1. Introduction
Elevated atmospheric CO
2 levels represent a significant threat, driving global warming and resulting in rising sea levels, altered weather patterns, and profound impacts on ecosystems [
1,
2]. Renewable fuels are crucial for climate change mitigation by reducing greenhouse gas emissions from transportation and energy sectors, offering a sustainable alternative to fossil fuels [
3]. Gas fermentation is regarded as an emerging biotechnological tool with significant potential to contribute to the production of renewable chemicals and fuels [
4,
5,
6]. Fermentable gaseous substrates can be derived from CO
2 captured from off-gases in combination with H
2 produced via electrolysis of surplus electricity [
7]. Alternatively, syngas, a mixture of H
2, CO, and CO
2, can be utilized [
8]. Syngas is typically generated through the anoxic thermal decomposition of organic matter. The fermentative process serves as a biochemical alternative to the chemo–catalytic route, commonly known as Fischer–Tropsch synthesis [
9,
10,
11]. CO
2 fixation in the form of acetate represents only the initial step. The ultimate goal is to generate more valuable products with higher molecular weights, such as volatile fatty acids (VFAS) or alcohols [
12]. Chain elongation, i.e., the elongation of the carbon backbone, is the key biochemical process for achieving this objective. It is a growth-dependent anaerobic metabolism that combines acetate and ethanol to produce butyrate, hexanoate, and octanoate [
13]. While the microbial conversion of such gas mixtures has primarily been investigated using pure cultures, several other studies highlight the potential advantages of employing mixed microbial consortia. In nature, microbes rarely live in isolation; rather, they exist in diverse and complex communities. Microbes in these communities interact in various ways, often performing complex tasks that single species cannot accomplish independently. Hence, mixed microbial cultures are well suited to conduct carbon capture and chain elongation in a single bioreactor system [
14]. There are rapidly growing efforts to understand natural consortia and to engineer synthetic consortia for biotechnology applications [
15,
16]. Well-engineered microbial consortia involving several species can take advantage of the functions of individual microbes and their interactions to realize synergistic division of labor and more efficient utilization of biochemical substrates than monocultures. In addition to the ability to perform advanced biosynthetic tasks, microbial consortia exhibit appealing properties, including enhanced resilience and adaptive capacity, as well as the ability to operate in non-sterile conditions, all of which contribute to reducing the overall financial costs compared to pure culture-based processes [
17]. These benefits are particularly well suited to the requirements for successful industrial-scale syngas fermentation, which include flexible operation with various feedstocks, a robust process capable of handling disturbances arising from syngas quality variability and the presence of inhibitors, as well as improvements in productivity and selectivity, all while ensuring efficient utilization of syngas components at reduced capital and operating costs [
14].
In recent years, studies focusing on chain-elongating cultures have primarily concentrated on species from the class Clostridia [
18]. However, other potential chain elongators, such as
Megasphaera spp., have been isolated and warrant further investigation [
19]. Evidence suggesting that the strain
Megasphaera sueciensis may also be effectively employed for chain elongation has been presented by Batlle-Vilanova and co-workers [
19]. Additionally, batch pre-experiments conducted in our laboratory supported this hypothesis, as the formation of butyrate from acetate was observed. In a parallel experimental approach, continuous gas fermentation was established in trickle-bed reactors using mixed microbial cultures. The microbial cultures originally derived from a biogas plant and were adapted to the acetogenesis through long-term gas fermentation. The predominant product of the established mixed culture was acetate, a low-value compound, albeit produced in high concentrations of up to 700–800 mmol/L. Results from these experiments have been published previously [
20]. In the current paper, we report on the attempt to enhance the formation of short- to medium-chain products through the bioaugmentation of the chain-elongating strain
