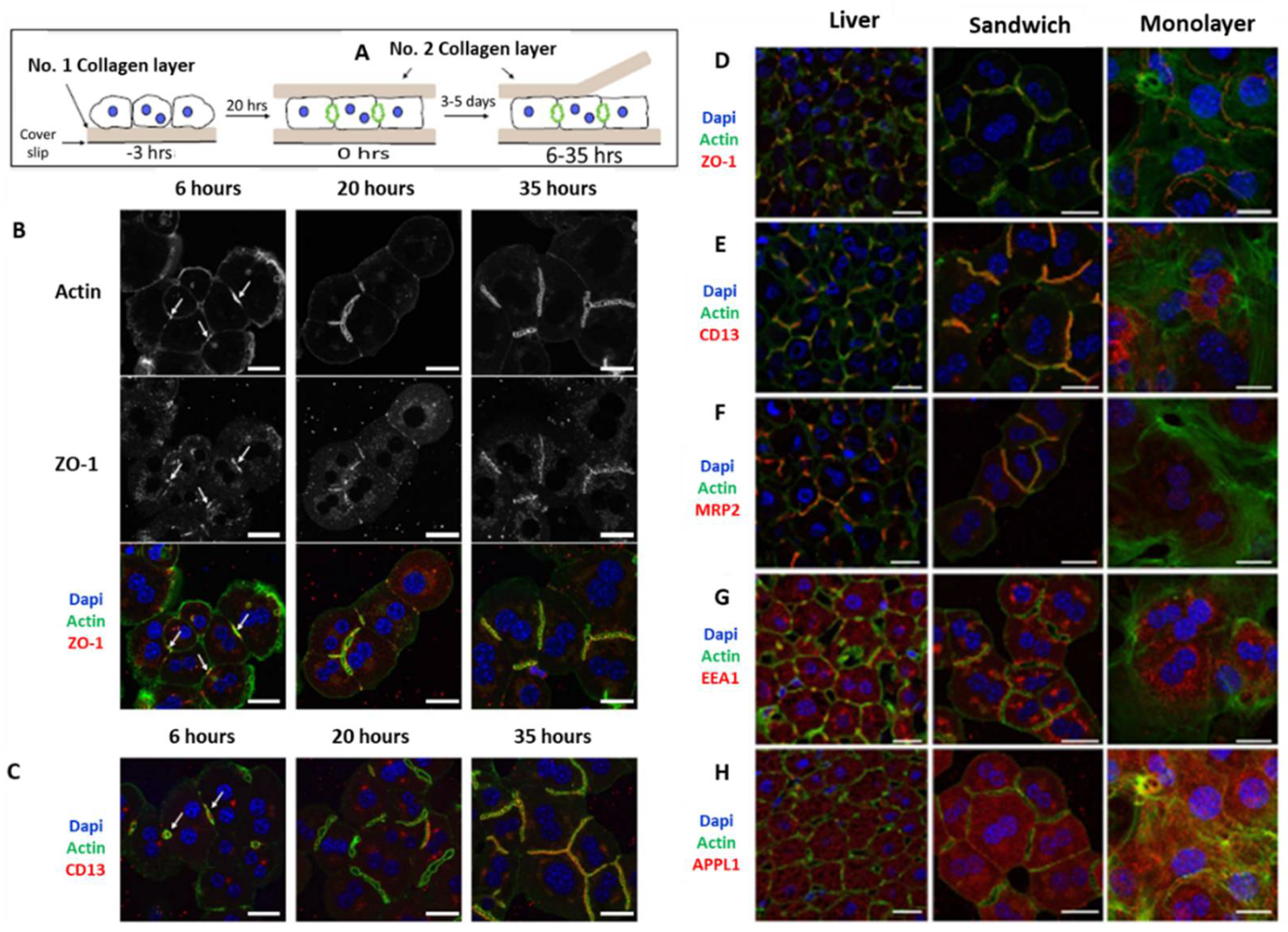
3.1. 2D Monolayer Culture
3.2. Sandwich Culture
3.3. Scaffold Free Spheroid/Organoid Development
3.4. Liver on a Chip
4. Application of Hepatic Organoids
4.1. Drug Screening
4.2. Disease Model
4.3. Metabolism Prediction
4.4. Multi Organ-on-a-Chip
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Asrani, S.K.; Devarbhavi, H.; Eaton, J.; Kamath, P.S. Burden of liver diseases in the world. J. Hepatol. 2019, 70, 151–171. [Google Scholar] [CrossRef]
- Fernandez-Checa, J.C.; Bagnaninchi, P.; Ye, H.; Sancho-Bru, P.; Falcon-Perez, J.M.; Royo, F.; Garcia-Ruiz, C.; Konu, O.; Miranda, J.; Lunov, O.; et al. Advanced preclinical models for evaluation of drug-induced liver injury-consensus statement by the European Drug-Induced Liver Injury Network [PRO-EURO-DILI-NET]. J. Hepatol. 2021, 75, 235–959. [Google Scholar] [CrossRef]
- Soldatow, V.Y.; Lecluyse, E.L.; Griffith, L.G.; Rusyn, I. In vitro models for liver toxicity testing. Toxicol. Res. (Camb) 2013, 2, 23–39. [Google Scholar] [CrossRef]
- Velasco, V.; Shariati, S.A.; Esfandyarpour, R. Microtechnology-based methods for organoid models. Microsyst. Nanoeng. 2020, 6, 76. [Google Scholar] [CrossRef]
- Du, Y.; Li, N.; Long, M. Chapter 6-Liver sinusoid on a chip. In Methods in Cell Biology; Doh, J., Fletcher, D., Piel, M., Eds.; Academic Press: Cambridge, MA, USA, 2018; Volume 146, pp. 105–134. [Google Scholar]
- Usta, O.; McCarty, W.; Bale, S.; Hegde, M.; Jindal, R.; Bhushan, A.; Golberg, I.; Yarmush, M. Microengineered cell and tissue systems for drug screening and toxicology applications: Evolution of in-vitro liver technologies. Technology 2015, 3, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Godoy, P.; Hewitt, N.J.; Albrecht, U.; Andersen, M.E.; Ansari, N.; Bhattacharya, S.; Bode, J.G.; Bolleyn, J.; Borner, C.; Boettger, J. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch. Toxicol. 2013, 87, 1315–1530. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.L.; Soroka, C.J. Bile formation and Secretion: An update. J. Hepatol. 2021, 3, 1035–1078. [Google Scholar] [CrossRef] [PubMed]
- Schulze, R.J.; Schott, M.B.; Casey, C.A.; Tuma, P.L.; McNiven, M.A. The cell biology of the hepatocyte: A membrane trafficking machine. J. Cell Biol. 2019, 218, 2096–2112. [Google Scholar] [CrossRef]
- Hendriks, H.F.; Blaner, W.S.; Wennekers, H.M.; Piantedosi, R.; Brouwer, A.; de Leeuw, A.M.; Goodman, D.S.; Knook, D.L. Distributions of retinoids, retinoid-binding proteins and related parameters in different types of liver cells isolated from young and old rats. Eur. J. Biochem. 1988, 171, 237–244. [Google Scholar] [CrossRef]
- Hendriks, H.; Verhoofstad, W.; Brouwer, A.; De Leeuw, A.; Knook, D. Perisinusoidal fat-storing cells are the main vitamin A storage sites in rat liver. Exp. Cell Res. 1985, 160, 138–149. [Google Scholar] [CrossRef]
