

Advancing Soft Tissue Reconstruction with a Ready-to-Use Human Adipose Allograft

Abstract

1. Introduction
2. Materials and Methods
2.1. Production of Human-Derived Adipose Allografts
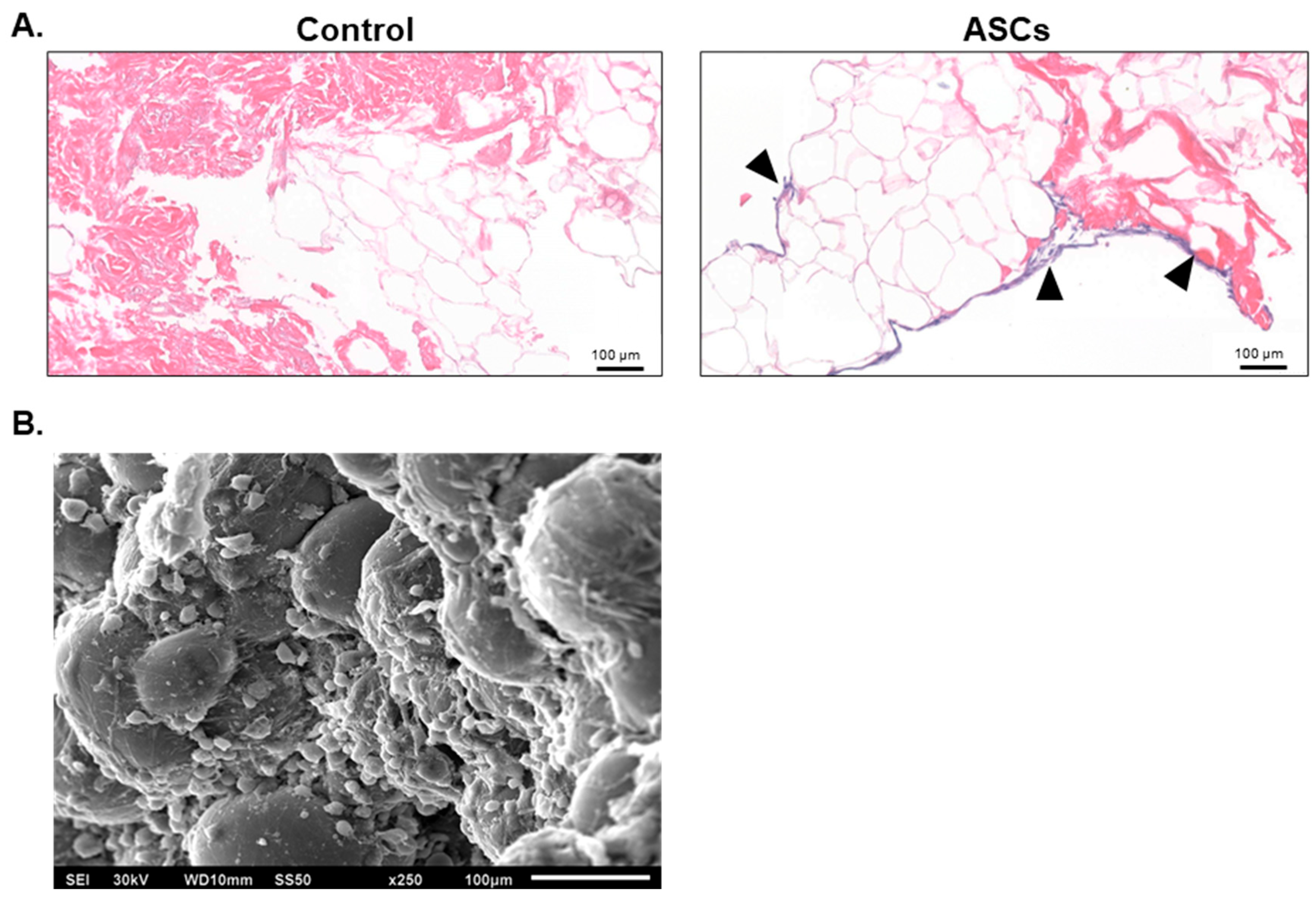
2.2. Histological Analysis
2.3. Scanning Electron Microscopy
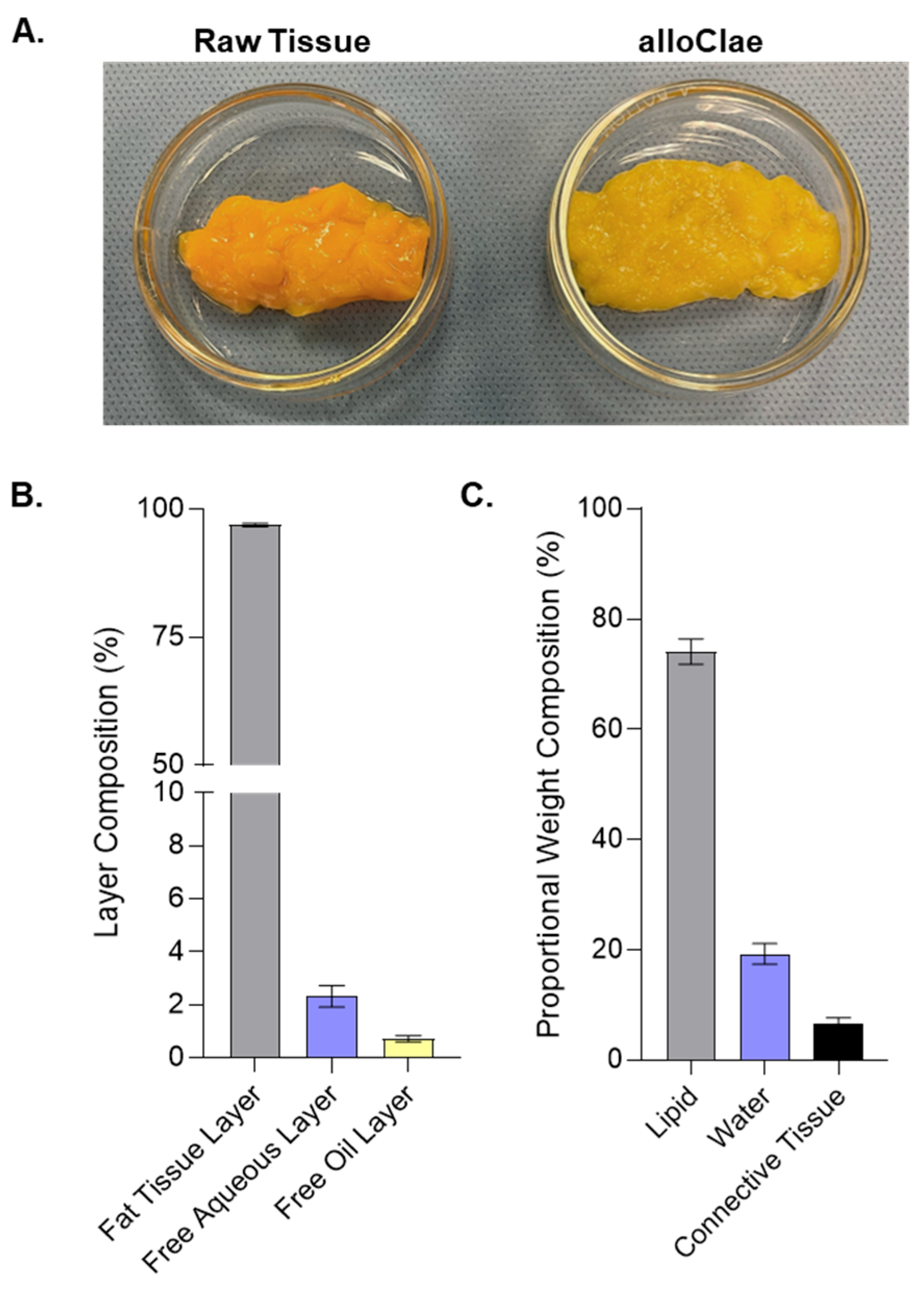
2.4. Weight Compositional Analysis
2.5. Layer Compositional Analysis
2.6. DNA Quantification
2.7. DAPI Staining of alloClae
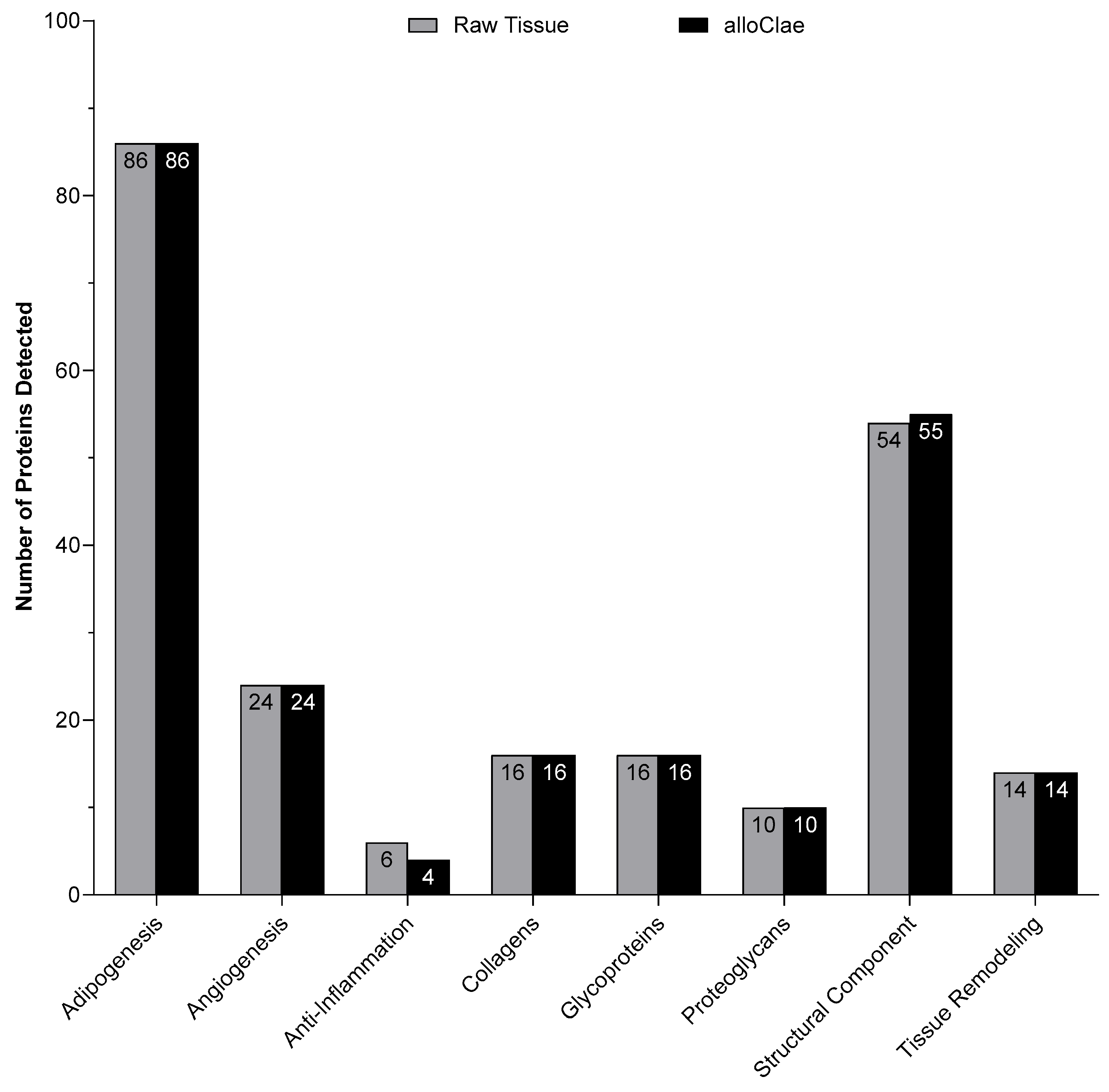
2.8. Proteomic Analysis
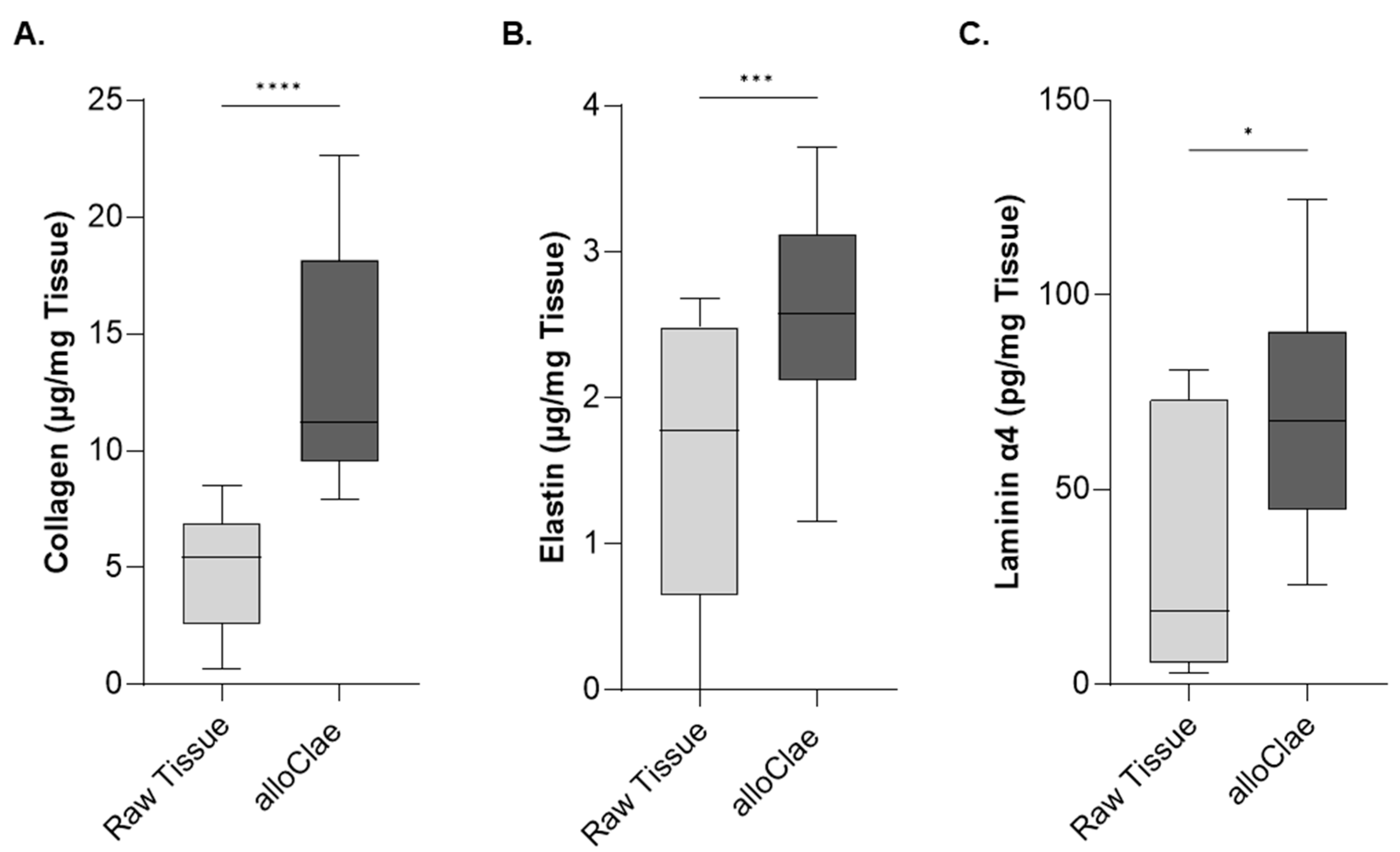
2.9. Collagen Analysis
2.10. Elastin Quantification
2.11. Laminin α4 ELISA
2.12. Biocompatibility Testing
2.13. Adipose-Derived Stem Cell (ASC) Adipogenesis Assay
2.14. Adipose-Derived Stem Cell Attachment Assay
2.15. Evaluation of alloClae in Athymic Mice
2.16. Statistical Analysis
3. Results and Discussion
3.1. Tissue Structure and Composition of alloClae
3.2. Proteomic and Quantitative Analysis of Extracellular Matrix Proteins in alloClae
3.3. In Vitro Biocompatibility of alloClae
3.4. Adipose-Derived Stem Cell Attachment and Differentiation with alloClae
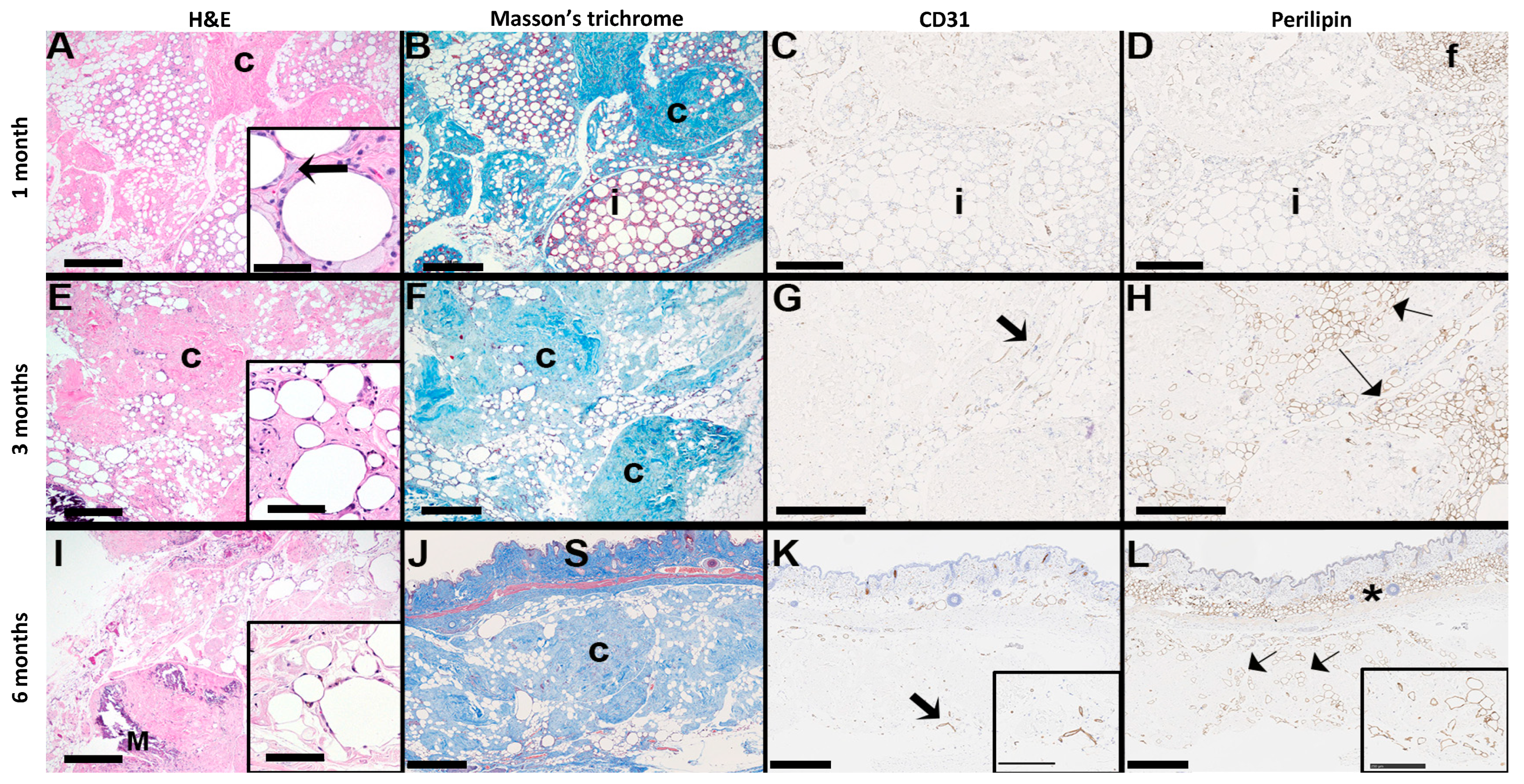
3.5. Long-Term Integration and Stability of alloClae in an Athymic Mouse Model
4. Conclusions
5. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AALAS | American Association for Laboratory Animal Science |
| AATB | American Association of Tissue Banks |
| ASC | Adipose-derived stem cell |
| CFR | Code of Federal Regulations |
| dAA | Decellularized adipose allograft |
| DNA | Deoxyribonucleic acid |
| ECM | Extracellular matrix |
| FDA | Food and Drug Administration |
| FOV | Field of view |
| GO | Gene Ontology |
| H&E | Hematoxylin and eosin |
| HMDS | Hexamethyldisilazane |
| IACUC | Institutional Animal Care and Use Committee |
| LDH | Lactate dehydrogenase |
| MST | Masson’s trichrome |
| SD | Standard deviation |
| SEM | Standard error of mean |
References
- Peirsman, A.; Nguyen, H.T.; Van Waeyenberge, M.; Ceballos, C.; Bolivar, J.; Kawakita, S.; Vanlauwe, F.; Tirpáková, Z.; Van Dorpe, S.; Van Damme, L.; et al. Vascularized adipose tissue engineering: Moving towards soft tissue reconstruction. Biofabrication 2023, 15, 032003. [Google Scholar] [CrossRef]
- Van Nieuwenhove, I.; Tytgat, L.; Ryx, M.; Blondeel, P.; Stillaert, F.; Thienpont, H.; Ottevaere, H.; Dubruel, P.; Van Vlierberghe, S. Soft tissue fillers for adipose tissue regeneration: From hydrogel development toward clinical applications. Acta Biomater. 2017, 63, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Fontes, T.; Brandão, I.; Negrão, R.; Martins, M.J.; Monteiro, R. Autologous fat grafting: Harvesting techniques. Ann. Med. Surg. 2018, 36, 212–218. [Google Scholar] [CrossRef]
