Evaluation of Peripheral Blood and Cord Blood Platelet Lysates in Isolation and Expansion of Multipotent Mesenchymal Stromal Cells


 ,
,  , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Human Platelet Lysate
2.1.1. Peripheral Blood Platelet Lysate
2.1.2. Cord Blood Platelet Lysate
2.2. Protein Determination and Quantification Using Multiple Reaction Monitoring
2.3. Liquid Chromatography-Mass Spectrometry Setup
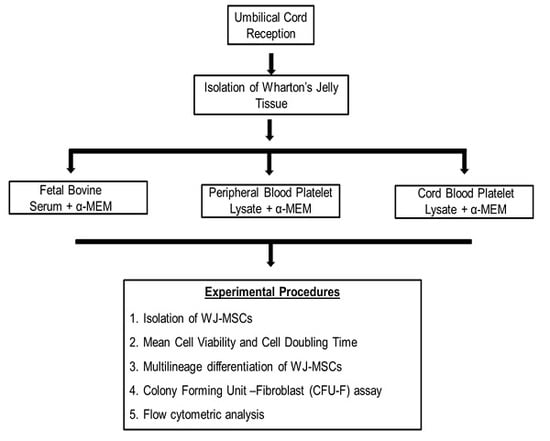
2.4. Collection of Human Umbilical Cords
2.5. Isolation and Culture of Wharton’s Jelly MSCs
2.6. Cell Viability and Growth Rate
2.7. Differentiation Capacity of MSCs
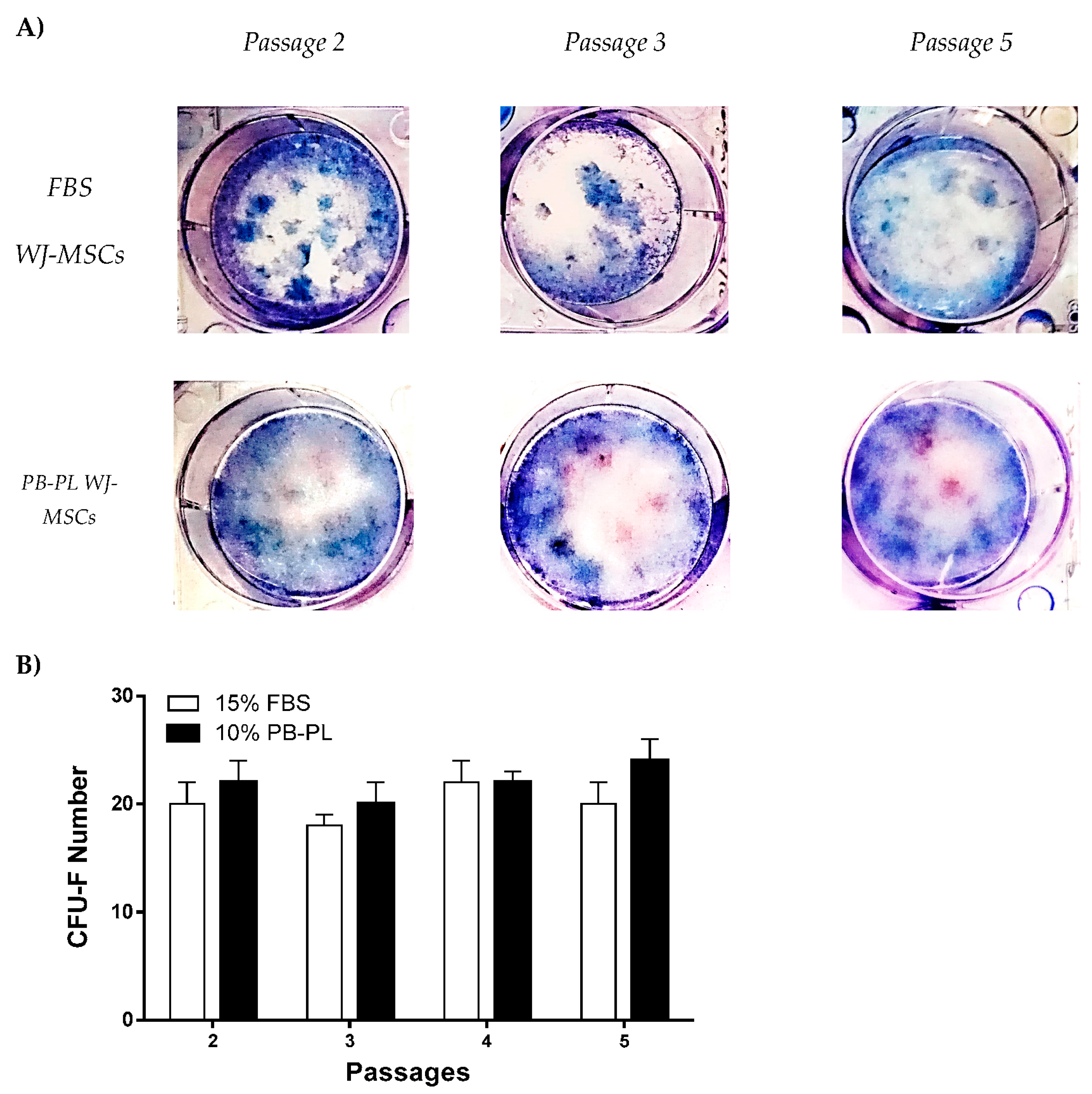
2.8. Colony-Forming Unit-Fibroblast (CFU-F) Assay
2.9. Phenotypic Characterization of WJ-MSCs
2.10. Growth Promotion Study and Media Validation
2.11. Statistical Analysis
3. Results
3.1. Preparation of Human Platelet Lysate
3.1.1. Peripheral Blood
3.1.2. Cord Blood
3.2. Protein Determination and Quantification in PB-PL and CB-PL
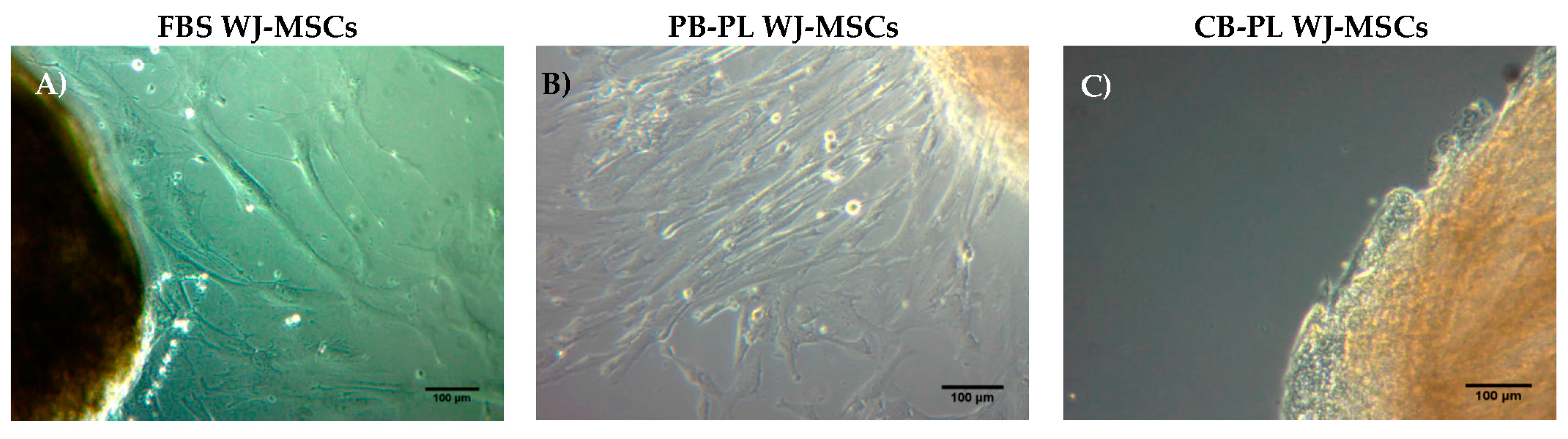
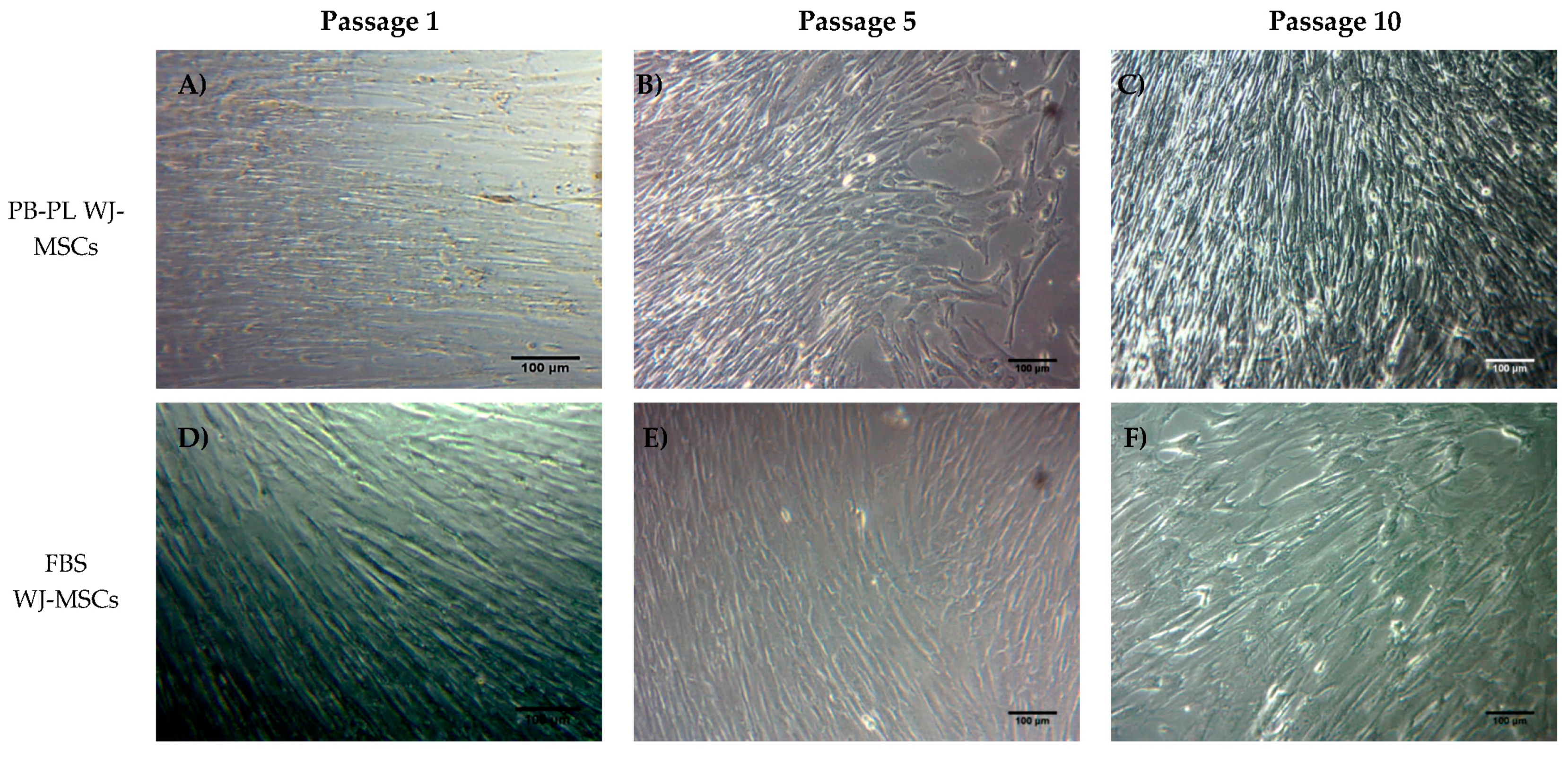
3.3. Isolation and Culture Characteristics of WJ-MSCs
3.4. Differentiation of WJ-MSCs
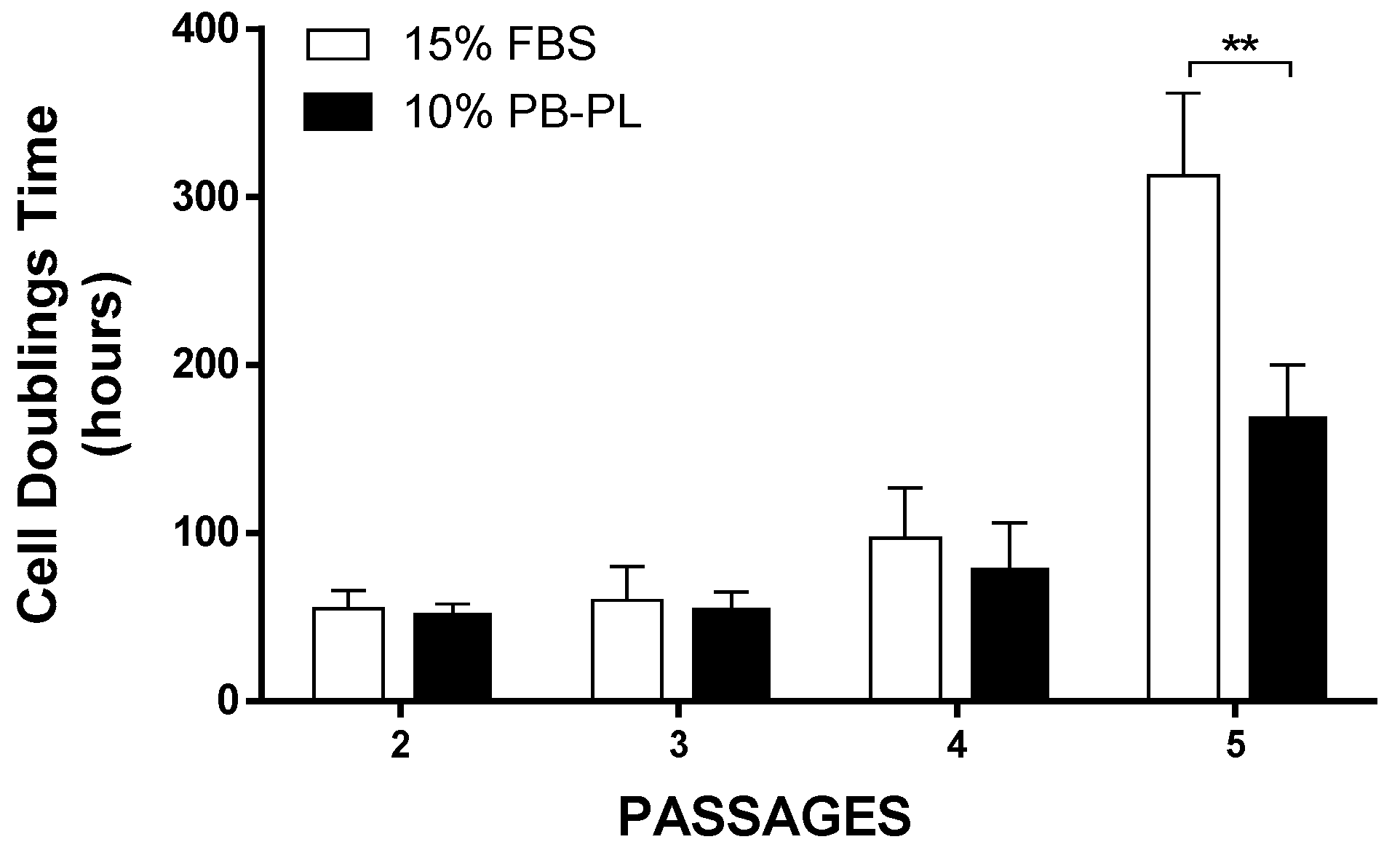
3.5. CFU–F of WJ-MSCs Cultured with FBS or PB-PL
3.6. Phenotypic Characterization
3.7. Growth Promotion Study and Media Validation Test Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Koshiro, S.; Kunihiro, Y.; Hiroaki, K.; Kaoru, Y.; Kei, S.; Xiangmei, Z.; Masahiro, K.; Yukichi, Z.; Ken, S.; Shingo, N.; et al. Spontaneous differentiation of human mesenchymal stem cells on poly-lactic-coglycolic acid nano-fiber scaffold. PLoS ONE 2016, 11, 1–15. [Google Scholar]
- Zhang, X.; Yamaoka, K.; Sonomoto, K.; Kaneko, H.; Satake, M.; Yamamoto, Y.; Kondo, M.; Zhao, J.; Miyagawa, I.; Yamagata, K.; et al. Local delivery of mesenchymal stem cells with poly-lactic-co-glycolic acid nano-fiber scaffold suppress arthritis in rats. PLoS ONE 2014, 9, 1–19. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Firestein, G.S. Evolving concepts of rheumatoid arthritis. Nature 2003, 423, 356–361. [Google Scholar] [CrossRef] [PubMed]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef] [PubMed]
