
Optimizing Yeast Surface-Displayed Unspecific Peroxygenase Production for Sustainable Biocatalysis




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Cultivation of YSD-UPO-Producing Komagataella phaffii in Shaking Flasks
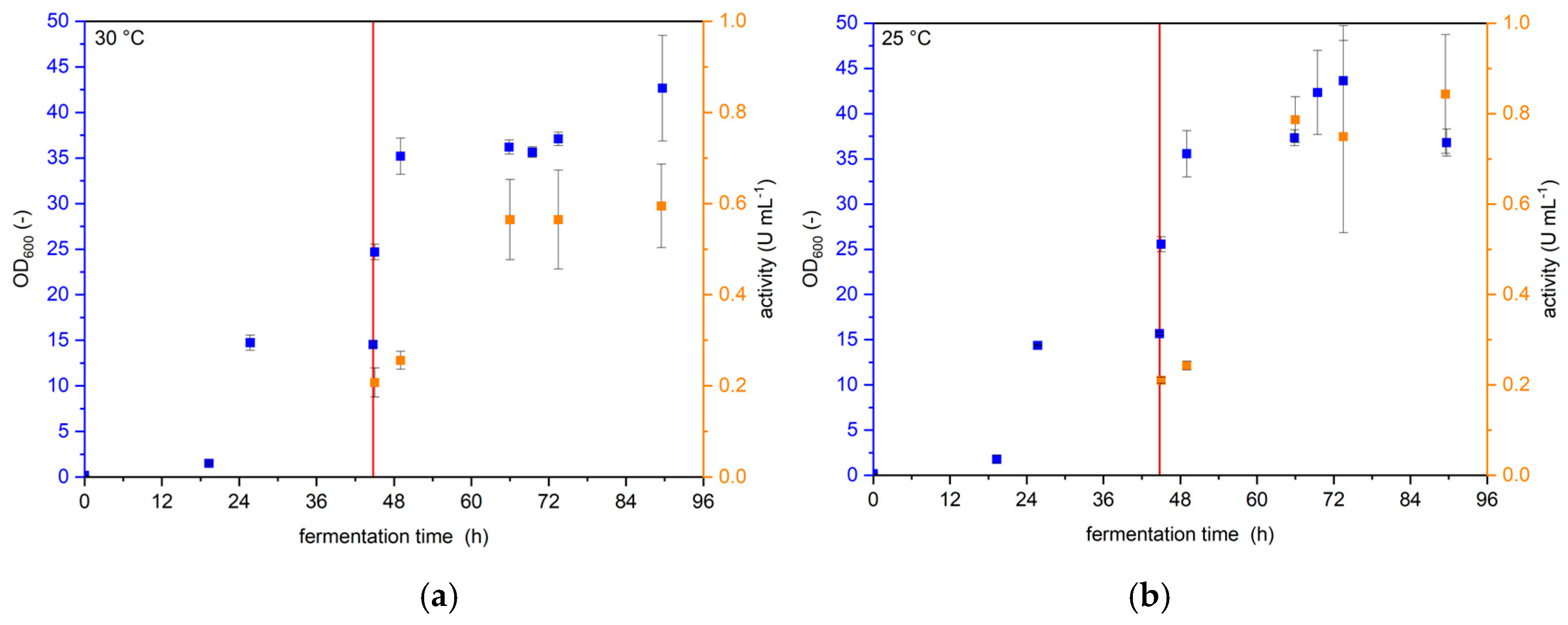