- Kakagia, D.; Pallua, N. Autologous fat grafting: In search of the optimal technique. Surg. Innov. 2014, 21, 327–336. [Google Scholar] [CrossRef]
- Mughal, M.; Sindali, K.; Man, J.; Roblin, P. ‘Fat chance’: A review of adipose tissue engineering and its role in plastic and reconstructive surgery. Ann. R. Coll. Surg. Engl. 2021, 103, 245–249. [Google Scholar] [CrossRef] [PubMed]
- Wederfoort, J.L.M.; Hebels, S.A.; Heuts, E.M.; van der Hulst, R.R.W.J.; Piatkowski, A.A. Donor site complications and satisfaction in autologous fat grafting for breast reconstruction: A systematic review. J. Plast. Reconstr. Aesthetic Surg. 2022, 75, 1316–1327. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.S.; Choi, Y.C.; Kim, J.D.; Kim, E.J.; Lee, H.Y.; Kwon, I.C.; Cho, Y.W. Adipose tissue: A valuable resource of biomaterials for soft tissue engineering. Macromol. Res. 2014, 22, 932–947. [Google Scholar] [CrossRef]
- Kajimura, S. Advances in the understanding of adipose tissue biology. Nat. Rev. Endocrinol. 2017, 13, 69–70. [Google Scholar] [CrossRef]
- Mahoney, C.M.; Imbarlina, C.; Yates, C.C.; Marra, K.G. Current Therapeutic Strategies for Adipose Tissue Defects/Repair Using Engineered Biomaterials and Biomolecule Formulations. Front. Pharmacol. 2018, 9, 507. [Google Scholar] [CrossRef]
- Ruiz-Ojeda, F.J.; Méndez-Gutiérrez, A.; Aguilera, C.M.; Plaza-Díaz, J. Extracellular Matrix Remodeling of Adipose Tissue in Obesity and Metabolic Diseases. Int. J. Mol. Sci. 2019, 20, 4888. [Google Scholar] [CrossRef]
- Costa, A.; Naranjo, J.D.; Londono, R.; Badylak, S.F. Biologic Scaffolds. Cold Spring Harb. Perspect. Med. 2017, 7, a025676. [Google Scholar] [CrossRef] [PubMed]
- Mohiuddin, O.A.; Campbell, B.; Poche, J.N.; Thomas-Porch, C.; Hayes, D.A.; Bunnell, B.A.; Gimble, J.M. Decellularized Adipose Tissue: Biochemical Composition, in vivo Analysis and Potential Clinical Applications. Adv. Exp. Med. Biol. 2020, 1212, 57–70. [Google Scholar]
- Kasravi, M.; Ahmadi, A.; Babajani, A.; Mazloomnejad, R.; Hatamnejad, M.R.; Shariatzadeh, S.; Bahrami, S.; Niknejad, H. Immunogenicity of decellularized extracellular matrix scaffolds: A bottleneck in tissue engineering and regenerative medicine. Biomater. Res. 2023, 27, 10. [Google Scholar] [CrossRef] [PubMed]
- Shanti, R.M.; Janjanin, S.; Li, W.-J.; Nesti, L.J.; Mueller, M.B.; Tzeng, M.B.; Tuan, R.S. In vitro adipose tissue engineering using an electrospun nanofibrous scaffold. Ann. Plast. Surg. 2008, 61, 566–571. [Google Scholar] [CrossRef] [PubMed]
