
Effect on Osteogenic Differentiation of Genetically Modified IL4 or PDGF-BB Over-Expressing and IL4-PDGF-BB Co-Over-Expressing Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro

 , ,
, ,  , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
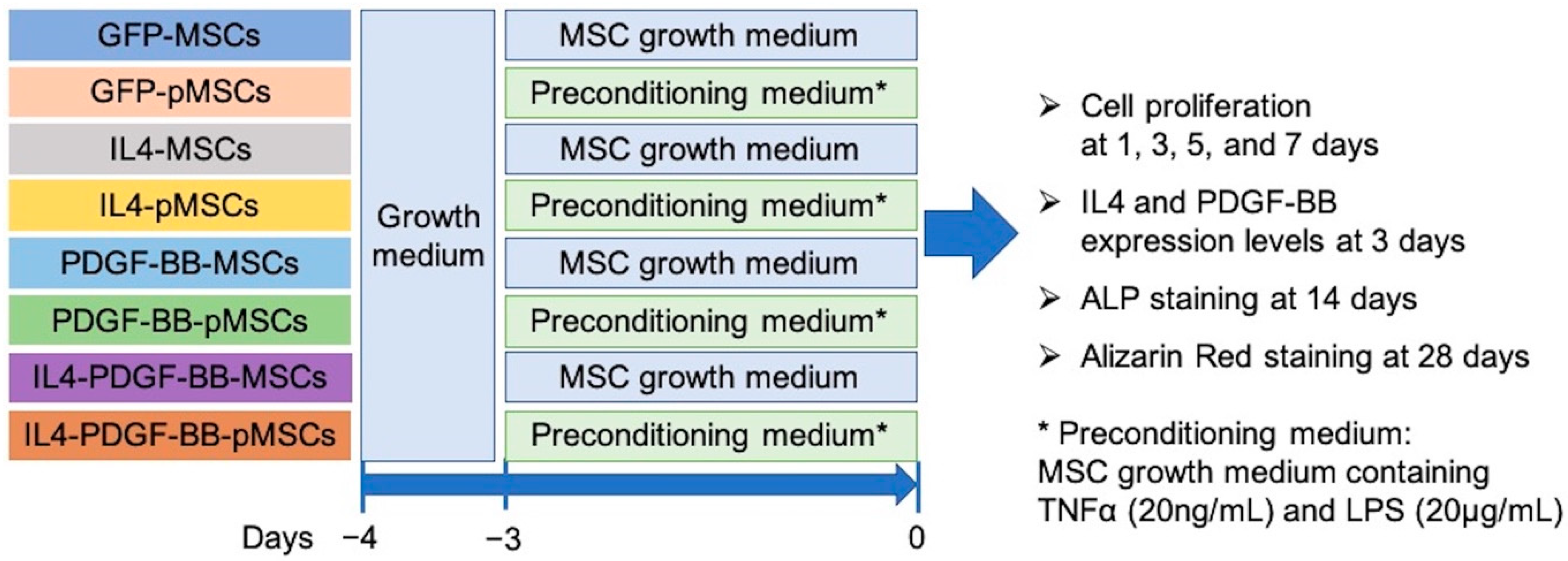
2.1. Rabbit Bone Marrow-Derived MSCs
2.2. Construction of Plasmids
2.3. Establishment of Genetically Modified Control MSCs (GFP-MSCs), IL4-MSCs, and PDGF-BB-MSCs
2.4. Establishment of IL4-PDGF-BB-MSCs
2.5. Cell Preparation for In Vitro Experiments
2.6. Cell Proliferation
2.7. IL4 and PDGF-BB Expression Levels
2.8. Osteogenic Differentiation
2.9. Statistical Analysis
3. Results
3.1. Cell Proliferation
3.2. IL4 and PDGF-BB Expression Levels
3.3. Osteogenic Differentiation
3.3.1. ALP Staining
3.3.2. Alizarin Red Staining
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mont, M.A.; Salem, H.S.; Piuzzi, N.S.; Goodman, S.B.; Jones, L.C. Nontraumatic Osteonecrosis of the Femoral Head: Where Do We Stand Today?: A 5-Year Update. J. Bone Joint Surg. Am. 2020, 102, 1084–1099. [Google Scholar] [CrossRef]
- Zhu, S.; Zhang, X.; Chen, X.; Wang, Y.; Li, S.; Qian, W. Comparison of cell therapy and other novel adjunctive therapies combined with core decompression for the treatment of osteonecrosis of the femoral head. Bone Joint Res. 2021, 10, 445–458. [Google Scholar] [CrossRef]
- Assouline-Dayan, Y.; Chang, C.; Greenspan, A.; Shoenfeld, Y.; Gershwin, M.E. Pathogenesis and natural history of osteonecrosis. Semin. Arthritis Rheum. 2002, 32, 94–124. [Google Scholar] [CrossRef]
- Rajpura, A.; Wright, A.C.; Board, T.N. Medical management of osteonecrosis of the hip: A review. Hip Int. J. Clin. Exp. Res. hip Pathol. Ther. 2011, 21, 385–392. [Google Scholar] [CrossRef]
- Cao, H.; Guan, H.; Lai, Y.; Qin, L.; Wang, X. Review of various treatment options and potential therapies for osteonecrosis of the femoral head. J. Orthop. Transl. 2016, 4, 57–70. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Lin, T.; Pan, C.-C.; Moeinzadeh, S.; Takagi, M.; Yang, Y.P.; Goodman, S.B. Cell-Based and Scaffold-Based Therapies for Joint Preservation in Early-Stage Osteonecrosis of the Femoral Head: A Review of Basic Research. JBJS Rev. 2019, 7, e5. [Google Scholar] [CrossRef]
- Gangji, V.; Hauzeur, J.P.; Schoutens, A.; Hinsenkamp, M.; Appelboom, T.; Egrise, D. Abnormalities in the replicative capacity of osteoblastic cells in the proximal femur of patients with osteonecrosis of the femoral head. J. Rheumatol. 2003, 30, 348–351. [Google Scholar]
- Mont, M.A.; Hungerford, D.S. Non-traumatic avascular necrosis of the femoral head. J. Bone Joint Surg. Am. 1995, 77, 459–474. [Google Scholar] [CrossRef]