- Ramadori, G.; Knittel, T.; Schwögler, S.; Bieber, F.; Rieder, H.; Büschenfelde, K.-H.M.Z. Dexamethasone modulates α2-macroglobulin and apolipoprotein E gene expression in cultured rat liver fat-storing (Ito) cells. Hepatology 1991, 14, 875–882. [Google Scholar] [CrossRef]
- Breitkopf, K.; Godoy, P.; Ciuclan, L.; Singer, M.; Dooley, S. TGF-β/Smad signaling in the injured liver. Z. für Gastroenterol. 2006, 44, 57–66. [Google Scholar] [CrossRef]
- Paik, Y.H.; Schwabe, R.F.; Bataller, R.; Russo, M.P.; Jobin, C.; Brenner, D.A. Toll-like receptor 4 mediates inflammatory signaling by bacterial lipopolysaccharide in human hepatic stellate cells. Hepatology 2003, 37, 1043–1055. [Google Scholar] [CrossRef]
- Fimmel, C.J.; Brown, K.E.; O’Neill, R.; Kladney, R.D. Complement C4 protein expression by rat hepatic stellate cells. J. Immunol. 1996, 157, 2601–2609. [Google Scholar]
- Marra, F.; Pinzani, M. Role of hepatic stellate cells in the pathogenesis of portal hypertension. Nefrologia 2002, 22, 34–40. [Google Scholar]
- Bonacchi, A.; Petrai, I.; Defranco, R.M.; Lazzeri, E.; Annunziato, F.; Efsen, E.; Cosmi, L.; Romagnani, P.; Milani, S.; Failli, P. The chemokine CCL21 modulates lymphocyte recruitment and fibrosis in chronic hepatitis C. Gastroenterology 2003, 125, 1060–1076. [Google Scholar] [CrossRef]
- Brun, P.; Castagliuolo, I.; Pinzani, M.; Palù, G.; Martines, D. Exposure to bacterial cell wall products triggers an inflammatory phenotype in hepatic stellate cells. Am. J. Physiol.-Gastrointest. Liver Physiol. 2005, 289, G571–G578. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Laskin, D.L. Nonparenchymal cells and hepatotoxicity. Semin. Liver Dis. 1990, 10, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Yokomori, H.; Oda, M.; Ogi, M.; Sakai, K.; Ishii, H. Enhanced expression of endothelial nitric oxide synthase and caveolin-1 in human cirrhosis. Liver 2002, 22, 150–158. [Google Scholar] [CrossRef]
- Gracia-Sancho, J.; Caparrós, E.; Fernández-Iglesias, A.; Francés, R. Role of liver sinusoidal endothelial cells in liver diseases. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 411–431. [Google Scholar] [CrossRef] [PubMed]
- Poisson, J.; Lemoinne, S.; Boulanger, C.; Durand, F.; Moreau, R.; Valla, D.; Rautou, P.-E. Liver sinusoidal endothelial cells: Physiology and role in liver diseases. J. Hepatol. 2017, 66, 212–227. [Google Scholar] [CrossRef] [PubMed]
- DeLeve, L.D. Liver sinusoidal endothelial cells in hepatic fibrosis. Hepatology 2015, 61, 1740–1746. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, A.L.; Qurashi, M.; Shetty, S. The Role of Sinusoidal Endothelial Cells in the Axis of Inflammation and Cancer Within the Liver. Front. Physiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Dixon, L.J.; Barnes, M.; Tang, H.; Pritchard, M.T.; Nagy, L.E. Kupffer cells in the liver. Compr. Physiol. 2013, 3, 785. [Google Scholar]