- Le Blanc, K.; Mougiakakos, D. Multipotent mesenchymal stromal cells and the innate immune system. Nat. Rev. Immun. 2012, 12, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Yen, B.L.; Huang, H.I.; Chien, C.C.; Jui, H.Y.; Ko, B.S.; Yao, M.; Shun, C.T.; Yen, M.L.; Lee, M.C.; Chen, Y.C. Isolation of multipotent cells from human term placenta. Stem Cell 2005, 23, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Kern, S.; Eichler, H.; Stoeve, J.; Kluter, H.; Bieback, K. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cell 2006, 24, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Battula, V.L.; Bareiss, P.M.; Treml, S.; Conrad, S.; Albert, I.; Hojak, S.; Abele, H.; Schewe, B.; Just, L.; Skutella, T.; et al. Human placenta and bone marrow derived MSC cultured in serum-free, b-FGF-containing medium express cell surface frizzled-9 and SSEA-4 and give rise to multilineage differentiation. Differentiation 2007, 75, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.F.; Halford, K.W.; O’Hara, M.D.; Leeper, D.B.; Sokolov, B.P.; Pollard, M.D.; Bagasra, O.; Prockop, D.J. Cultured adherent cells from marrow can serve as long-lasting precursor cells for bone, cartilage and lung in irradiated mice. Proc. Natl. Acad. Sci. USA 1995, 92, 4857–4861. [Google Scholar] [CrossRef] [PubMed]
- Friedenstein, A.J.; Gorskaja, J.F.; Kulagina, N.N. Fibroblast precursors in normal and irradiated mouse hematopoietic organs. Exp. Hematol. 1976, 4, 267–274. [Google Scholar] [PubMed]
- Tuli, R.; Seghatoleslami, M.R.; Tuli, S.; Wang, M.L.; Hozack, W.J.; Manner, P.A.; Danielson, K.G.; Tuan, R.S. A simple, high-yield method for obtaining multi-potential mesenchymal progenitor cells from trabecular bone. Mol. Biotechnol. 2003, 23, 37–49. [Google Scholar] [CrossRef]
- Bakhshi, T.; Zabriskie, R.C.; Bodie, S.; Kidd, S.; Ramin, S.; Paganessi, L.A.; Gregory, S.A.; Fung, H.C.; Christopherson, K.W. Mesenchymal stem cells from the Wharton’s jelly of umbilical cord segments provide stromal support for the maintenance of cord blood hematopoietic stem cells during long-term ex vivo culture. Transfusion 2008, 48, 2638–2644. [Google Scholar] [CrossRef] [PubMed]
- Caplan, A.I. Mesenchymal stem cells. J. Orthop. Res. 1991, 9, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Prockop, D.J. Marrow stromal cells as stem cells for nonhematopoietic tissues. Science 1997, 276, 71–74. [Google Scholar] [CrossRef] [PubMed]
- Pittenger, M.F.; Mackay, A.M.; Beck, S.C.; Jaiswal, R.K.; Douglas, R.; Mosca, J.D.; Moorman, M.A.; Simonetti, D.W.; Craig, S.; Marshak, D.R. Multilineage potential of adult human mesenchymal stem cells. Science 1999, 284, 143–147. [Google Scholar] [CrossRef] [PubMed]
- Dennis, J.E.; Merriam, A.; Awadallah, A.; Yoo, J.U.; Johnstone, B.; Caplan, A.I. A quadripotential mesenchymal progenitor cell isolated from the marrow of an adult mouse. J. Bone Miner. Res. 1999, 14, 700–709. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef] [PubMed]