- Huang, C.; Dai, J.; Zhang, X.A. Environmental physical cues determine the lineage specification of mesenchymal stem cells. Biochim. Biophys. Acta - Gen. Subj. 2015, 1850, 1261–1266. [Google Scholar] [CrossRef] [PubMed]
- Fulda, S.; Gorman, A.M.; Hori, O.; Samali, A. Cellular stress responses: Cell survival and cell death. Int. J. Cell Biol. 2010, 2010, 214074. [Google Scholar] [CrossRef] [PubMed]
- Haider, H.K.; Ashraf, M. Preconditioning and stem cell survival. J. Cardiovasc. Transl. Res. 2010, 3, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Pajarinen, J.; Nabeshima, A.; Lu, L.; Nathan, K.; Jämsen, E.; Yao, Z.; Goodman, S.B. Preconditioning of murine mesenchymal stem cells synergistically enhanced immunomodulation and osteogenesis. Stem Cell Res. Ther. 2017, 8, 277. [Google Scholar] [CrossRef]
- Diomede, F.; Gugliandolo, A.; Scionti, D.; Merciaro, I.; Cavalcanti, M.F.; Mazzon, E.; Trubiani, O. Biotherapeutic Effect of Gingival Stem Cells Conditioned Medium in Bone Tissue Restoration. Int. J. Mol. Sci. 2018, 19, 329. [Google Scholar] [CrossRef]
- Diomede, F.; D’Aurora, M.; Gugliandolo, A.; Merciaro, I.; Orsini, T.; Gatta, V.; Piattelli, A.; Trubiani, O.; Mazzon, E. Biofunctionalized scaffold in bone tissue repair. Int. J. Mol. Sci. 2018, 19, 1022. [Google Scholar] [CrossRef]
- Pizzicannella, J.; Diomede, F.; Gugliandolo, A.; Chiricosta, L.; Bramanti, P.; Merciaro, I.; Orsini, T.; Mazzon, E.; Trubiani, O. 3D Printing PLA/Gingival Stem Cells/ EVs Upregulate miR-2861 and -210 during Osteoangiogenesis Commitment. Int. J. Mol. Sci. 2019, 20, 3256. [Google Scholar] [CrossRef]
- Loi, F.; Córdova, L.A.; Pajarinen, J.; Lin, T.; Yao, Z.; Goodman, S.B. Inflammation, fracture and bone repair. Bone 2016, 86, 119–130. [Google Scholar] [CrossRef]
- Sato, T.; Pajarinen, J.; Behn, A.; Jiang, X.; Lin, T.-H.; Loi, F.; Yao, Z.; Egashira, K.; Yang, F.; Goodman, S.B. The effect of local IL-4 delivery or CCL2 blockade on implant fixation and bone structural properties in a mouse model of wear particle induced osteolysis. J. Biomed. Mater. Res. A 2016, 104, 2255–2262. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Pajarinen, J.; Nabeshima, A.; Lu, L.; Nathan, K.; Yao, Z.; Goodman, S.B. Establishment of NF-κB sensing and interleukin-4 secreting mesenchymal stromal cells as an “on-demand” drug delivery system to modulate inflammation. Cytotherapy 2017, 19, 1025–1034. [Google Scholar] [CrossRef]