- Weiser, B.; Prantl, L.; Schubert, T.E.; Zellner, J.; Fischbach-Teschl, C.; Spruss, T.; Seitz, A.K.; Tessmar, J.; Goepferich, A.; Blunk, T. In vivo development and long-term survival of engineered adipose tissue depend on in vitro precultivation strategy. Tissue Eng. 2008, 14, 275–284. [Google Scholar] [CrossRef]
- Mauney, J.R.; Nguyen, T.; Gillen, K.; Kirker-Head, C.; Gimble, J.M.; Kaplan, D.L. Engineering adipose-like tissue in vitro and in vivo utilizing human bone marrow and adipose-derived mesenchymal stem cells with silk fibroin 3D scaffolds. Biomaterials 2007, 28, 5280–5290. [Google Scholar] [CrossRef]
- Choi, J.S.; Kim, B.S.; Kim, J.Y.; Kim, J.D.; Choi, Y.C.; Yang, H.; Park, K.; Lee, H.Y.; Cho, Y.W. Decellularized extracellular matrix derived from human adipose tissue as a potential scaffold for allograft tissue engineering. J. Biomed. Mater. Res. Part A 2011, 97, 292–299. [Google Scholar] [CrossRef]
- Mendibil, U.; Ruiz-Hernandez, R.; Retegi-Carrion, S.; Garcia-Urquia, N.; Olalde-Graells, B.; Abarrategi, A. Tissue-Specific Decellularization Methods: Rationale and Strategies to Achieve Regenerative Compounds. Int. J. Mol. Sci. 2020, 21, 5447. [Google Scholar] [CrossRef]
- Banyard, D.A.; Borad, V. Preparation, Characterization, and Clinical Implications of Human Decellularized Adipose Tissue Extra-cellular Matrix (hDAM): A Comprehensive Review. Aesthet. Surg. J. 2016, 36, 349–357. [Google Scholar] [CrossRef]
- Flynn, L. The use of decellularized adipose tissue to provide an inductive microenvironment for the adipogenic differentiation of human adipose-derived stem cells. Biomaterials 2010, 31, 4715–4724. [Google Scholar] [CrossRef]
- Wang, L.; Johnson, J.A.; Zhang, Q.; Beahm, E.K. Combining decellularized human adipose tissue extracellular matrix and adipose-derived stem cells for adipose tissue engineering. Acta Biomater. 2013, 9, 8921–8931. [Google Scholar] [CrossRef] [PubMed]
- Lin, M.; Ge, J.; Wang, X.; Dong, Z.; Xing, M.; Lu, F.; He, Y. Biochemical and biomechanical comparisions of decellularized scaffolds derived from porcine subcutaneous and visceral adipose tissue. J. Tissue Eng. 2019, 10, 2041731419888168. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Fu, S.; Luan, J. Selection of Mechanical Fragmentation Methods Based on Enzyme-Free Preparation of Decellularized Adipose-Derived Matrix. Bioengineering 2023, 10, 758. [Google Scholar] [CrossRef] [PubMed]