- Wagner, W.; Wein, F.; Seckinger, A.; Frankhauser, M.; Wirkner, U.; Krause, U.; Blake, J.; Schwager, C.; Eckstein, V.; Ansorge, W.; et al. Comparative characteristics of mesenchymal stem cells from human bone marrow, adipose tissue and umbilical cord blood. Exp. Hematol. 2005, 33, 1402–1416. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.L.; Anderson, C.; Medicetty, S.; Seshareddy, K.B.; Weiss, R.J.; Van der Werff, I.; Troyer, D.; McIntosh, K.R. Immune properties of human umbilical cord Wharton’s jelly-derived cells. Stem Cell 2008, 26, 2865. [Google Scholar] [CrossRef] [PubMed]
- Prasanna, S.J.; Gopalakrishnan, D.; Shankar, S.R.; Vasandan, A.B. Proinflammatory cytokines, IFNgamma and TNFalpha, influence immune properties of human bone marrow and Wharton jelly mesenchymal stem cells differentially. PLoS ONE 2010, 5, 9016. [Google Scholar] [CrossRef] [PubMed]
- Yoon, J.H.; Roh, E.Y.; Shin, S.; Jung, N.H.; Song, E.Y.; Chang, J.Y.; Kim, B.J.; Jeon, H.W. Comparison of explant derived and enzymatic digestion-derived MSCs and the growth factors from Wharton’s jelly. Biomed. Res. Int. 2013, 2013, 428726. [Google Scholar] [CrossRef] [PubMed]
- Costa-Pinto, A.R.; Salgado, A.J.; Correlo, V.M.; Sol, P.; Bhattacharya, M.; Charbord, P.; Reis, R.L.; Neves, N.M. Adhesion, proliferation and osteogenic differentiation of a mouse mesenchymal stem cell line (BMC9) seeded on novel melt-based chitosan/polyester 3D porous scaffolds. Tissue Eng. Part A 2008, 14, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Costa-Pinto, A.R.; Correlo, V.M.; Sol, P.C.; Bhattacharya, M.; Charbord, P.; Delorme, B.; Reis, R.L.; Neves, N.M. Osteogenic differentiation of human bone marrow mesenchymal stem cells seeded on melt based chitosan scaffolds for bone tissue engineering applications. Biomacromolecules 2009, 10, 2067–2073. [Google Scholar] [CrossRef] [PubMed]
- Hajivalili, M.; Pourgholi, F.; Kafil, H.S.; Jadidi-Niaragh, F.; Yousefi, M. Mesenchymal stem cells in the treatment of amyotrophic lateral sclerosis. Curr. Stem Cell Res. Ther. 2016, 11, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Connick, P.; Kolappan, M.; Patani, R.; Scott, M.A.; Crawley, C.; He, X.L.; Richardson, K.; Barber, K.; Webber, D.J.; Wheeler-Kingshott, C.A.; et al. The mesenchymal stem cells in multiple sclerosis (MSCIMS) trial protocol and baseline cohort characteristics: An open-label pre-test: post-test study with blinded outcome assessments. Trials 2011, 12, 62. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, F.E.; Carrión, F.; Villanueva, S.; Khoury, M. Mesenchymal stem cell treatment for autoimmune diseases: A critical review. Biol. Res. 2012, 45, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Dazzi, F.; Krampera, M. Mesenchymal stem cells and autoimmune diseases. Best. Pract. Res. Clin. Haematol. 2011, 24, 49–57. [Google Scholar] [CrossRef] [PubMed]