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of Kupffer cells in the pathogenesis of liver disease. World J. Gastroenterol. WJG 2006, 12, 7413. [Google Scholar] [CrossRef] [PubMed]
- Bilzer, M.; Roggel, F.; Gerbes, A.L. Role of Kupffer cells in host defense and liver disease. Liver Int. 2006, 26, 1175–1186. [Google Scholar] [CrossRef]
- Sirica, A.E.; Mathis, G.A.; Sano, N.; Elmore, L.W. Isolation, culture, and transplantation of intrahepatic biliary epithelial cells and oval cells. Pathobiology 1990, 58, 44–64. [Google Scholar] [CrossRef]
- Alison, M.R.; Golding, M.; Sarraf, C.E.; Edwards, R.J.; Lalani, E.-N. Liver damage in the rat induces hepatocyte stem cells from biliary epithelial cells. Gastroenterology 1996, 110, 1182–1190. [Google Scholar] [CrossRef]
- Deng, J.; Zhang, X.; Chen, Z.; Luo, Y.; Lu, Y.; Liu, T.; Wu, Z.; Jin, Y.; Zhao, W.; Lin, B. A cell lines derived microfluidic liver model for investigation of hepatotoxicity induced by drug-drug interaction. Biomicrofluidics 2019, 13, 024101. [Google Scholar] [CrossRef]
- Milner, E.; Ainsworth, M.; McDonough, M.; Stevens, B.; Buehrer, J.; Delzell, R.; Wilson, C.; Barnhill, J. Emerging Three-Dimensional Hepatic Models in Relation to Traditional Two-Dimensional In Vitro Assays for Evaluating Drug Metabolism and Hepatoxicity. Med. Drug Discov. 2020, 8, 100060. [Google Scholar] [CrossRef]
- Aday, S.; Hasirci, N.; Deliloglu Gurhan, I. A cost-effective and simple culture method for primary hepatocytes. Anim. Cells Syst. 2011, 15, 19–27. [Google Scholar] [CrossRef]
- Lin, P.; Chan, W.C.; Badylak, S.F.; Bhatia, S.N. Assessing porcine liver-derived biomatrix for hepatic tissue engineering. Tissue Eng. 2004, 10, 1046–1053. [Google Scholar] [CrossRef]
- Natarajan, V.; Berglund, E.J.; Chen, D.X.; Kidambi, S. Substrate stiffness regulates primary hepatocyte functions. RSC Adv. 2015, 5, 80956–80966. [Google Scholar] [CrossRef]
- Foucher, J.; Chanteloup, E.; Vergniol, J.; Castéra, L.; Le Bail, B.; Adhoute, X.; Bertet, J.; Couzigou, P.; de Lédinghen, V. Diagnosis of cirrhosis by transient elastography (FibroScan): A prospective study. Gut 2006, 55, 403–408. [Google Scholar] [CrossRef]
- Takebe, T.; Sekine, K.; Enomura, M.; Koike, H.; Kimura, M.; Ogaeri, T.; Zhang, R.-R.; Ueno, Y.; Zheng, Y.-W.; Koike, N. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature 2013, 499, 481–484. [Google Scholar] [CrossRef]
- Dunn, J.C.Y.; Yarmush, M.L.; Koebe, H.G.; Tompkins, R.G. Hepatocyte function and extracellular matrix geometry: Long-term culture in a sandwich configuration. FASEB J. 1989, 3, 174–177. [Google Scholar] [CrossRef] [PubMed]
- Keemink, J.; Oorts, M.; Annaert, P. Primary hepatocytes in sandwich culture. In Protocols in In Vitro Hepatocyte Research; Springer: Berlin/Heidelberg, Germany, 2015; pp. 175–188. [Google Scholar]