2.1.1. Temperature During Protein Expression
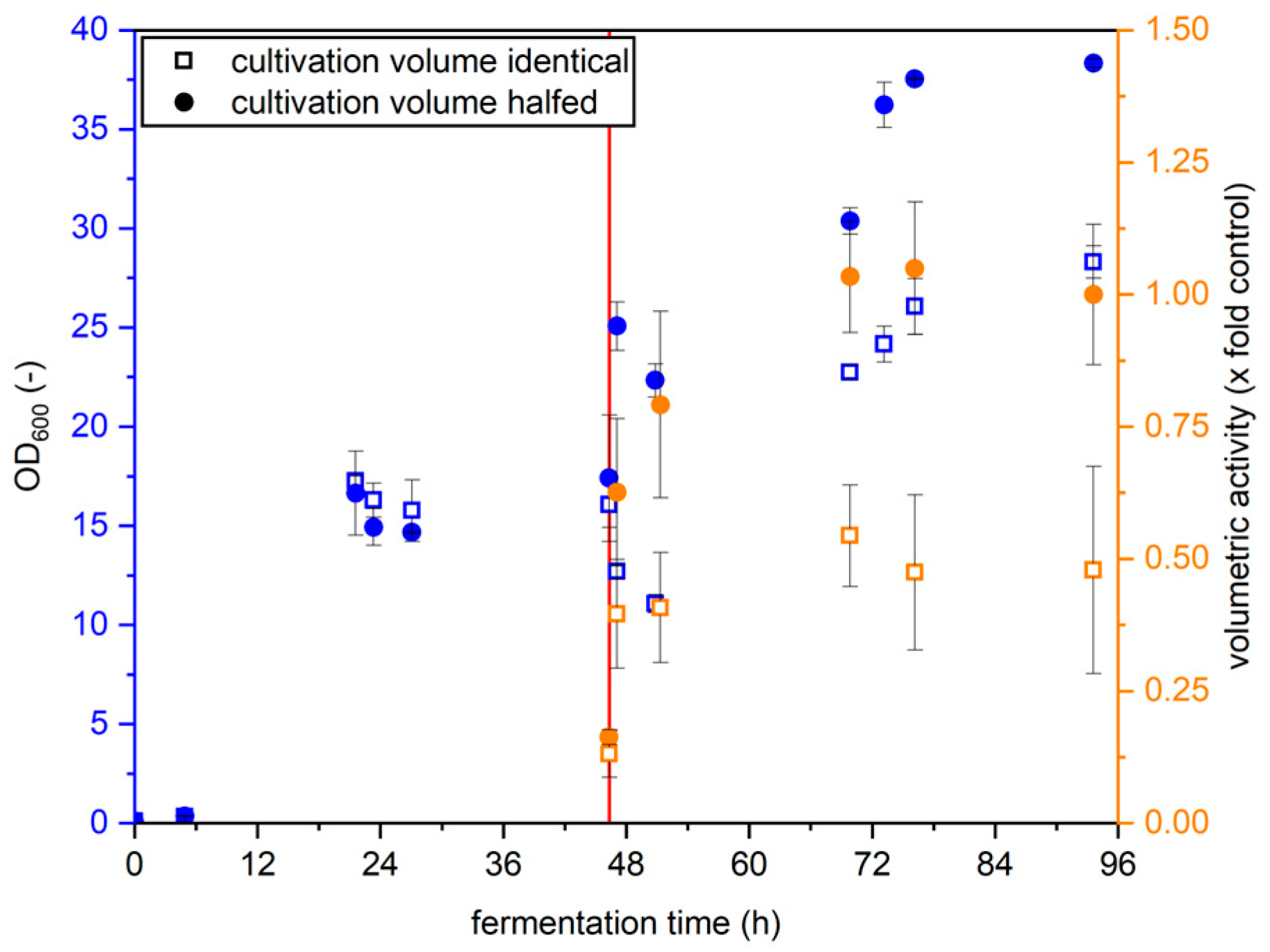
2.1.2. Cultivation Volume Reduction
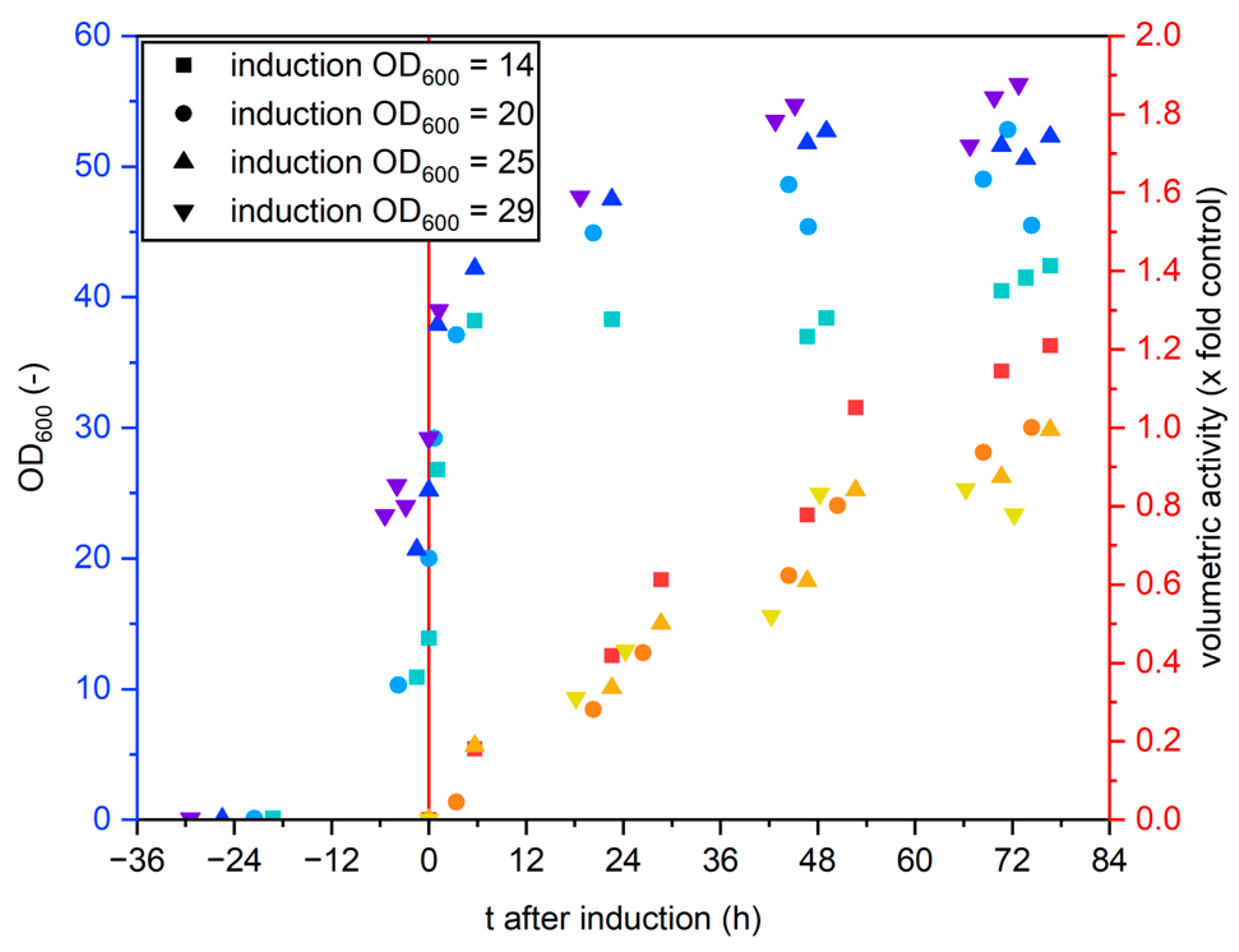
2.1.3. Induction at Different Optical Densities
2.2. Cultivation of YSD-UPO-Producing Komagataella phaffii in a Bioreactor