M. sueciensis and
C. carboxidivorans to an already established mixed culture.
3. Results and Discussion
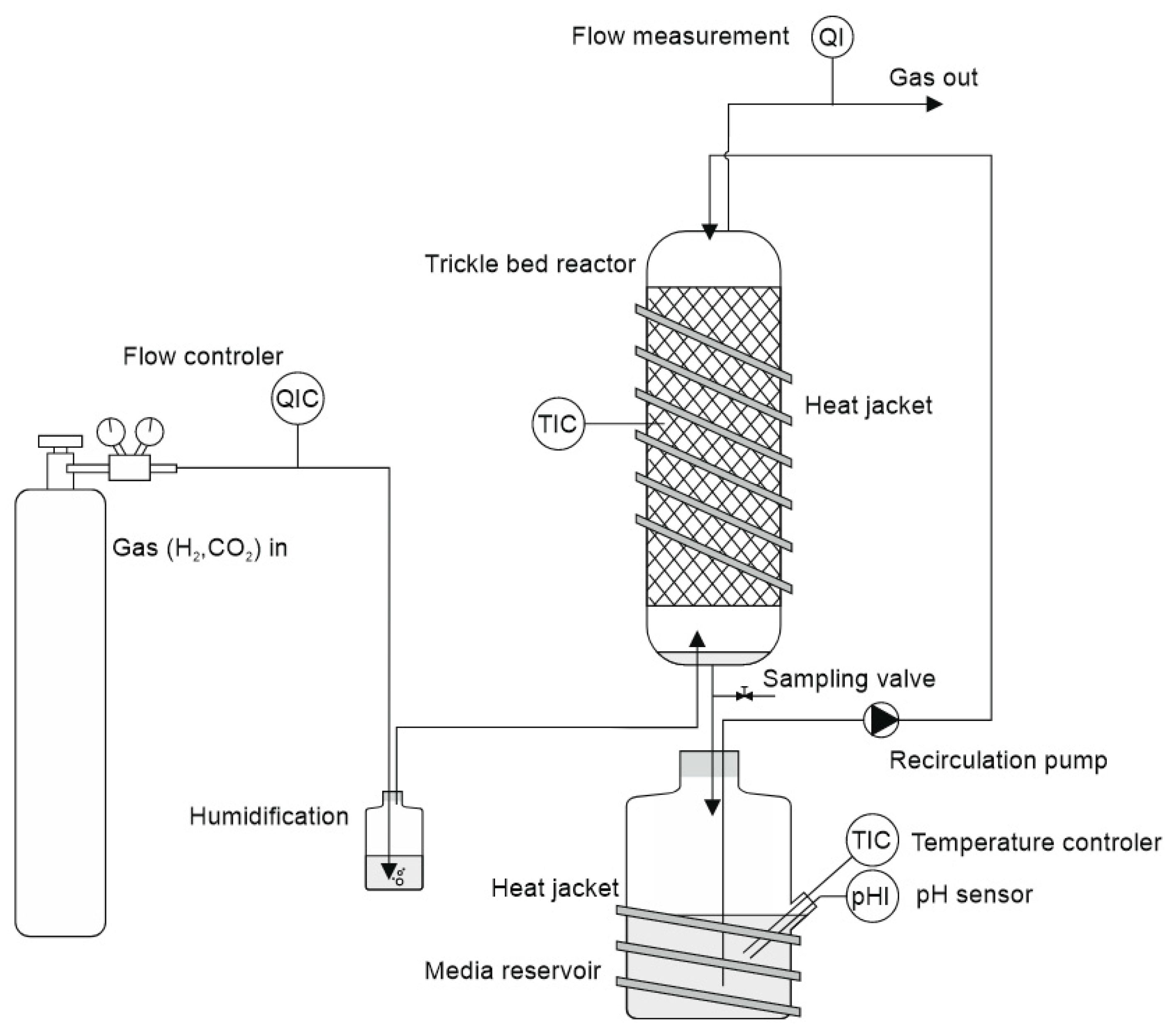
In the preceding syngas fermentation experiments mentioned above, a high acetate concentration of up to 40.9 g/L was achieved. However, the production of compounds with a longer chain length remained very low, with a maximum concentration of 0.93 g/L butyric acid, reaching only 2% of the main product, acetate [
20]. As explained, we tried to accelerate chain elongation in the continuous culture by the addition of the strain
M. sueciensis. For the current study, a new column was set up and inoculated with 50 mL (total media 400 mL) of a refrigerated mixed culture from the previous experiment. The intention was to revive the culture and promote attachment to the fresh carriers without allowing the development of a mature biofilm that could hinder the introduction of a new species. After an initial 7-day period, a freshly grown culture of
M. sueciensis was added to the chain-elongating reactor I (CHER I). As illustrated in
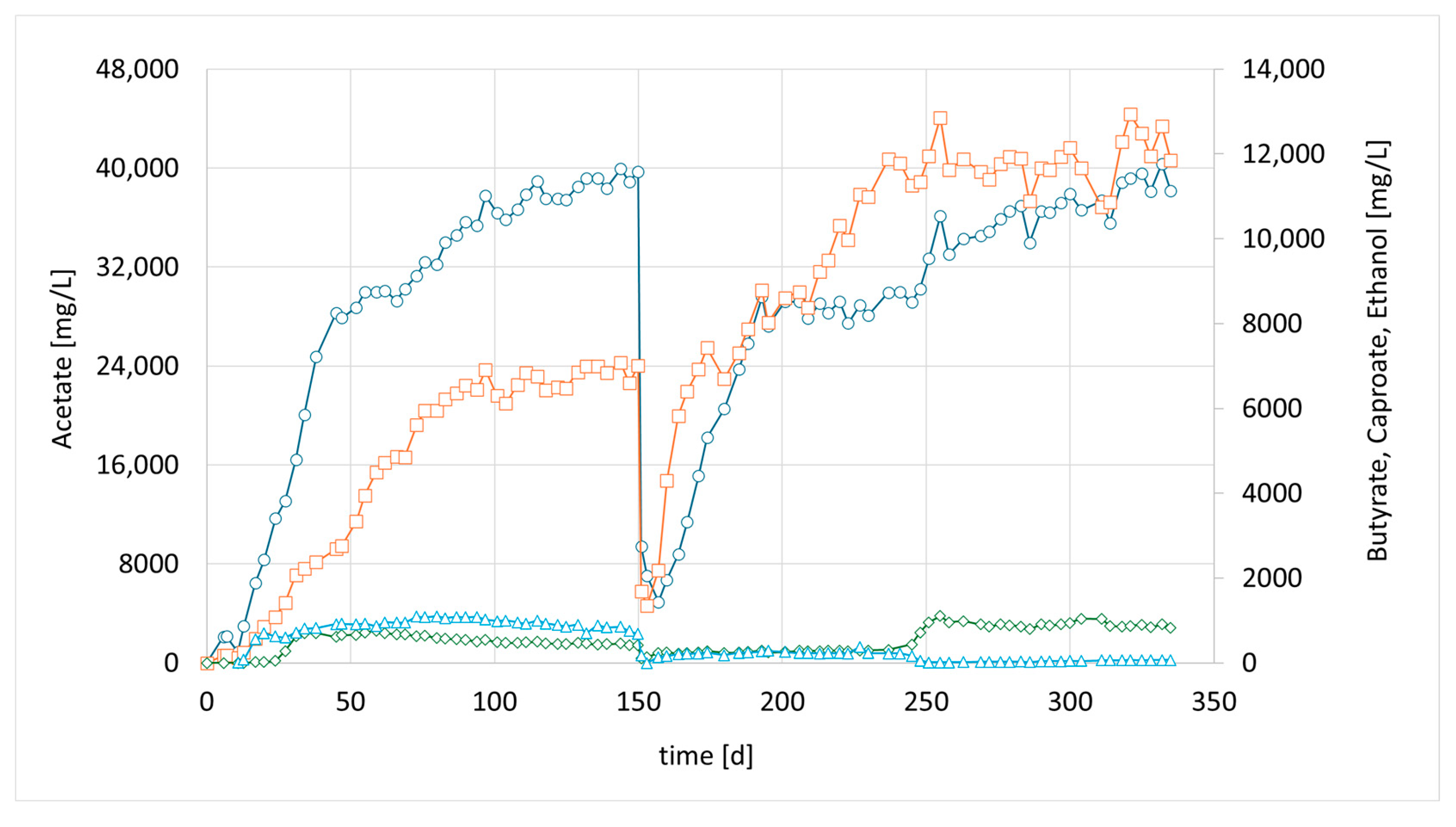