- Kokai, L.E.; Schilling, B.K.; Chnari, E.; Huang, Y.-C.; Imming, E.A.; Karunamurthy, A.; Khouri, R.K.; D’amico, R.A.; Coleman, S.R.; Marra, K.G.; et al. Injectable Allograft Adipose Matrix Supports Adipogenic Tissue Remodeling in the Nude Mouse and Human. Plast. Reconstr. Surg. 2019, 143, 299e–309e. [Google Scholar] [CrossRef]
- Giatsidis, G.; Succar, J.; Haddad, A.; Lago, G.; Schaffer, C.; Wang, X.; Schilling, B.; Chnari, E.; Matsumine, H.; Orgill, D.P.; et al. Preclinical Optimization of a Shelf-Ready, Injectable, Human-Derived, Decellularized Allograft Adipose Matrix. Tissue Eng. Part A 2019, 25, 271–287. [Google Scholar] [CrossRef]
- Zhang, J.; Xiang, Y.; Yang, Q.; Chen, J.; Liu, L.; Jin, J.; Zhu, S. Adipose-derived stem cells derived decellularized extracellular matrix enabled skin regeneration and remodeling. Front. Bioeng. Biotechnol. 2024, 12, 1347995. [Google Scholar] [CrossRef]
- Kokai, L.E.; Sivak, W.N.; Schilling, B.K.; Karunamurthy, A.; Egro, F.M.M.; Schusterman, M.A.; Minteer, D.M.; Simon, P.R.; D’amico, R.A.; Rubin, J.P. Clinical Evaluation of an Off-the-Shelf Allogeneic Adipose Matrix for Soft Tissue Reconstruction. Plast. Reconstr. Surg.-Glob. Open 2020, 8, e2574. [Google Scholar] [CrossRef]
- Salgado, C.J.; Gonzalez, D.I.; Gelles-Soto, D.; Mercado, A. Use of allograft fat for aesthetic and functional restoration of soft tissue contour deformities. J. Surg. Case Rep. 2023, 2023, rjac629. [Google Scholar] [CrossRef]
- Shahin, T.B.; Vaishnav, K.V.; Watchman, M.; Subbian, V.; Larson, E.; Chnari, E.; Armstrong, D.G. Tissue Augmentation with Allograft Adipose Matrix For the Diabetic Foot in Remission. Plast. Reconstr. Surg. -Glob. Open 2017, 5, e1555. [Google Scholar] [CrossRef]
- Anderson, A.E.; Wu, I.; Parrillo, A.J.; Wolf, M.T.; Maestas, D.R.; Graham, I.; Tam, A.J.; Payne, R.M.; Aston, J.; Cooney, C.M.; et al. An Immunologically Active, Adipose-Derived Extracellular Matrix Biomaterial For Soft Tissue Reconstruction: Concept To Clinical Trial. npj Regen. Med. 2022, 7, 6. [Google Scholar] [CrossRef]
- Wu, I.B.; Nahas, Z.; Kimmerling, K.A.B.; Rosson, G.D.; Elisseeff, J.H. An Injectable Adipose Matrix for Soft-Tissue Reconstruction. Plast. Reconstr. Surg. 2012, 129, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Panayi, A.C.; Mi, B.; Fu, S.; Orgill, D.P. Comparative Analysis of Two Automated Fat-processing Systems. Plast. Reconstr. Surg. Glob. Open 2020, 8, e2587. [Google Scholar] [CrossRef] [PubMed]
- Bourdon, A.; François, V.; Zhang, L.; Lafoux, A.; Fraysse, B.; Toumaniantz, G.; Larcher, T.; Girard, T.; Ledevin, M.; Lebreton, C.; et al. Evaluation of the dystrophin carboxy-terminal domain for micro-dystrophin gene therapy in cardiac and skeletal muscles in the DMDmdx rat model. Gene Ther. 2022, 29, 520–535. [Google Scholar] [CrossRef] [PubMed]