2.3. Optimization of Cultivation Parameters in BioLector XT® Cultivations and Design of Experiments
2.4. Enzyme Activity Assays
3. Results
3.1. Cultivation of YSD-UPO-Producing Komagataella phaffii in Shaking Flasks
3.1.1. Temperature During Protein Expression
3.1.2. Cultivation Volume Reduction
3.1.3. Induction at Different Optical Densities
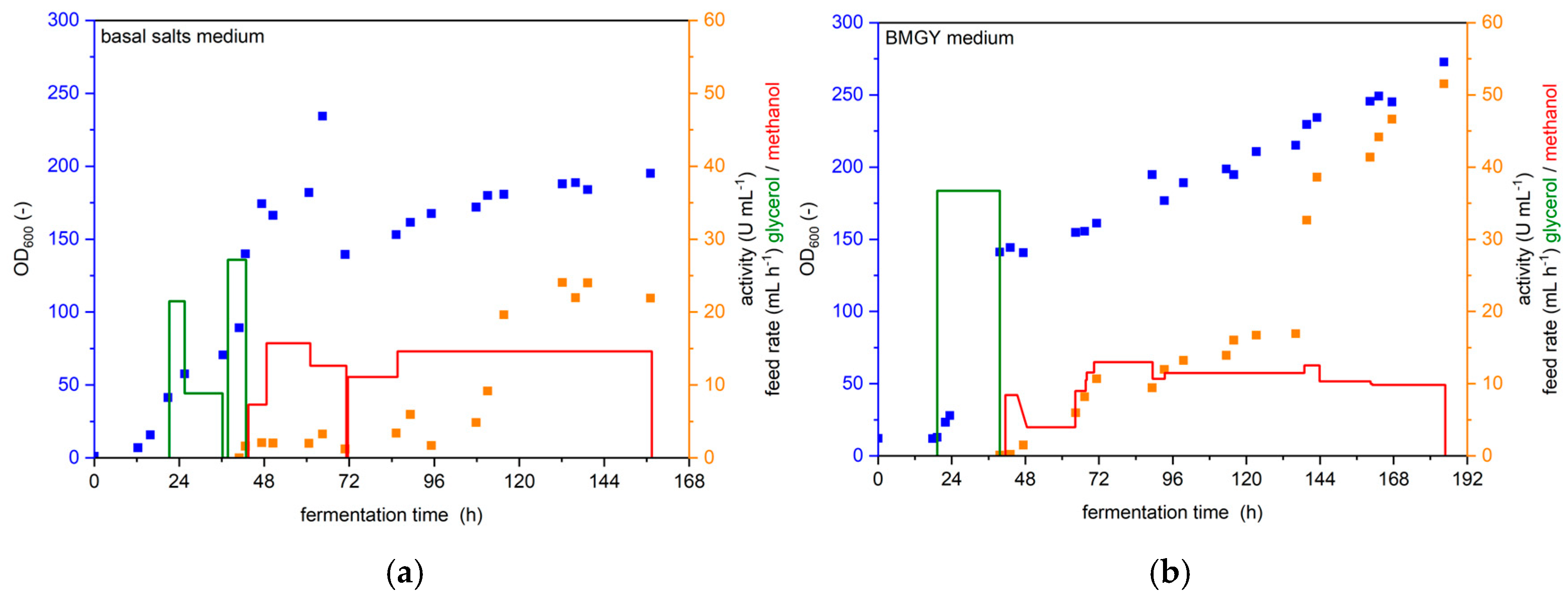
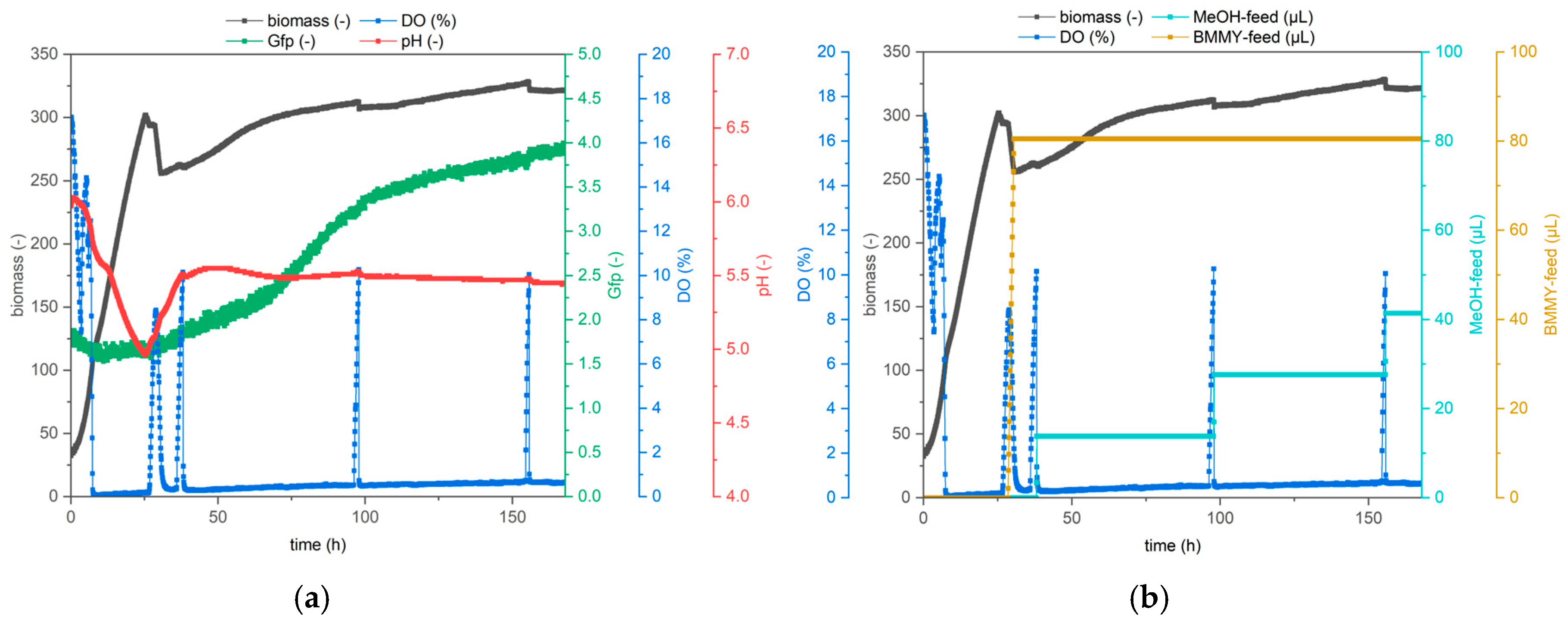
3.2. Comparison of Protocols for YSD-UPO Production in a Bioreactor
3.3. Optimization of YSD-UPO Production in a Parallelized Microfermentation System—BioLector XT®