- Treijtel, N.; Barendregt, A.; Freidig, A.P.; Blaauboer, B.J.; van Eijkeren, J.C. Modeling the in vitro intrinsic clearance of the slowly metabolized compound tolbutamide determined in sandwich-cultured rat hepatocytes. Drug Metab. Dispos. 2004, 32, 884–891. [Google Scholar] [CrossRef] [PubMed]
- Zeigerer, A.; Wuttke, A.; Marsico, G.; Seifert, S.; Kalaidzidis, Y.; Zerial, M. Functional properties of hepatocytes in vitro are correlated with cell polarity maintenance. Exp. Cell Res. 2017, 350, 242–252. [Google Scholar] [CrossRef]
- Turncliff, R.Z.; Tian, X.; Brouwer, K.L.R. Effect of culture conditions on the expression and function of Bsep, Mrp2, and Mdr1a/b in sandwich-cultured rat hepatocytes. Biochem. Pharmacol. 2006, 71, 1520–1529. [Google Scholar] [CrossRef] [PubMed]
- Swift*, B.; Pfeifer*, N.D.; Brouwer, K.L.R. Sandwich-cultured hepatocytes: An in vitro model to evaluate hepatobiliary transporter-based drug interactions and hepatotoxicity. Drug Metab. Rev. 2010, 42, 446–471. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Toprakhisar, B.; Van Haele, M.; Antoranz, A.; Boon, R.; Chesnais, F.; De Smedt, J.; Tricot, T.; Idoype, T.I.; Canella, M. A fully defined matrix to support a pluripotent stem cell derived multi-cell-liver steatohepatitis and fibrosis model. Biomaterials 2021, 276, 121006. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.-L.; Kim, Y.-K.; Cho, K.-H.; Jang, Y.-C.; Choi, Y.-J.; Chung, J.-H.; Cho, C.-S. Roles of spheroid formation of hepatocytes in liver tissue engineering. Int. J. Stem Cells 2010, 3, 69–73. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bell, C.C.; Hendriks, D.F.G.; Moro, S.M.L.; Ellis, E.; Walsh, J.; Renblom, A.; Fredriksson Puigvert, L.; Dankers, A.C.A.; Jacobs, F.; Snoeys, J.; et al. Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease. Sci. Rep. 2016, 6, 25187. [Google Scholar] [CrossRef]
- Hendriks, D.F.; Fredriksson Puigvert, L.; Messner, S.; Mortiz, W.; Ingelman-Sundberg, M. Hepatic 3D spheroid models for the detection and study of compounds with cholestatic liability. Sci. Rep. 2016, 6, 35434. [Google Scholar] [CrossRef]
- D’Souza, A.A.; Devarajan, P.V. Asialoglycoprotein receptor mediated hepatocyte targeting-strategies and applications. J. Control. Release 2015, 203, 126–139. [Google Scholar] [CrossRef]
- Kobayashi, A.; Kobayashi, K.; Akaike, T. Control of adhesion and detachment of parenchymal liver cells using lactose-carrying polystyrene as substratum. J. Biomater. Sci. Polym. Ed. 1992, 3, 499–508. [Google Scholar] [CrossRef]
- Tobe, S.; Takei, Y.; Kobayashi, K.; Akaike, T. Receptor-mediated formation of multilayer aggregates of primary cultured adult rat hepatocytes on lactose-substituted polystyrene. Biochem. Biophys. Res. Commun. 1992, 184, 225–230. [Google Scholar] [CrossRef]