- Chiu, W.-K.; Fracol, M.; Feld, L.N.; Qiu, C.S.; Kim, J.Y. A Comparison of Fat Graft Processing Techniques: Outcomes in 1,158 Procedures in Prosthetic Breast Reconstructions. Plast. Reconstr. Surg. Glob. Open 2019, 7, e2276. [Google Scholar] [CrossRef]
- Nemir, S.; Hanson, S.E.; Chu, C.K. Surgical Decision Making in Autologous Fat Grafting: An Evidence-Based Review of Techniques to Maximize Fat Survival. Aesthetic Surg. J. 2021, 41 (Suppl. 1), S3–S15. [Google Scholar] [CrossRef]
- Fang, C.; Patel, P.B.; Li, H.; Huang, L.T.; Wan, H.B.; Collins, S.B.; Connell, T.L.B.; Xu, H. Physical, Biochemical, and Biologic Properties of Fat Graft Processed via Different Methods. Plast. Reconstr. Surg. Glob. Open 2020, 8, e3010. [Google Scholar] [CrossRef]
- Hivernaud, V.; Lefourn, B.; Robard, M.; Guicheux, J.; Weiss, P. Autologous fat grafting: A comparative study of four current commercial protocols. J. Plast. Reconstr. Aesthetic Surg. 2017, 70, 248–256. [Google Scholar] [CrossRef]
- McKee, T.J.; Perlman, G.; Morris, M.; Komarova, S.V. Extracellular matrix composition of connective tissues: A systematic review and meta-analysis. Sci. Rep. 2019, 9, 10542. [Google Scholar] [CrossRef]
- Hussein, K.H.; Saleh, T.; Ahmed, E.; Kwak, H.H.; Park, K.M.; Yang, S.R.; Woo, H.M. Biocompatibility and hemocompatibility of efficiently decellularized whole porcine kidney for tissue engineering. J. Biomed. Mater. Res. Part A 2018, 106, 2034–2047. [Google Scholar] [CrossRef]
- Stroh, A.M.; Lynch, C.E.; Lester, B.E.; Minchev, K.; Chambers, T.L.; Montenegro, C.F.; Martinez, C.C.; Fountain, W.A.; Trappe, T.A.; Trappe, S.W. Human adipose and skeletal muscle tissue DNA, RNA, and protein content. J. Appl. Physiol. 2021, 131, 1370–1379. [Google Scholar] [CrossRef]
- Porzionato, A.; Sfriso, M.M.; Macchi, V.; Rambaldo, A.; Lago, G.; Lancerotto, L.; Vindigni, V.; De Caro, R. Decellularized omentum as novel biologic scaffold for reconstructive surgery and regenerative medicine. Eur. J. Histochem. 2014, 57, e4. [Google Scholar] [CrossRef] [PubMed]
- Gimble, J.M.; Katz, A.J.; Bunnell, B.A. Adipose-Derived Stem Cells for Regenerative Medicine. Circ. Res. 2007, 100, 1249–1260. [Google Scholar] [CrossRef]
- Brown, B.N.; Badylak, S.F. Extracellular matrix as an inductive scaffold for functional tissue reconstruction. Transl. Res. 2014, 163, 268–285. [Google Scholar] [CrossRef] [PubMed]