3.3.1. UPO Activity/sfGfp Fluorescence Correlation
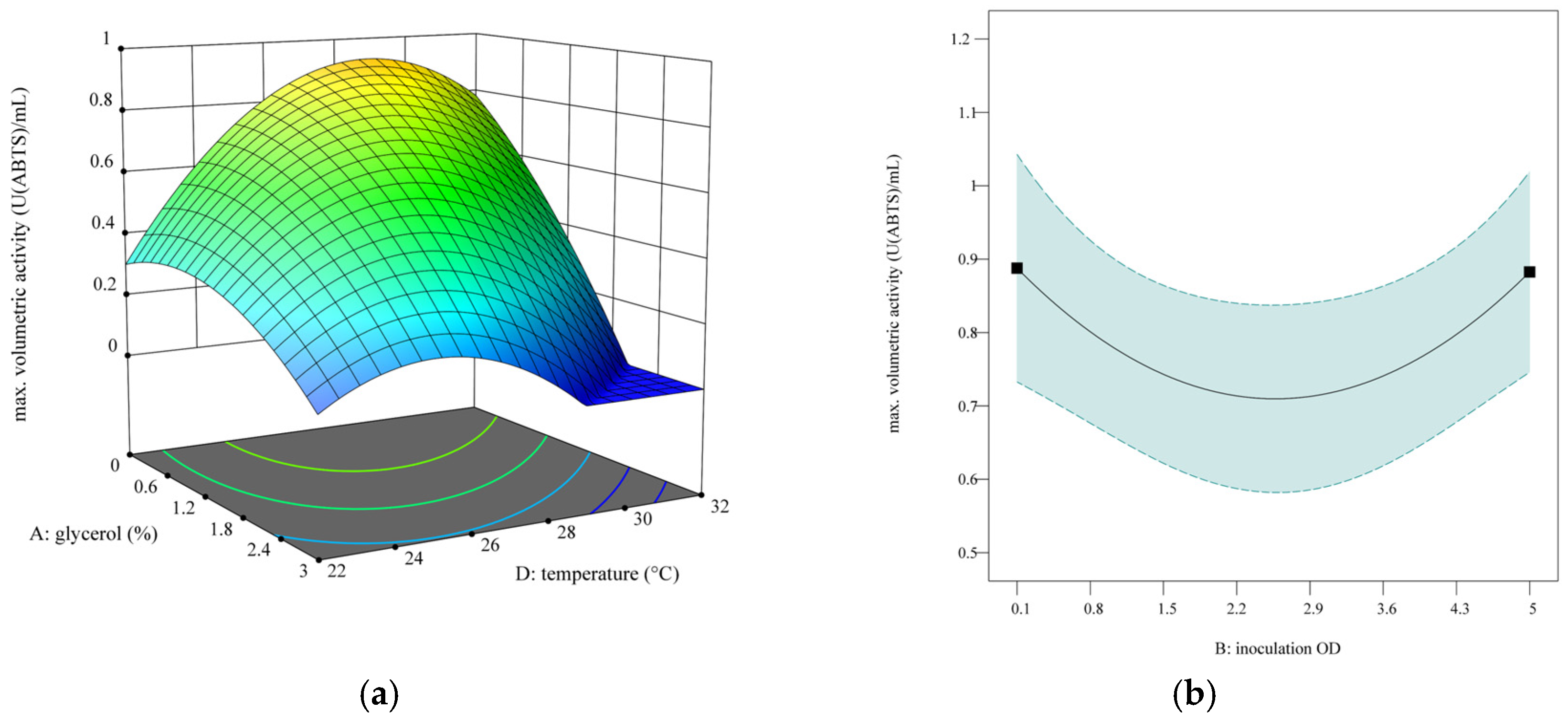
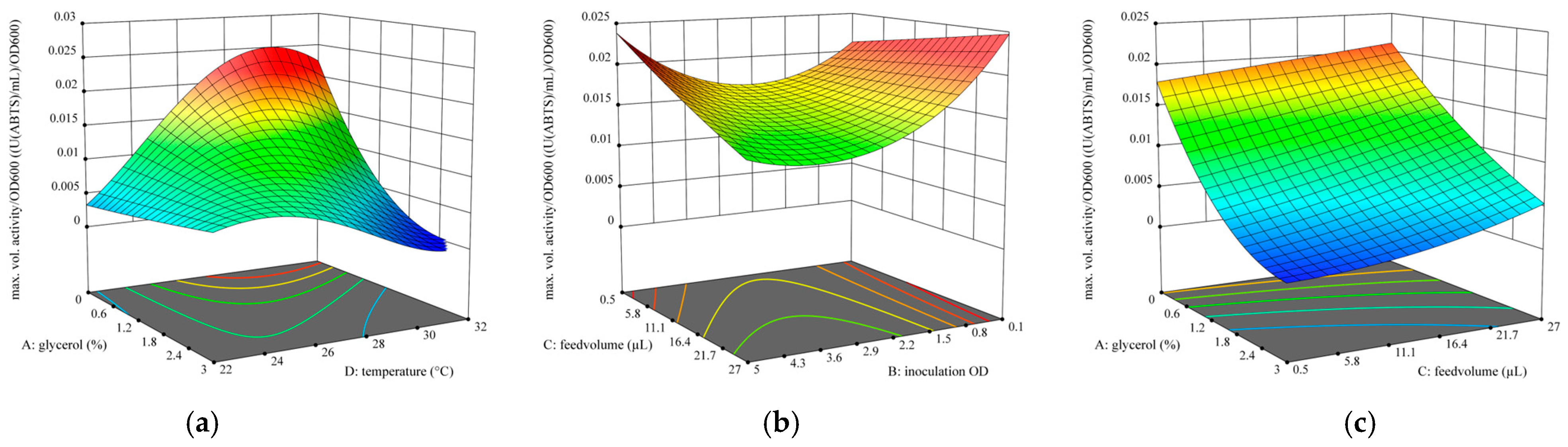
3.3.2. Design of Experiments
- Goal: maximize the maximally reached activity with consideration of reaching high activity per OD (six replicates).