- Shri, M.; Agrawal, H.; Rani, P.; Singh, D.; Onteru, S.K. Hanging Drop, A Best Three-Dimensional (3D) Culture Method for Primary Buffalo and Sheep Hepatocytes. Sci. Rep. 2017, 7, 1203. [Google Scholar] [CrossRef]
- Cho, C.-Y.; Chiang, T.-H.; Hsieh, L.-H.; Yang, W.-Y.; Hsu, H.-H.; Yeh, C.-K.; Huang, C.-C.; Huang, J.-H. Development of a Novel Hanging Drop Platform for Engineering Controllable 3D Microenvironments. Front. Cell Dev. Biol. 2020, 8, 327. [Google Scholar] [CrossRef]
- Puryear Iii, J.R.; Yoon, J.-K.; Kim, Y. Advanced Fabrication Techniques of Microengineered Physiological Systems. Micromachines 2020, 11, 730. [Google Scholar] [CrossRef] [PubMed]
- Bhise, N.S.; Manoharan, V.; Massa, S.; Tamayol, A.; Ghaderi, M.; Miscuglio, M.; Lang, Q.; Shrike Zhang, Y.; Shin, S.R.; Calzone, G.; et al. A liver-on-a-chip platform with bioprinted hepatic spheroids. Biofabrication 2016, 8, 014101. [Google Scholar] [CrossRef] [PubMed]
- Grant, J.; Özkan, A.; Oh, C.; Mahajan, G.; Prantil-Baun, R.; Ingber, D.E. Simulating drug concentrations in PDMS microfluidic organ chips. Lab Chip 2021, 21, 3509–3519. [Google Scholar] [CrossRef]
- Rennert, K.; Steinborn, S.; Gröger, M.; Ungerböck, B.; Jank, A.-M.; Ehgartner, J.; Nietzsche, S.; Dinger, J.; Kiehntopf, M.; Funke, H.; et al. A microfluidically perfused three dimensional human liver model. Biomaterials 2015, 71, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Broutier, L.; Mastrogiovanni, G.; Verstegen, M.M.A.; Francies, H.E.; Gavarró, L.M.; Bradshaw, C.R.; Allen, G.E.; Arnes-Benito, R.; Sidorova, O.; Gaspersz, M.P.; et al. Human primary liver cancer–derived organoid cultures for disease modeling and drug screening. Nat. Med. 2017, 23, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- Bartfeld, S.; Bayram, T.; van de Wetering, M.; Huch, M.; Begthel, H.; Kujala, P.; Vries, R.; Peters, P.J.; Clevers, H. In vitro expansion of human gastric epithelial stem cells and their responses to bacterial infection. Gastroenterology 2015, 148, 126–136. e126. [Google Scholar] [CrossRef]
- Au, S.H.; Chamberlain, M.D.; Mahesh, S.; Sefton, M.V.; Wheeler, A.R. Hepatic organoids for microfluidic drug screening. Lab Chip 2014, 14, 3290–3299. [Google Scholar] [CrossRef]
- Zhu, X.; Zhang, B.; He, Y.; Bao, J. Liver Organoids: Formation Strategies and Biomedical Applications. Tissue Eng. Regen. Med. 2021, 18, 573–585. [Google Scholar] [CrossRef]
- Lee, J.; Choi, B.; No, D.Y.; Lee, G.; Lee, S.-r.; Oh, H.; Lee, S.-H. A 3D alcoholic liver disease model on a chip. Integr. Biol. 2016, 8, 302–308. [Google Scholar] [CrossRef]
- Bhatia, S.N.; Underhill, G.H.; Zaret, K.S.; Fox, I.J. Cell and tissue engineering for liver disease. Sci. Transl. Med. 2014, 6, 245sr242. [Google Scholar] [CrossRef]
- Lee, G.H.; Lee, J.S.; Wang, X.; Hoon Lee, S. Bottom-Up Engineering of Well-Defined 3D Microtissues Using Microplatforms and Biomedical Applications. Adv. Healthc. Mater. 2016, 5, 56–74. [Google Scholar] [CrossRef] [PubMed]