- Kohno, Y.; Lin, T.; Pajarinen, J.; Jämsen, E.; Romero-Lopez, M.; Maruyama, M.; Lo, C.-W.; Ueno, M.; Nathan, K.; Yao, Z.; et al. Treating Titanium Particle-Induced Inflammation with Genetically Modified NF-κB Sensing IL-4 Secreting or Preconditioned Mesenchymal Stem Cells in Vitro. ACS Biomater. Sci. Eng. 2019, 5, 3032–3038. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, M.; Moeinzadeh, S.; Guzman, R.A.; Zhang, N.; Storaci, H.W.; Utsunomiya, T.; Lui, E.; Huang, E.E.; Rhee, C.; Gao, Q.; et al. The efficacy of lapine preconditioned or genetically modified IL4 over-expressing bone marrow-derived mesenchymal stromal cells in corticosteroid-associated osteonecrosis of the femoral head in rabbits. Biomaterials 2021, 275, 120972. [Google Scholar] [CrossRef]
- Xie, H.; Cui, Z.; Wang, L.; Xia, Z.; Hu, Y.; Xian, L.; Li, C.; Xie, L.; Crane, J.; Wan, M.; et al. PDGF-BB secreted by preosteoclasts induces CD31hi Emcnhi vessel subtype in coupling osteogenesis. Nat. Med. 2014, 20, 1270–1278. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Baylink, D.J.; Brier-Jones, J.; Neises, A.; Kiroyan, J.B.; Rundle, C.H.; Lau, K.-H.W.; Zhang, X.-B. PDGFB-based stem cell gene therapy increases bone strength in the mouse. Proc. Natl. Acad. Sci. USA 2015, 112, E3893-900. [Google Scholar] [CrossRef]
- Montaseri, A.; Busch, F.; Mobasheri, A.; Buhrmann, C.; Aldinger, C.; Rad, J.S.; Shakibaei, M. IGF-1 and PDGF-bb suppress IL-1β-induced cartilage degradation through down-regulation of NF-κB signaling: Involvement of Src/PI-3K/AKT pathway. PLoS ONE 2011, 6, e28663. [Google Scholar] [CrossRef]
- Zhang, N.; Lo, C.W.; Utsunomiya, T.; Maruyama, M.; Huang, E.; Rhee, C.; Gao, Q.; Yao, Z.; Goodman, S.B. PDGF-BB and IL-4 co-overexpression is a potential strategy to enhance mesenchymal stem cell-based bone regeneration. Stem Cell Res. Ther. 2021, 12, 1–11. [Google Scholar] [CrossRef]
- Karpov, A.A.; Udalova, D.V.; Pliss, M.G.; Galagudza, M.M. Can the outcomes of mesenchymal stem cell-based therapy for myocardial infarction be improved? Providing weapons and armour to cells. Cell Prolif. 2017, 50. [Google Scholar] [CrossRef]
- Haider, H.K.; Ashraf, M. Strategies to promote donor cell survival: Combining preconditioning approach with stem cell transplantation. J. Mol. Cell. Cardiol. 2008, 45, 554–566. [Google Scholar] [CrossRef] [PubMed]
- Amiri, F.; Jahanian-Najafabadi, A.; Roudkenar, M.H. In vitro augmentation of mesenchymal stem cells viability in stressful microenvironments: In vitro augmentation of mesenchymal stem cells viability. Cell Stress Chaperones 2015, 20, 237–251. [Google Scholar] [CrossRef]
- English, K.; Wood, K.J. Mesenchymal stromal cells in transplantation rejection and tolerance. Cold Spring Harb. Perspect. Med. 2013, 3, a015560. [Google Scholar] [CrossRef]