- Jadalannagari, S.; Converse, G.; McFall, C.; Buse, E.; Filla, M.; Villar, M.T.; Artigues, A.; Mellot, A.J.; Wang, J.; Detamore, M.S.; et al. Decellularized Wharton’s Jelly from human umbilical cord as a novel 3D scaffolding material for tissue engineering applications. PLoS ONE 2017, 12, e0172098. [Google Scholar] [CrossRef]
- Naba, A. Mechanisms of assembly and remodelling of the extracellular matrix. Nat. Rev. Mol. Cell Biol. 2024, 25, 865–885. [Google Scholar] [CrossRef]
- Trębacz, H.; Barzycka, A. Mechanical Properties and Functions of Elastin: An Overview. Biomolecules 2023, 13, 574. [Google Scholar] [CrossRef] [PubMed]
- Dörschmann, P.; Böser, S.; Isik, D.; Arndt, C.; Roider, J.; Selhuber-Unkel, C.; Klettner, A. Influence of carrier materials and coatings on retinal pigment epithelium cultivation and functions. Exp. Eye Res. 2022, 219, 109063. [Google Scholar] [CrossRef]
- Patten, J.; Wang, K. Fibronectin in development and wound healing. Adv. Drug Deliv. Rev. 2021, 170, 353–368. [Google Scholar] [CrossRef]
- Stamati, K.; Priestley, J.V.; Mudera, V.; Cheema, U. Laminin promotes vascular network formation in 3D in vitro collagen scaffolds by regulating VEGF uptake. Exp. Cell Res. 2014, 327, 68–77. [Google Scholar] [CrossRef]
- Neishabouri, A.; Khaboushan, A.S.; Daghigh, F.; Kajbafzadeh, A.-M.; Zolbin, M.M. Decellularization in Tissue Engineering and Regenerative Medicine: Evaluation, Modification, and Application Methods. Front. Bioeng. Biotechnol. 2022, 10, 805299. [Google Scholar] [CrossRef]
- Feoktistova, M.; Geserick, P.; Leverkus, M. Crystal Violet Assay for Determining Viability of Cultured Cells. Cold Spring Harb. Protoc. 2016, 2016, 087379. [Google Scholar] [CrossRef] [PubMed]
- Lujan-Hernandez, J.; Appasani, R.; Sullivan, K.; Siegel-Reamer, L.; Lalikos, J.F. Experimental In-Vivo Models Used in Fat Grafting Research for Volume Augmentation in Soft Tissue Reconstruction. Arch. Plast. Surg. 2017, 44, 361–369. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Tran, V.V.T.; Hong, K.Y.; Chang, H. Effect of Injectable Acellular Adipose Matrix on Soft Tissue Reconstruction in a Murine Model. Aesthetic Plast. Surg. 2024, 48, 2210–2219. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kim, S.E.; Kim, Y.J.; Kim, Y.W.; Cheon, Y.W. Comparison of Volume Retention and Biocompatibility of Acellular Dermal Matrix/Hyaluronic Acid Filler to Autologous Fat Grafts in a Mouse Model. Aesthetic Plast. Surg. 2020, 44, 986–992. [Google Scholar] [CrossRef]
- Yu, Y.; Zhang, H.; Sun, Z.; Wang, S.; Zhao, X.; Zhao, B.; Zhang, A. Exploring Preclinical Experiments with Different Fat Types for Autologous Fat Grafting. Aesthetic Plast. Surg. 2024, 48, 2330–2342. [Google Scholar] [CrossRef]