Figure 2, the formation of medium-chain acids increased quickly after adding the chain-elongating microorganism. Almost 10% of the total product was butyrate. Around 4 weeks after inoculation (day 35), even caproic acid was detected for the first time. The final levels in this first attempt were 7.2 g/L acetic acid 3.2 g/L butyric acid and 1.1 g/L caproic acid (
Figure 2, at 40 days). After that, a media swap was conducted to see if the performance of the established mixed culture remains stable. To our disappointment, the initial results did not seem to be reproducible. It took a while for the reason to be discovered: a crack in a three-way sampling valve, allowing small amounts of oxygen to enter the reactor. However, after sealing the leak, high butyrate concentrations were observed once again. In this second successful phase, even higher final butyrate concentrations were achieved compared to the previous phase. After around another 150 days, another media swap was conducted. Again, high concentrations were observed, and the share of butyrate and caproate was higher, even though overall concentrations were lower than in phase 2. The typical concentration of dissolved bacteria in the liquid medium, as determined by OD, ranged from approximately 0.8 to 1.4. Lower values were observed only at the initiation of each run; however, within a few days, the concentration stabilized within this range and exhibited slight fluctuations with a gradual increase over time. Notably, the OD did not significantly affect the product spectrum or concentration, suggesting that the majority of the active microorganisms were present within the biofilm. Biofilm formation was clearly visible; unfortunately, no methods were available to quantify the total bacterial biomass within the biofilm.
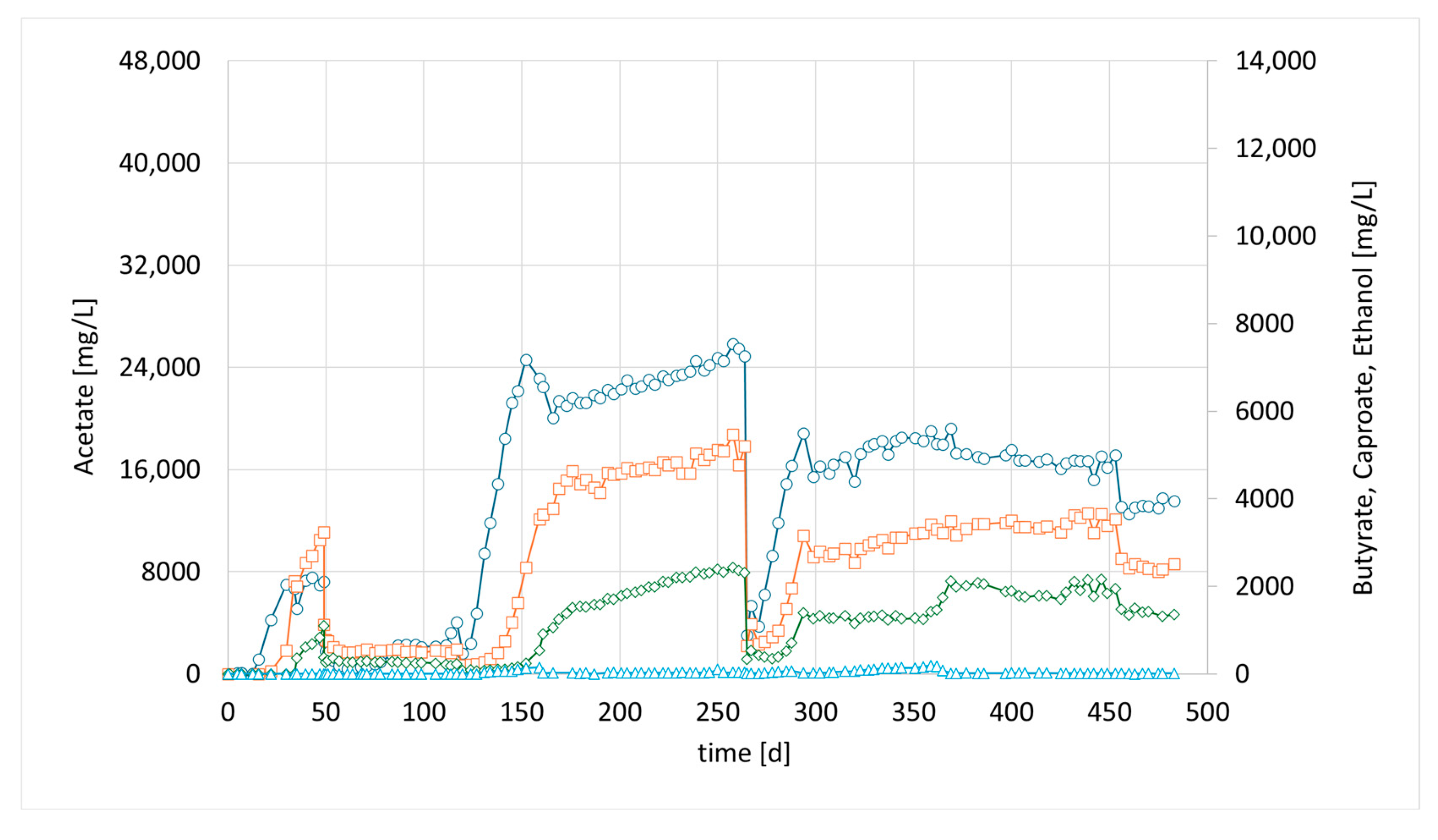
Meanwhile, more bioreactors became available, which have been used for bio methanation experiments before. CHER II was established with the aim of checking whether the modified mixed culture can be transferred to a new bioreactor and to confirm the observations made in the first setup. To inoculate this second column, 50 mL of the media of the first column (CHER I) was used. The results are illustrated in
Figure 3. The data showed good reproducibility of the earlier findings. After the first media swap, the ratio of butyrate to acetate shifted from 1:5 [1:4] to 1:2, reaching acetate values of 21 g/L and butyrate levels of up to 11 g/L, while caproate reached a concentration of 2 g/L. An overview of the best results achieved in the individual reactors is provided in
Table 6. To contextualize the concentrations achieved in the experiments, values of up to 43 g/L of acetate have been reported in pure cultures utilizing syngas [
25]. For mixed consortia, only lower concentrations have been observed, ranging from 2 to 15 g/L of acetate [
26], with up to 2.3 g/L of butyrate [
27].
While the observation of medium-chain metabolites was encouraging, the next goal was the formation of the carboxylic acids’ respective alcohols. Carboxylates are one of the major platform chemicals for the generation of bioalcohols, the main drop-in fuels to substitute petroleum gasoline [
28]. Typically, the added value of the alcohols increases with the number of carbon atoms. For example, hexanoate/hexanol has twice the market value of ethanol [
29]. Based on current scientific understanding, alcohol fermentation is facilitated at lower pH levels [
21,
30,
31]. According to a recent review on syngas fermentation, studies on various
Clostridium species revealed that a low pH (4.5–5.0) favors solventogenesis, whereas a higher pH range (5.0–6.0) promotes acetogenesis [
32].