- De Miguel, M.P.; Fuentes-Julián, S.; Blázquez-Martínez, A.; Pascual, C.Y.; Aller, M.A.; Arias, J.; Arnalich-Montiel, F. Immunosuppressive properties of mesenchymal stem cells: Advances and applications. Curr. Mol. Med. 2012, 12, 574–591. [Google Scholar] [CrossRef] [PubMed]
- Sensebe, L.; Bourin, P.; Tarte, K. Good manufacturing practices production of mesenchymal stem/stromal cells. Hum. Gene Ther. 2011, 22, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, M.E.; Cometa, A.M.; Pagliara, D.; Vinti, L.; Rossi, F.; Cristantielli, R.; Palumbo, G.; Locatelli, F. Ex vivo expansion of mesenchymal stromal cells. Best. Pract. Res. Clin. Haematol. 2011, 24, 73. [Google Scholar] [CrossRef] [PubMed]
- Tuschong, L.; Soenen, S.L.; Blaese, R.M.; Candotti, F.; Muul, L.M. Immune response to foetal calf serum by two adenosine deaminase-deficient patients after T cell gene therapy. Hum. Gene Ther. 2002, 13, 1605–1610. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Panchalingam, K.M.; Rosenberg, L.; Behie, L.A. Ex vivo expansion of human mesenchymal stem cells in defined serum-free media. Stem Cells Int. 2012, 2012, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Schallmoser, K.; Bartmann, C.; Rohde, E.; Reinisch, A.; Kashofer, K.; Stadelmeyer, E.; Drexler, C.; Lanzer, G.; Linkesch, W.; Strunk, D. Human platelet lysate can replace foetal bovine serum for clinical-scale expansion of functional mesenchymal stromal cells. Transfusion 2007, 47, 1436–1446. [Google Scholar] [CrossRef] [PubMed]
- Bieback, K.; Hecker, A.; Kocaomer, A.; Lannert, H.; Schallmoser, K.; Strunk, D.; Ter, H. Human alternatives to foetal bovine serum for the expansion of mesenchymal stromal cells from bone marrow. Stem Cell 2009, 27, 2331–2341. [Google Scholar] [CrossRef] [PubMed]
- Muller, I.; Kordowich, S.; Holzwarth, C.; Spano, C.; Isensee, G.; Staiber, A.; Viebahn, S.; Gieseke, F.; Langer, H.; Gawaz, M.P.; et al. Animal serum-free culture conditions for isolation and expansion of multipotent mesenchymal stromal cells from human BM. Cytotherapy 2006, 8, 437–444. [Google Scholar] [CrossRef] [PubMed]