- o
- % glycerol = 0
- o
- Inoculation OD = 0.1
- o
- MeOH feed volume = 0.65 µL
- o
- Temperature = 29.2 °C
- Goal: good maximally reached activity without consideration of activity per OD and using significantly different parameters than for one (four replicates).
- o
- % glycerol = 0
- o
- Inoculation OD = 5
- o
- MeOH feed volume = 17.2 µL
- o
- Temperature = 25.6 °C
- Inoculate BMMY medium from a preculture to an OD600 of 0.1 or from a colony to a volume of 800 µL.
- Choose your filter modules (DO required).
- Cycle time 00:00:00–00:10:00 is recommended.
- Humidity control: on.
- Temperature: constant 29 °C.
- Shaking frequency: constant 1200 rpm.
- Air: constant 30 mL/min.
- Set the maximum volume of cultivation wells to 1400 µL.
- Define feed wells for methanol feed.
- Enable signal-triggered methanol feed.
- Signal: DO > 15.00; pause: 15 min; pump volume: 0.65 µL (increase pause time for higher pump volumes to avoid accidental multiple feeding).
- 1. Start condition: DO < 12.0; block 20 min.
- 2. Start condition: DO > 15.0; block 20 min.
3.4. Sterilization and Storage of YSD-UPO
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Abbreviations
| ABTS | 2,2′-azino-bis(3ethylbenzothiazoline-6-sulfonic acid) |
| BMGY | Buffered medium with glycerol and yeast extract |
| BMMY | Buffered medium with methanol and yeast extract |
| DO | Dissolved oxygen |
| IEP | Immobilized enzyme process |
| KPI | Key performance indicators |
| MeOH | Methanol |
| OD600 | Optical density at 600 nm |
| Pi | Phosphate |
| PTM1 | Pichia Trace Metal solution 1 |
| RAE | Recovered activity efficiency |
| RT | Room temperature (≈20 °C) |
| SDG | Sustainable development goals |
| sfGfp | Superfolder Green fluorescent protein |
| STA | Space–time activity |
| STR | Stirred tank reactor |
| UN | United Nations |
| UPO | Unspecific peroxygenase |
| YPD | Yeast-Peptone-Dextrose (medium) |
| YSD | Yeast surface display |
References
- Beltrán-Nogal, A.; Sánchez-Moreno, I.; Méndez-Sánchez, D.; Santos, P.G.d.; Hollmann, F.; Alcalde, M. Surfing the wave of oxyfunctionalization chemistry by engineering fungal unspecific peroxygenases. Curr. Opin. Struct. Biol. 2022, 73, 102342. [Google Scholar] [CrossRef]
- Aranda, C.; Carro, J.; Gonzalez-Benjumea, A.; Babot, E.D.; Olmedo, A.; Linde, D.; Martinez, A.T.; Gutierrez, A. Advances in enzymatic oxyfunctionalization of aliphatic compounds. Biotechnol. Adv. 2021, 51, 107703. [Google Scholar] [CrossRef]
- Hobisch, M.; Holtmann, D.; Gomez de Santos, P.; Alcalde, M.; Hollmann, F.; Kara, S. Recent developments in the use of peroxygenases—Exploring their high potential in selective oxyfunctionalisations. Biotechnol. Adv. 2021, 51, 107615. [Google Scholar] [CrossRef]
- Bormann, S.; Gomez Baraibar, A.; Ni, Y.; Holtmann, D.; Hollmann, F. Specific oxyfunctionalisations catalysed by peroxygenases: Opportunities, challenges and solutions. Catal. Sci. Technol. 2015, 5, 2038–2052. [Google Scholar] [CrossRef]
- Hofrichter, M.; Ullrich, R. Oxidations catalyzed by fungal peroxygenases. Curr. Opin. Chem. Biol. 2014, 19, 116–125. [Google Scholar] [CrossRef]
- United Nations Sustainable Development Goals. Available online: https://sdgs.un.org/goals (accessed on 17 June 2025).