- Qi, K.; Li, N.; Zhang, Z.; Melino, G. Tissue regeneration: The crosstalk between mesenchymal stem cells and immune response. Cell. Immunol. 2018, 326, 86–93. [Google Scholar] [CrossRef]
- Mountziaris, P.M.; Tzouanas, S.N.; Mikos, A.G. Dose effect of tumor necrosis factor-alpha on in vitro osteogenic differentiation of mesenchymal stem cells on biodegradable polymeric microfiber scaffolds. Biomaterials 2010, 31, 1666–1675. [Google Scholar] [CrossRef]
- Croes, M.; Oner, F.C.; Kruyt, M.C.; Blokhuis, T.J.; Bastian, O.; Dhert, W.J.A.; Alblas, J. Proinflammatory mediators enhance the osteogenesis of Human Mesenchymal stem cells after lineage commitment. PLoS ONE 2015, 10, e0132781. [Google Scholar] [CrossRef]
- Herzmann, N.; Salamon, A.; Fiedler, T.; Peters, K. Lipopolysaccharide induces proliferation and osteogenic differentiation of adipose-derived mesenchymal stromal cells in vitro via TLR4 activation. Exp. Cell Res. 2017, 350, 115–122. [Google Scholar] [CrossRef]
- Phillips, M.I.; Tang, Y.L. Genetic modification of stem cells for transplantation. Adv. Drug Deliv. Rev. 2008, 60, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Sage, E.K.; Thakrar, R.M.; Janes, S.M. Genetically modified mesenchymal stromal cells in cancer therapy. Cytotherapy 2016, 18, 1435–1445. [Google Scholar] [CrossRef]
- Foppiani, E.M.; Candini, O.; Mastrolia, I.; Murgia, A.; Grisendi, G.; Samarelli, A.V.; Boscaini, G.; Pacchioni, L.; Pinelli, M.; De Santis, G.; et al. Impact of HOXB7 overexpression on human adipose-derived mesenchymal progenitors. Stem Cell Res. Ther. 2019, 10, 1–12. [Google Scholar] [CrossRef]
- Lin, T.; Pajarinen, J.; Kohno, Y.; Maruyama, M.; Romero-Lopez, M.; Huang, J.-F.; Nathan, K.; Khan, T.N.; Yao, Z.; Goodman, S.B. Transplanted interleukin-4--secreting mesenchymal stromal cells show extended survival and increased bone mineral density in the murine femur. Cytotherapy 2018, 20, 1028–1036. [Google Scholar] [CrossRef] [PubMed]
- Bischoff, S.C.; Sellge, G.; Lorentz, A.; Sebald, W.; Raab, R.; Manns, M.P. IL-4 enhances proliferation and mediator release in mature human mast cells. Proc. Natl. Acad. Sci. USA 1999, 96, 8080–8085. [Google Scholar] [CrossRef]
- Bhattarai, P.; Thomas, A.K.; Cosacak, M.I.; Papadimitriou, C.; Mashkaryan, V.; Froc, C.; Reinhardt, S.; Kurth, T.; Dahl, A.; Zhang, Y.; et al. IL4/STAT6 Signaling Activates Neural Stem Cell Proliferation and Neurogenesis upon Amyloid-β42 Aggregation in Adult Zebrafish Brain. Cell Rep. 2016, 17, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Zhang, S.; Zeng, D.; Xia, L.; Lamichhane, A.; Jiang, X.; Zhang, F. RhPDGF-BB promotes proliferation and osteogenic differentiation of bone marrow stromal cells from streptozotocin-induced diabetic rats through ERK pathway. Biomed Res. Int. 2014, 2014, 637415. [Google Scholar] [CrossRef] [PubMed]