- Bernardo, M.E.; Avanzini, M.A.; Perotti, C.; Cometa, A.M.; Moretta, A.; Lenta, E.; Del Fante, C.; Novara, F.; de Silvestri, A.; Amendola, G.; et al. Optimization of in vitro expansion of human multipotent mesenchymal stromal cells for cell-therapy approaches: Further insights in the search for a foetal calf serum substitute. J. Cell. Physiol. 2007, 211, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Lange, C.; Cakiroglu, F.; Spiess, A.N.; Cappallo-Obermann, H.; Dierlamm, J.; Zander, A.R. Accelerated and safe expansion of human mesenchymal stromal cells in animal serum-free medium for transplantation and regenerative medicine. J. Cell. Physiol. 2007, 213, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Schallmoser, K.; Strunk, D. Preparation of pooled human platelet lysate (pHPL) as an efficient supplement for animal serum-free human stem cell cultures. J. Vis. Exp. 2009, 32, 1523. [Google Scholar] [CrossRef] [PubMed]
- Fekete, N.; Gadelorge, M.; Fürst, D.; Maurer, C.; Dausend, J.; Fleury-Cappellesso, S.; Mailänder, V.; Lotfi, R.; Ignatius, A.; Sensebé, L.; et al. Platelet lysate from whole blood-derived pooled platelet concentrates and apheresis-derived platelet concentrates for the isolation and expansion of human bone marrow mesenchymal stromal cells: Production process, content and identification of active components. Cytotherapy 2011, 14, 540–554. [Google Scholar]
- Murphy, M.B.; Blashki, D.; Buchanan, R.M.; Yazdi, I.K.; Ferrari, M.; Simmons, P.J.; Tasciotti, E. Adult and umbilical cord blood-derived platelet-rich plasma for mesenchymal stem cell proliferation, chemotaxis and cryo-preservation. Biomaterials 2012, 33, 5308–5316. [Google Scholar] [CrossRef] [PubMed]
- Longo, V.; Rebulla, P.; Pupella, S.; Zolla, L.; Rinalducci, S. Proteomic characterization of platelet gel releasate from adult peripheral and cord blood. Proteom. Clin. Appl. 2016, 10, 870–882. [Google Scholar] [CrossRef] [PubMed]
- Parazzi, V.; Lavazza, C.; Boldrin, V.; Montelatici, E.; Pallotti, F.; Marconi, M.; Lazzari, L. Extensive characterization of platelet gel releasate from cord blood in regenerative medicine. Cell Transplant. 2015, 24, 2573–2584. [Google Scholar] [CrossRef] [PubMed]
- Chatzistamatiou, T.K.; Papassavas, A.C.; Michalopoulos, E.; Gamaloutsos, C.; Mallis, P.; Gontika, I.; Panagouli, E.; Koussoulakos, S.L.; Stavropoulos-Giokas, C. Optimizing isolation culture and freezing methods to preserve Wharton’s jelly’s mesenchymal stem cell (MSC) properties: An MSC banking protocol validation for the Hellenic Cord Blood Bank. Transfusion 2014, 54, 3108–3120. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, T.C.; Ferrari, H.F.; Garcia, A.F.; Novais, J.B.; Silva-Frade, C.; Ferrarezi, M.C.; Andrade, A.L.; Gameiro, R. Isolation and characterization of Wharton’s jelly-derived multipotent mesenchymal stromal cells obtained from bovine umbilical cord and maintained in a defined serum-free three-dimensional system. BMC Biotechnol. 2012, 4, 12–18. [Google Scholar] [CrossRef] [PubMed]