- Yoon No, D.; Lee, K.-H.; Lee, J.; Lee, S.-H. 3D liver models on a microplatform: Well-defined culture, engineering of liver tissue and liver-on-a-chip. Lab Chip 2015, 15, 3822–3837. [Google Scholar] [CrossRef]
- Wang, S.; Wang, X.; Tan, Z.; Su, Y.; Liu, J.; Chang, M.; Yan, F.; Chen, J.; Chen, T.; Li, C.; et al. Human ESC-derived expandable hepatic organoids enable therapeutic liver repopulation and pathophysiological modeling of alcoholic liver injury. Cell Res. 2019, 29, 1009–1026. [Google Scholar] [CrossRef]
- Deng, J.; Chen, Z.; Zhang, X.; Luo, Y.; Wu, Z.; Lu, Y.; Liu, T.; Zhao, W.; Lin, B. A liver-chip-based alcoholic liver disease model featuring multi-non-parenchymal cells. Biomed. Microdevices 2019, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Teng, Y.; Zhao, Z.; Tasnim, F.; Huang, X.; Yu, H. A scalable and sensitive steatosis chip with long-term perfusion of in situ differentiated HepaRG organoids. Biomaterials 2021, 275, 120904. [Google Scholar] [CrossRef] [PubMed]
- Elbadawy, M.; Yamanaka, M.; Goto, Y.; Hayashi, K.; Tsunedomi, R.; Hazama, S.; Nagano, H.; Yoshida, T.; Shibutani, M.; Ichikawa, R. Efficacy of primary liver organoid culture from different stages of non-alcoholic steatohepatitis (NASH) mouse model. Biomaterials 2020, 237, 119823. [Google Scholar] [CrossRef]
- Ortega-Prieto, A.M.; Skelton, J.K.; Wai, S.N.; Large, E.; Lussignol, M.; Vizcay-Barrena, G.; Hughes, D.; Fleck, R.A.; Thursz, M.; Catanese, M.T.; et al. 3D microfluidic liver cultures as a physiological preclinical tool for hepatitis B virus infection. Nat. Commun. 2018, 9, 682. [Google Scholar] [CrossRef] [PubMed]
- Weaver, R.J.; Jochemsen, R. Nonclinical pharmacokinetics and toxicokinetics. In International Pharmaceutical Product Registration; CRC Press: Boca Raton, FL, USA, 2016; pp. 352–392. [Google Scholar]
- Williams, J.A.; Hyland, R.; Jones, B.C.; Smith, D.A.; Hurst, S.; Goosen, T.C.; Peterkin, V.; Koup, J.R.; Ball, S.E. Drug-drug interactions for UDP-glucuronosyltransferase substrates: A pharmacokinetic explanation for typically observed low exposure (AUCI/AUC) ratios. Drug Metab. Dispos. 2004, 32, 1201. [Google Scholar] [CrossRef]
- Fasinu, P.; Bouic, P.J.; Rosenkranz, B. Liver-based in vitro technologies for drug biotransformation studies-a review. Curr. Drug Metab. 2012, 13, 215–224. [Google Scholar] [CrossRef]
- Hanioka, N.; Saito, K.; Isobe, T.; Ohkawara, S.; Jinno, H.; Tanaka-Kagawa, T. Favipiravir biotransformation in liver cytosol: Species and sex differences in humans, monkeys, rats, and mice. Biopharm. Drug Dispos. 2021, 42, 218–225. [Google Scholar] [CrossRef]