- Sotoca, A.M.; Roelofs-Hendriks, J.; Boeren, S.; van der Kraan, P.M.; Vervoort, J.; van Zoelen, E.J.J.; Piek, E. Comparative proteome approach demonstrates that platelet-derived growth factor C and D efficiently induce proliferation while maintaining multipotency of hMSCs. Exp. Cell Res. 2013, 319, 2649–2662. [Google Scholar] [CrossRef] [PubMed]
- Pajarinen, J.; Tamaki, Y.; Antonios, J.K.; Lin, T.-H.; Sato, T.; Yao, Z.; Takagi, M.; Konttinen, Y.T.; Goodman, S.B. Modulation of mouse macrophage polarization in vitro using IL-4 delivery by osmotic pumps. J. Biomed. Mater. Res. A 2015, 103, 1339–1345. [Google Scholar] [CrossRef][Green Version]
- Maruyama, M.; Rhee, C.; Utsunomiya, T.; Zhang, N.; Ueno, M.; Yao, Z.; Goodman, S.B. Modulation of the Inflammatory Response and Bone Healing. Front. Endocrinol. (Lausanne). 2020, 11, 386. [Google Scholar] [CrossRef]
- Pajarinen, J.; Lin, T.; Gibon, E.; Kohno, Y.; Maruyama, M.; Nathan, K.; Lu, L.; Yao, Z.; Goodman, S.B. Mesenchymal stem cell-macrophage crosstalk and bone healing. Biomaterials 2019, 196, 80–89. [Google Scholar] [CrossRef]
- Claes, L.; Recknagel, S.; Ignatius, A. Fracture healing under healthy and inflammatory conditions. Nat. Rev. Rheumatol. 2012, 8, 133–143. [Google Scholar] [CrossRef]
- Ueno, M.; Lo, C.-W.; Barati, D.; Conrad, B.; Lin, T.; Kohno, Y.; Utsunomiya, T.; Zhang, N.; Maruyama, M.; Rhee, C.; et al. Interleukin-4 overexpressing mesenchymal stem cells within gelatin-based microribbon hydrogels enhance bone healing in a murine long bone critical-size defect model. J. Biomed. Mater. Res. A 2020, 108, 2240–2250. [Google Scholar] [CrossRef] [PubMed]
- Bastidas-Coral, A.P.; Hogervorst, J.M.A.; Forouzanfar, T.; Kleverlaan, C.J.; Koolwijk, P.; Klein-Nulend, J.; Bakker, A.D. IL-6 counteracts the inhibitory effect of IL-4 on osteogenic differentiation of human adipose stem cells. J. Cell. Physiol. 2019, 234, 20520–20532. [Google Scholar] [CrossRef] [PubMed]
- Guzman, R.A.; Maruyama, M.; Moeinzadeh, S.; Lui, E.; Zhang, N.; Storaci, H.W.; Tam, K.; Huang, E.E.; Utsunomiya, T.; Rhee, C.; et al. The effect of genetically modified platelet-derived growth factor-BB over-expressing mesenchymal stromal cells during core decompression for steroid-associated osteonecrosis of the femoral head in rabbits. Stem Cell Res. Ther. 2021, 12, 503. [Google Scholar] [CrossRef] [PubMed]
- Ni, X.; Ou, C.; Guo, J.; Liu, B.; Zhang, J.; Wu, Z.; Li, H.; Chen, M. Lentiviral vector-mediated co-overexpression of VEGF and Bcl-2 improves mesenchymal stem cell survival and enhances paracrine effects in vitro. Int. J. Mol. Med. 2017, 40, 418–426. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day 3 | Day 5 | Day 7 | |