- Fong, C.Y.; Subramanian, A.; Biswas, A.; Bongso, A. Freezing of fresh Wharton’s Jelly from human umbilical cords yields high post-thaw mesenchymal stem cell numbers for cell-based therapies. J. Cell. Biochem. 2016, 117, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Ben, A.N.; Jenhani, F.; Regaya, Z.; Berraeis, L.; Ben, O.T.; Ducrocq, E.; Domenech, J. Phenotypical and functional characteristics of mesenchymal stem cells from bone marrow: Comparison of culture using different media supplemented with human platelet lysate or foetal bovine serum. Stem Cell Res. Ther. 2012, 3, 25043. [Google Scholar]
- Bernardi, M.; Albiero, E.; Alghisi, A.; Chieregato, K.; Lievore, C.; Madeo, D.; Rodeghiero, F.; Astori, G. Production of human platelet lysate by use of ultrasound for ex vivo expansion of human bone marrow-derived mesenchymal stromal cells. Cytotherapy 2013, 15, 920–929. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, S.; Baraniak, P.R.; Copland, I.B.; Nerem, R.M.; McDevitt, T.C. Human platelet lysate stimulates high-passage and senescent human multipotent mesenchymal stromal cell growth and rejuvenation in vitro. Cytotherapy 2013, 15, 1469–1483. [Google Scholar] [CrossRef] [PubMed]
- Chevallier, N.; Anagnostou, F.; Zilber, S.; Bodivit, G.; Maurin, S.; Barrault, A.; Bierling, P.; Hernigou, P.; Layrolle, P.; Rouard, H. Osteoblastic differentiation of human mesenchymal stem cells with platelet lysate. Biomaterials 2010, 2010, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Ranzato, E.; Martinotti, S.; Volante, A.; Mazzucco, L.; Burlando, B. Platelet lysate modulates MMP-2 and MMP-9 expression, matrix deposition and cell-to-matrix adhesion in keratinocytes and fibroblasts. Exp. Dermatol. 2011, 20, 308–313. [Google Scholar] [CrossRef] [PubMed]
- Mojica-Henshaw, M.P.; Jacobson, P.; Morris, J.; Kelley, L.; Pierce, J.; Boyer, M.; Reems, J.A. Serum-converted platelet lysate can substitute for fetal bovine serum in human mesenchymal stromal cell cultures. Cytotherapy 2013, 15, 1458–1468. [Google Scholar] [CrossRef] [PubMed]
- Shih, D.T.; Burnouf, T. Preparation, quality criteria and properties of human blood platelet lysate supplements for ex vivo stem cell expansion. New Biotechnol. 2015, 32, 199–211. [Google Scholar] [CrossRef] [PubMed]