- Liu, J.; Lu, Y.-F.; Corton, J.C.; Klaassen, C.D. Expression of cytochrome P450 isozyme transcripts and activities in human livers. Xenobiotica 2021, 51, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Riede, J.; Wollmann, B.M.; Molden, E.; Ingelman-Sundberg, M. Primary Human Hepatocyte Spheroids as an In Vitro Tool for Investigating Drug Compounds with Low Hepatic Clearance. Drug Metab. Dispos. 2021, 49, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Ouchi, R.; Togo, S.; Kimura, M.; Shinozawa, T.; Koido, M.; Koike, H.; Thompson, W.; Karns, R.A.; Mayhew, C.N.; McGrath, P.S.; et al. Modeling Steatohepatitis in Humans with Pluripotent Stem Cell-Derived Organoids. Cell Metab. 2019, 30, 374–384.e376. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Patel, D.; Kwa, T.; Haque, A.; Matharu, Z.; Stybayeva, G.; Gao, Y.; Diehl, A.M.; Revzin, A. Liver injury-on-a-chip: Microfluidic co-cultures with integrated biosensors for monitoring liver cell signaling during injury. Lab Chip 2015, 15, 4467–4478. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Sung, J.H. Gut-liver on a chip toward an in vitro model of hepatic steatosis. Biotechnol. Bioeng. 2018, 115, 2817–2827. [Google Scholar] [CrossRef]
- Jeon, J.w.; Lee, S.H.; Kim, D.; Sung, J.H. In vitro hepatic steatosis model based on gut–liver-on-a-chip. Biotechnol. Prog. 2021, 37, e3121. [Google Scholar] [CrossRef]
- Esch, M.B.; Mahler, G.J.; Stokol, T.; Shuler, M.L. Body-on-a-chip simulation with gastrointestinal tract and liver tissues suggests that ingested nanoparticles have the potential to cause liver injury. Lab Chip 2014, 14, 3081–3092. [Google Scholar] [CrossRef]
- Yin, F.; Zhang, X.; Wang, L.; Wang, Y.; Zhu, Y.; Li, Z.; Tao, T.; Chen, W.; Yu, H.; Qin, J. HiPSC-derived multi-organoids-on-chip system for safety assessment of antidepressant drugs. Lab Chip 2021, 21, 571–581. [Google Scholar] [CrossRef]
- Schimek, K.; Frentzel, S.; Luettich, K.; Bovard, D.; Rütschle, I.; Boden, L.; Rambo, F.; Erfurth, H.; Dehne, E.-M.; Winter, A.; et al. Human multi-organ chip co-culture of bronchial lung culture and liver spheroids for substance exposure studies. Sci. Rep. 2020, 10, 7865. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panwar, A.; Das, P.; Tan, L.P. 3D Hepatic Organoid-Based Advancements in LIVER Tissue Engineering. Bioengineering 2021, 8, 185. https://doi.org/10.3390/bioengineering8110185
Panwar A, Das P, Tan LP. 3D Hepatic Organoid-Based Advancements in LIVER Tissue Engineering. Bioengineering. 2021; 8(11):185. https://doi.org/10.3390/bioengineering8110185
Chicago/Turabian StylePanwar, Amit, Prativa Das, and Lay Poh Tan. 2021. "3D Hepatic Organoid-Based Advancements in LIVER Tissue Engineering" Bioengineering 8, no. 11: 185. https://doi.org/10.3390/bioengineering8110185
APA StylePanwar, A., Das, P., & Tan, L. P. (2021). 3D Hepatic Organoid-Based Advancements in LIVER Tissue Engineering. Bioengineering, 8(11), 185. https://doi.org/10.3390/bioengineering8110185