|---|---|---|---|
| GFP-MSCs | 1.8 ± 0.1 | 4.4 ± 0.1 | 5.8 ± 0.1 |
| GFP-pMSCs | 2.2 ± 0.2 | 4.0 ± 0.3 | 5.5 ± 0.5 |
| IL4-MSCs | 6.8 ± 0.5 | 20.2 ± 2.4 | 30.4 ± 4.4 |
| IL4-pMSCs | 7.3 ± 0.1 | 12.5 ± 0.6 | 19.9 ± 3.0 |
| PDGF-BB-MSCs | 3.3 ± 0.7 | 12.5 ± 0.6 | 27.4 ± 1.7 |
| PDGF-BB-pMSCs | 2.3 ± 0.1 | 6.7 ± 0.1 | 12.6 ± 0.2 |
| IL4-PDGF-BB-MSCs | 6.4 ± 1.3 | 20.5 ± 4.2 | 26.7 ± 3.7 |
| IL4-PDGF-BB-pMSCs | 5.5 ± 1.1 | 11.1 ± 1.7 | 16.3 ± 2.9 |
| GFP-MSCs | GFP-pMSCs | IL4-MSCs | IL4-pMSCs | PDGF-BB-MSCs | PDGF-BB-pMSCs | IL4-PDGF-BB-MSCs | IL4-PDGF-BB-pMSCs | |
| Cell proliferation | + | + | +++ | ++ | +++ | ++ | +++ | ++ |
| IL4/PDGF-BB expression | −/− | −/− | +++/− | ++/− | −/+++ | −/++ | +/+ | ++/++ |
| Osteogenic differentiation | ++ | ++ | − | − | +++ | ++ | + | ++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsubosaka, M.; Maruyama, M.; Huang, E.E.; Zhang, N.; Utsunomiya, T.; Gao, Q.; Shen, H.; Li, X.; Kushioka, J.; Hirata, H.; et al. Effect on Osteogenic Differentiation of Genetically Modified IL4 or PDGF-BB Over-Expressing and IL4-PDGF-BB Co-Over-Expressing Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro. Bioengineering 2021, 8, 165. https://doi.org/10.3390/bioengineering8110165
Tsubosaka M, Maruyama M, Huang EE, Zhang N, Utsunomiya T, Gao Q, Shen H, Li X, Kushioka J, Hirata H, et al. Effect on Osteogenic Differentiation of Genetically Modified IL4 or PDGF-BB Over-Expressing and IL4-PDGF-BB Co-Over-Expressing Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro. Bioengineering. 2021; 8(11):165. https://doi.org/10.3390/bioengineering8110165
Chicago/Turabian StyleTsubosaka, Masanori, Masahiro Maruyama, Elijah Ejun Huang, Ning Zhang, Takeshi Utsunomiya, Qi Gao, Huaishuang Shen, Xueping Li, Junichi Kushioka, Hirohito Hirata, and et al. 2021. "Effect on Osteogenic Differentiation of Genetically Modified IL4 or PDGF-BB Over-Expressing and IL4-PDGF-BB Co-Over-Expressing Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro" Bioengineering 8, no. 11: 165. https://doi.org/10.3390/bioengineering8110165
APA StyleTsubosaka, M., Maruyama, M., Huang, E. E., Zhang, N., Utsunomiya, T., Gao, Q., Shen, H., Li, X., Kushioka, J., Hirata, H., Yao, Z., Yang, Y. P., & Goodman, S. B. (2021). Effect on Osteogenic Differentiation of Genetically Modified IL4 or PDGF-BB Over-Expressing and IL4-PDGF-BB Co-Over-Expressing Bone Marrow-Derived Mesenchymal Stromal Cells In Vitro. Bioengineering, 8(11), 165. https://doi.org/10.3390/bioengineering8110165