- Ebhardt, H.A.; Root, A.; Sander, C.; Aebersold, R. Applications of targeted proteomics in systems biology and translational medicine. Proteomics 2015, 15, 3193–3208. [Google Scholar] [CrossRef] [PubMed]
- Lawson, C.; Wolf, S. ICAM-1 signaling in endothelial cells. Pharmacol. Rep. 2009, 61, 22–32. [Google Scholar] [CrossRef]
- Le Blanc, K.; Tammik, C.; Rosendahl, K.; Zetterberg, E.; Ringdén, O. HLA expression and immunologic properties of differentiated and undifferentiated mesenchymal stem cells. Exp. Hematol. 2003, 31, 890–896. [Google Scholar] [CrossRef]
- Sheng, H.; Wang, Y.; Jin, Y.; Zhang, Q.; Zhang, Y.; Wang, L.; Shen, B.; Yin, S.; Liu, W.; Cui, L.; et al. A critical role of IFNgamma in priming MSC-mediated suppression of T cell proliferation through up-regulation of B7-H1. Cell Res. 2008, 18, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Ploederl, K.; Strasser, C.; Hennerbichler, S.; Peterbauer-Scherb, A.; Gabriel, C. Development and validation of a production process of platelet lysate for autologous use. Platelets 2010, 22, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Su, C.Y.; Kuo, Y.P.; Lin, Y.C.; Huang, C.T.; Tseng, Y.H.; Burnouf, T. A virally inactivated functional growth factor preparation from human platelet concentrates. Vox Sang. 2009, 97, 119–128. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Platelet Concentration (× 106/mL) | p-Value | |
|---|---|---|
| CB PRP Pools | PB PRP Pools | |
| 698 ± 23 | 655 ± 21 | 0.17 |
| Protein Identification | Accession Number | Ratio PB-PL/CB-PL |
|---|---|---|
| Interferon gamma receptor 1 precursor | INGR1_HUMAN | 6.8 ± 1.2 |
| Interleukin 1A | IL1A_HUMAN | 7.0 ± 2.1 |
| Interferon gamma precursor | IFNG_HUMAN | 5.6 ± 1.0 |
| Interleukin 1B | IL1B_HUMAN | 5.2 ± 0.8 |
| Tumour necrosis factor receptor type 1-associated DEATH domain protein | TRADD_HUMAN | 5.4 ± 1.9 |
| Intercellular adhesion molecule 1 precursor | ICAM1_HUMAN | 4.4 ± 1.5 |
| Tumour Necrosis Factor A | TNFA_HUMAN | 4.3 ± 1.7 |
| Interleukin 6 | IL6_HUMAN | 3.6 ± 0.6 |
| Vascular Endothelial Growth Factor A | VEGFA_HUMAN | 6.2 ± 4.1 |
| Fibroblast Growth Factor 2 | FGF2_HUMAN | 3.8 ± 0.8 |
| Platelet Derived Growth Factor A | PDGFA_HUMAN | 3.9 ± 1.6 |
| Interleukin 8 | IL8_HUMAN | 3.2 ± 0.6 |
| C-C motif chemokine 3 precursor | CCL3_HUMAN | 3.2 ± 0.7 |
| Transforming Growth Factor B1 precursor | TGFB1_HUMAN | 2.9 ± 0.3 |
| C-C motif chemokine 5 precursor | CCL5_HUMAN | 2.7 ± 0.3 |
| Vascular Cell Adhesion protein 1 precursor | VCAM1_HUMAN | 2.4 ± 0.3 |
| FBS Expanded WJ-MSCs | ||||
|---|---|---|---|---|