- Whitaker, R.; Hernaez-Estrada, B.; Hernandez, R.M.; Santos-Vizcaino, E.; Spiller, K.L. Immunomodulatory Biomaterials for Tissue Repair. Chem. Rev. 2021, 121, 11305–11335. [Google Scholar] [CrossRef]
- Yanez, M.; Blanchette, J.; Jabbarzadeh, E. Modulation of Inflammatory Response to Implanted Biomaterials Using Natural Compounds. Curr. Pharm. Des. 2017, 23, 6347–6357. [Google Scholar] [CrossRef]
- Stroumza, N.; Gauthier, N.; Senet, P.; Moguelet, P.; Barthelemy, R.N.; Atlan, M. Paradoxical Adipose Hypertrophy (PAH) After Cryolipolysis. Aesthetic Surg. J. 2018, 38, 411–417. [Google Scholar] [CrossRef]
- Song, J.H.; Kim, S.J.; Kwon, S.; Jeon, S.Y.; Park, S.E.; Choi, S.-J.; Oh, S.-Y.; Jeon, H.B.; Chang, J.W. Nervonic acid improves fat transplantation by promoting adipogenesis and angiogenesis. Int. J. Mol. Med. 2024, 54, 108. [Google Scholar] [CrossRef]
- Zhang, K.; Chen, X.; Zhang, P.; Liu, G. Perilipin2 is an Earlier Marker Than Perilipin1 for Identifying Adipocyte Regeneration in Fat Grafts. Aesthetic Surg. J. 2021, 41, NP646–NP652. [Google Scholar] [CrossRef]
- Goncharov, N.V.; Nadeev, A.D.; Jenkins, R.O.; Avdonin, P.V. Markers and Biomarkers of Endothelium: When Something Is Rotten in the State. Oxid. Med. Cell. Longev. 2017, 2017, 9759735. [Google Scholar] [CrossRef] [PubMed]
- Eto, H.; Kato, H.; Suga, H.; Aoi, N.; Doi, K.; Kuno, S.; Yoshimura, K. The fate of adipocytes after nonvascularized fat grafting: Evidence of early death and replacement of adipocytes. Plast Reconstr. Surg. 2012, 129, 1081–1092. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Role | Related Proteins Detected in Raw Tissue and alloClae | Related Proteins Detected Only in Raw Tissue | Related Proteins Detected Only in alloClae |
|---|---|---|---|
| Glycoproteins | LMNB1, LMNB2, LAMA2, LAMA3, LAMA4, LAMA5, LAMB1, LAMB2, LAMC1, EMILIN1, EMILIN2, FN1, FBN1, MYOC, THBS1, VIT, VWF, ECM1 | ||
| Collagens | COL1A1, COL3A1, COL4A1, COL5A1, COL6A1, COL12A1, COL14A1, COL15A1, COL18A1, COL28A1, COL1A2, COL4A2, COL5A2, COL6A2, COL6A3, COL6A6 | ||
| Proteoglycans | ASPN, BGN, DCN, HSPG2, LUM, OGN, PRELP, VCAN, FMOD, AGRN | ||
| ECM’s structural components | SERPINA1, LGALS1, PODN, MFAP4, VWF, VCAN, LTBP1, SPARCL1, ADAMTSL4, EFEMP1, EFEMP2, FBLN5, LTBP2, OGN, PRELP, LUM, TNXB, ECM1, COMP, MMP9, APOE, LGALS3BP, ANGPTL4, LPL, ASPN, BGN, ADAMTS15, TGM2, MYOC, VWA1, FBN1, TNC, TGFBI, COL1A1, COL3A1, COL4A1, COL5A1, COL6A1, COL12A1, COL14A1, COL15A1, COL18A1, COL28A1, COL1A2, COL4A2, COL5A2, COL6A2, COL6A3, COL6A6, SPARC, LOX1, CALR, ANXA2 | CCDC80 | ADAMTS15, HMCN2 |