- Freakley, S.J.; Kochius, S.; van Marwijk, J.; Fenner, C.; Lewis, R.J.; Baldenius, K.; Marais, S.S.; Opperman, D.J.; Harrison, S.T.L.; Alcalde, M.; et al. A chemo-enzymatic oxidation cascade to activate C–H bonds with in situ generated H2O2. Nat. Commun. 2019, 10, 4178. [Google Scholar] [CrossRef]
- Babot, E.D.; Río, J.C.d.; Kalum, L.; Martínez, A.T.; Gutiérrez, A. Oxyfunctionalization of aliphatic compounds by a recombinant peroxygenase from Coprinopsis cinerea. Biotechnol. Bioeng. 2013, 110, 2323–2332. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.; Babot, E.D.; Ullrich, R.; Hofrichter, M.; Martinez, A.T.; del Rio, J.C. Regioselective oxygenation of fatty acids, fatty alcohols and other aliphatic compounds by a basidiomycete heme-thiolate peroxidase. Arch. Biochem. Biophys. 2011, 514, 33–43. [Google Scholar] [CrossRef]
- Teetz, N.; Lang, S.; Liese, A.; Holtmann, D. Yeast Surface Display Enables One-Step Production and Immobilization of Unspecific Peroxygenases. ChemCatChem 2024, 16, e202400908. [Google Scholar] [CrossRef]
- Santis, P.d.; Petrovai, N.; Meyer, L.-E.; Hobisch, M.; Kara, S. A holistic carrier-bound immobilization approach for unspecific peroxygenase. Front. Chem. 2022, 10, 985997. [Google Scholar] [CrossRef] [PubMed]
- Teetz, N.; Drommershausen, A.-L.; Gebele, L.; Holtmann, D. Holistic Evaluation of Enzyme Immobilization Processes: A Method for Evaluating the Entire Production Process. ChemCatChem 2025, 17, e00699. [Google Scholar] [CrossRef]
- Invitrogen “Pichia Fermentation Process Guidelines” Version B. Available online: https://tools.thermofisher.com/content/sfs/manuals/pichiaferm_prot.pdf (accessed on 17 June 2025).
- Peng, Y.; Wang, Y.; Liu, X.; Zhou, R.; Liao, X.; Min, Y.; Ma, L.; Wang, Y.; Rao, B. Expression and Surface Display of an Acidic Cold-Active Chitosanase in Pichia pastoris Using Multi-Copy Expression and High-Density Cultivation. Molecules 2022, 27, 800. [Google Scholar] [CrossRef] [PubMed]
- Rao, B.; Zhou, R.; Dong, Q.; Liao, X.; Liu, F.; Chen, W.; Liu, X.; Min, Y.; Wang, Y. Efficient Surface Display of L-glutamate Oxidase and L-amino Acid Oxidase on Pichia pastoris Using Multi-copy Expression Strains. Biotechnol. Bioprocess Eng. 2020, 25, 571–579. [Google Scholar] [CrossRef]
- Tonin, F.; Tieves, F.; Willot, S.; van Troost, A.; van Oosten, R.; Breestraat, S.; van Pelt, S.; Alcalde, M.; Hollmann, F. Pilot-Scale Production of Peroxygenase from Agrocybe aegerita. Org. Process Res. Dev. 2021, 25, 1414–1418. [Google Scholar] [CrossRef] [PubMed]
- Molina-Espeja, P.; Ma, S.; Mate, D.M.; Ludwig, R.; Alcalde, M. Tandem-yeast expression system for engineering and producing unspecific peroxygenase. Enzym. Microb. Technol. 2015, 73–74, 29–33. [Google Scholar] [CrossRef]
- Bormann, S.; Burek, B.O.; Ulber, R.; Holtmann, D. Immobilization of unspecific peroxygenase expressed in Pichia pastoris by metal affinity binding. Mol. Catal. 2020, 492, 110999. [Google Scholar] [CrossRef]
- Rotilio, L.; Swoboda, A.; Ebner, K.; Rinnofner, C.; Glieder, A.; Kroutil, W.; Mattevi, A. Structural and Biochemical Studies Enlighten the Unspecific Peroxygenase from Hypoxylon sp. EC38 as an Efficient Oxidative Biocatalyst. ACS Catal. 2021, 11, 11511–11525. [Google Scholar] [CrossRef]
- Zhao, L.-X.; Zou, S.-P.; Shen, Q.; Xue, Y.-P.; Zheng, Y.-G.; Zhao, L.-X.; Zou, S.-P.; Shen, Q.; Xue, Y.-P.; Zheng, Y.-G. Enhancing the expression of the unspecific peroxygenase in Komagataella phaffii through a combination strategy. Appl. Microbiol. Biotechnol. 2024, 108, 320. [Google Scholar] [CrossRef] [PubMed]
- Püllmann, P.; Weissenborn, M.J. Improving the Heterologous Production of Fungal Peroxygenases through an Episomal Pichia pastoris Promoter and Signal Peptide Shuffling System. ACS Synth. Biol. 2021, 10, 1360–1372. [Google Scholar] [CrossRef]