| n = 10 | n = 10 | n = 10 | n = 10 | |
| Passage | 2 | 3 | 4 | 5 |
| Mean cell Viability (%) | 83 ± 1 † | 88 ± 2 | 87 ± 3 | 85 ± 4 |
| Cell Doubling Time (hours) | 55 ± 11 | 60 ± 20 | 97 ± 30 | 313 ± 49 ‡ |
| PB-PL Expanded WJ-MSCs | ||||
| n = 10 | n = 10 | n = 10 | n = 10 | |
| Passage | 2 | 3 | 4 | 5 |
| Mean cell Viability (%) | 88 ± 2 † | 86 ± 1 | 88 ± 2 | 86 ± 3 |
| Cell Doubling Time (hours) | 50 ± 8 | 53 ± 12 | 77 ± 29 | 167 ± 33 ‡ |
| Cell Surface Markers | FBS Expanded WJ-MSCs | PB-Expanded WJ-MSCs | p Value |
|---|---|---|---|
| CD90 | 96.2 ± 0.6 | 96.7 ± 0.4 | 0.3707 |
| CD105 | 96.8 ± 0.1 | 98.6 ± 0.5 | 0.0053 |
| HLA-ABC | 94.0 ± 0.1 | 94.7 ± 0.5 | 0.1927 |
| CD73 | 96.4 ± 0.7 | 98.7 ± 0.6 | 0.0300 |
| CD29 | 95.6 ± 0.7 | 94.6 ± 0.5 | 0.1784 |
| CD44 | 96.0 ± 0.6 | 93.7 ± 0.3 | 0.0176 |
| CD19 | 1.2 ± 0.1 | 1.4 ± 0.1 | 0.1295 |
| CD3 | 1.8 ± 0.1 | 1.6 ± 0.1 | 0.0066 |
| CD31 | 1.6 ± 0.2 | 1.6 ± 0.1 | 0.8620 |
| CD14 | 1.3 ± 0.1 | 1.7 ± 0.2 | 0.1159 |
| HLA-DR | 1.1 ± 0.1 | 1.4 ± 0.2 | 0.1561 |
| CD45 | 1.4 ± 0.3 | 1.3 ± 0.1 | 0.7445 |
| CD34 | 1.5 ± 0.1 | 1.6 ± 0.2 | 0.4423 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christou, I.; Mallis, P.; Michalopoulos, E.; Chatzistamatiou, T.; Mermelekas, G.; Zoidakis, J.; Vlahou, A.; Stavropoulos-Giokas, C. Evaluation of Peripheral Blood and Cord Blood Platelet Lysates in Isolation and Expansion of Multipotent Mesenchymal Stromal Cells. Bioengineering 2018, 5, 19. https://doi.org/10.3390/bioengineering5010019
Christou I, Mallis P, Michalopoulos E, Chatzistamatiou T, Mermelekas G, Zoidakis J, Vlahou A, Stavropoulos-Giokas C. Evaluation of Peripheral Blood and Cord Blood Platelet Lysates in Isolation and Expansion of Multipotent Mesenchymal Stromal Cells. Bioengineering. 2018; 5(1):19. https://doi.org/10.3390/bioengineering5010019
Chicago/Turabian StyleChristou, Ioanna, Panagiotis Mallis, Efstathios Michalopoulos, Theofanis Chatzistamatiou, George Mermelekas, Jerome Zoidakis, Antonia Vlahou, and Catherine Stavropoulos-Giokas. 2018. "Evaluation of Peripheral Blood and Cord Blood Platelet Lysates in Isolation and Expansion of Multipotent Mesenchymal Stromal Cells" Bioengineering 5, no. 1: 19. https://doi.org/10.3390/bioengineering5010019
APA StyleChristou, I., Mallis, P., Michalopoulos, E., Chatzistamatiou, T., Mermelekas, G., Zoidakis, J., Vlahou, A., & Stavropoulos-Giokas, C. (2018). Evaluation of Peripheral Blood and Cord Blood Platelet Lysates in Isolation and Expansion of Multipotent Mesenchymal Stromal Cells. Bioengineering, 5(1), 19. https://doi.org/10.3390/bioengineering5010019