| Angiogenesis | HMCN2, SPARC, PXDN, LOXL2, COL18A1, COL4A2, COL4A1, COL5A1, COL28A1, COL6A1, COL15A1, ANXA2, EFEMP2, FBLN4, THBS4, LAMB2, VWA1, FBN1, TNC, AMBP, ARP4, ADAMTSL4, FMOD, CASP | ||
| Anti-inflammatory activity | TGF-βR2, TGF-βR3, CD40, TNFSF13 | SMAD5, STAT5A | |
| Tissue remodeling | PDGFRα, PDGFRβ, TGF-βR2, TGF-βR3, FGF-2, GRN, MATN2, TIMP-1, TIMP-2, TIMP-3, MMP2, MMP9, ADAMTS4, ADAMTS15 | ||
| Adipogenesis | MYMX, AAAS, ABHD15, ACAT1, ACOX1, ACSL1, ACVR1C, ADAMTS15, ADAMTS4, ADGRF5,ADIPOQ, ADIRF, AGPAT2, AKR1C2, AKT1, ANP32E, APOE, ARHGEF15, ASPH, ATG7, C3, CAV1, CAV2, CD248, CD5L, CLDN5, CLMP, COL6A1, COMP, CTBP1, DARS1, DHRS7B, DOCK11, DSG2, DSP, ECHDC3, ENO1, ENPP2, FAP, FASN, FBN1, FTO, FZD4, GPD1, GPT2, GRN, HEPACAM, HEXA, HSPB7, IGF1R, INSR, JUP, LIPA, LMNA, LOXL2, LPL, LRP1, MFN2, MTARC1, OLFML3, PARP1, PGRMC2, PIK3CA, PIK3R1, PLAAT3, PLXNA4, PNPLA2, AMPK1, PRKAR2B, RARRES2, RHOA, RUVBL1, S100B, SCARA5, SLC25A25, SLC27A4, SNAPIN, SNX17, SPART, SPTLC2, SRPX2, SVEP1, TBL1XR1, TPP1, TRPM4, VIM, VIT |
| Integration to the Overlying Skin (0–3) | Integration to the Underlying Tissue (0–3) | Coloration of the Transplant (0–4) | Vascularization (0–4) | |
|---|---|---|---|---|
| 1 month | 1.86 | 1.86 | 0.71 | 1.86 |
| 3 months | 2.00 | 1.67 | 1.00 | 2.00 |
| 6 months | 2.00 | 2.00 | 1.00 | 2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fanniel, V.; Atawneh, I.; Savoie, J.; Izaguirre-Ramirez, M.; Marquez, J.; Khorsandi, C.; Hill, S. Advancing Soft Tissue Reconstruction with a Ready-to-Use Human Adipose Allograft. Bioengineering 2025, 12, 612. https://doi.org/10.3390/bioengineering12060612
Fanniel V, Atawneh I, Savoie J, Izaguirre-Ramirez M, Marquez J, Khorsandi C, Hill S. Advancing Soft Tissue Reconstruction with a Ready-to-Use Human Adipose Allograft. Bioengineering. 2025; 12(6):612. https://doi.org/10.3390/bioengineering12060612
Chicago/Turabian StyleFanniel, Victor, Ihab Atawneh, Jonathan Savoie, Michelle Izaguirre-Ramirez, Joanna Marquez, Christopher Khorsandi, and Shauna Hill. 2025. "Advancing Soft Tissue Reconstruction with a Ready-to-Use Human Adipose Allograft" Bioengineering 12, no. 6: 612. https://doi.org/10.3390/bioengineering12060612
APA StyleFanniel, V., Atawneh, I., Savoie, J., Izaguirre-Ramirez, M., Marquez, J., Khorsandi, C., & Hill, S. (2025). Advancing Soft Tissue Reconstruction with a Ready-to-Use Human Adipose Allograft. Bioengineering, 12(6), 612. https://doi.org/10.3390/bioengineering12060612