- Ebner, K.; Pfeifenberger, L.J.; Rinnofner, C.; Schusterbauer, V.; Glieder, A.; Winkler, M.; Ebner, K.; Pfeifenberger, L.J.; Rinnofner, C.; Schusterbauer, V.; et al. Discovery and Heterologous Expression of Unspecific Peroxygenases. Catalysts 2023, 13, 206. [Google Scholar] [CrossRef]
- Molina-Espeja, P.; Garcia-Ruiz, E.; Gonzalez-Perez, D.; Ullrich, R.; Hofrichter, M.; Alcalde, M. Directed evolution of unspecific peroxygenase from Agrocybe aegerita. Appl. Environ. Microbiol. 2014, 80, 3496–3507. [Google Scholar] [CrossRef]
- Martin-Diaz, J.; Molina-Espeja, P.; Hofrichter, M.; Hollmann, F.; Alcalde, M. Directed evolution of unspecific peroxygenase in organic solvents. Biotechnol. Bioeng. 2021, 118, 3002–3014. [Google Scholar] [CrossRef] [PubMed]
- Obst, U.; Lu, T.K.; Sieber, V. A Modular Toolkit for Generating Pichia pastoris Secretion Libraries. ACS Synth. Biol. 2017, 6, 1016–1025. [Google Scholar] [CrossRef]
- Portela, R.M.C.; Vogl, T.; Kniely, C.; Fischer, J.E.; Oliveira, R.; Glieder, A. Synthetic Core Promoters as Universal Parts for Fine-Tuning Expression in Different Yeast Species. ACS Synth. Biol. 2016, 6, 471–484. [Google Scholar] [CrossRef]
- Camboni, G.; Cartwright, J.; Grogan, G.; Camboni, G.; Cartwright, J.; Grogan, G. Identification of improved signal peptides for heterologous expression in Saccharomyces using a screen that exploits Gaussia luciferase. Sci. Rep. 2025, 15, 23962. [Google Scholar] [CrossRef]
- Yang, X.; Tang, H.; Song, M.; Shen, Y.; Hou, J.; Bao, X.; Yang, X.; Tang, H.; Song, M.; Shen, Y.; et al. Development of novel surface display platforms for anchoring heterologous proteins in Saccharomyces cerevisiae. Microb. Cell Factories 2019, 18, 85. [Google Scholar] [CrossRef] [PubMed]
- Lim, S.; Glasgow, J.E.; Interrante, M.F.; Storm, E.M.; Cochran, J.R. Dual display of proteins on the yeast cell surface simplifies quantification of binding interactions and enzymatic bioconjugation reactions. Biotechnol. J. 2017, 12, 1600696. [Google Scholar] [CrossRef]
- Hou, J.; Tyo, K.E.J.; Liu, Z.; Petranovic, D.; Nielsen, J. Metabolic engineering of recombinant protein secretion by Saccharomyces cerevisiae. FEMS Yeast Res. 2012, 12, 491–510. [Google Scholar] [CrossRef]
- Idiris, A.; Tohda, H.; Sasaki, M.; Okada, K.; Kumagai, H.; Giga-Hama, Y.; Takegawa, K.; Idiris, A.; Tohda, H.; Sasaki, M.; et al. Enhanced protein secretion from multiprotease-deficient fission yeast by modification of its vacuolar protein sorting pathway. Appl. Microbiol. Biotechnol. 2009, 85, 667–677. [Google Scholar] [CrossRef]
- Komeda, T.; Sakai, Y.; Kato, N.; Kondo, K. Construction of Protease-deficient Candida boidinii Strains Useful for Recombinant Protein Production: Cloning and Disruption of Proteinase A Gene (PEP4) and Proteinase B Gene. Biosci. Biotechnol. Biochem. 2002, 66, 628–631. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teetz, N.; Zuhse, L.; Holtmann, D. Optimizing Yeast Surface-Displayed Unspecific Peroxygenase Production for Sustainable Biocatalysis. Bioengineering 2025, 12, 822. https://doi.org/10.3390/bioengineering12080822
Teetz N, Zuhse L, Holtmann D. Optimizing Yeast Surface-Displayed Unspecific Peroxygenase Production for Sustainable Biocatalysis. Bioengineering. 2025; 12(8):822. https://doi.org/10.3390/bioengineering12080822
Chicago/Turabian StyleTeetz, Niklas, Luc Zuhse, and Dirk Holtmann. 2025. "Optimizing Yeast Surface-Displayed Unspecific Peroxygenase Production for Sustainable Biocatalysis" Bioengineering 12, no. 8: 822. https://doi.org/10.3390/bioengineering12080822
APA StyleTeetz, N., Zuhse, L., & Holtmann, D. (2025). Optimizing Yeast Surface-Displayed Unspecific Peroxygenase Production for Sustainable Biocatalysis. Bioengineering, 12(8), 822. https://doi.org/10.3390/bioengineering12080822