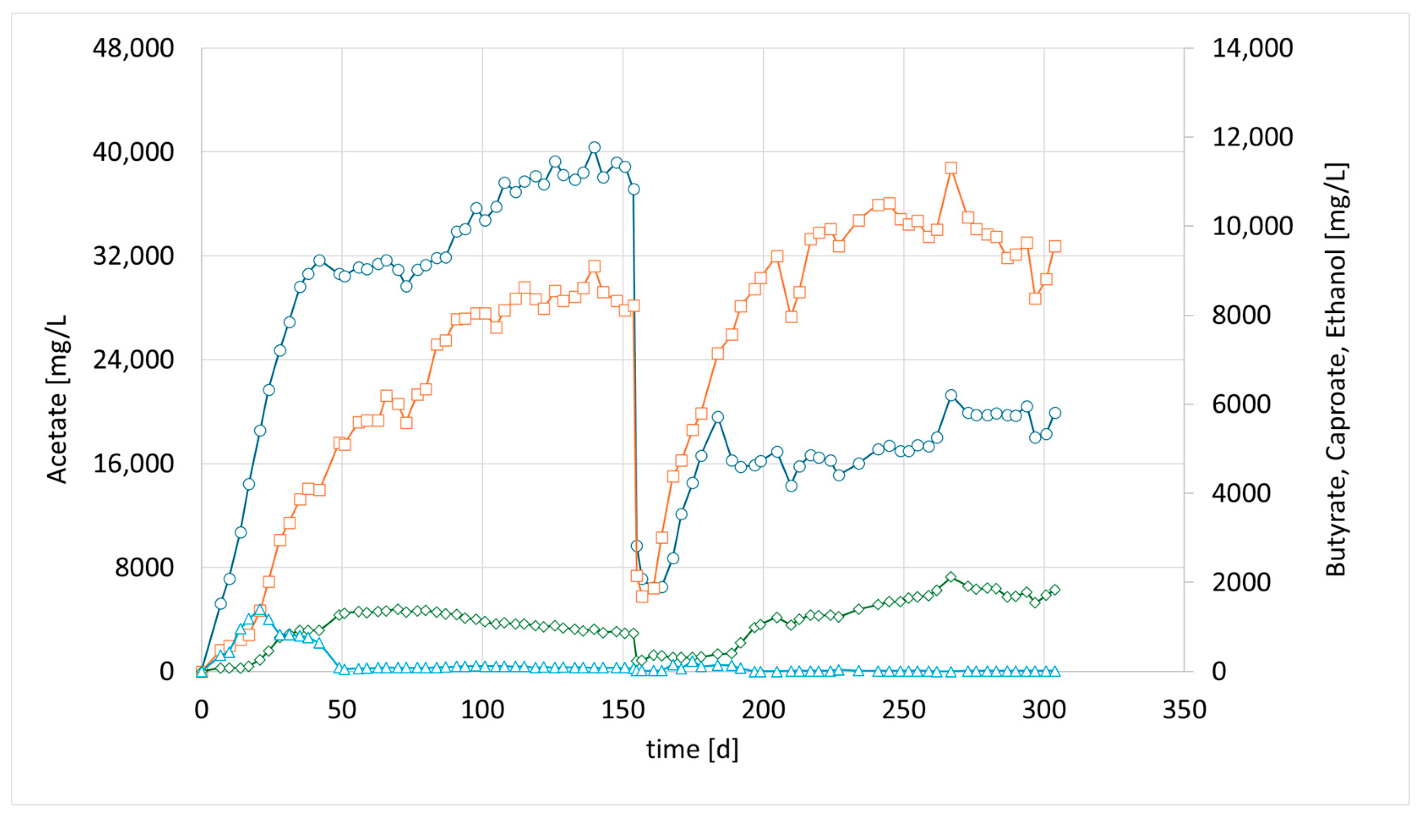
Accordingly, in a third parallel experiment (CHER III), the impact of a lower pH was investigated. The pH set point of this unit was 0.5 units lower than in CHER II. An attempt was made to avoid a too low pH range as the optimum of
M. sueciensis lies at 7.0. Typical endpoints before readjustment of pH were 5.0–5.2. As shown in
Figure 4, the lowering of the pH led to elevated EtOH values that were five times higher than those observed in CHER II. However, no butanol formation was observed during these trials. CHER III best product ratios were 37.4 g/L acetate, 10 g/L butyrate and 1.6 g/L caproate at around day 255 (
Figure 4). The highest measured ethanol content was 3.8 g/L. Ethanol production decreased after the first media swap, leading to an increase in butyrate concentration and an improved acetate-to-butyrate ratio.
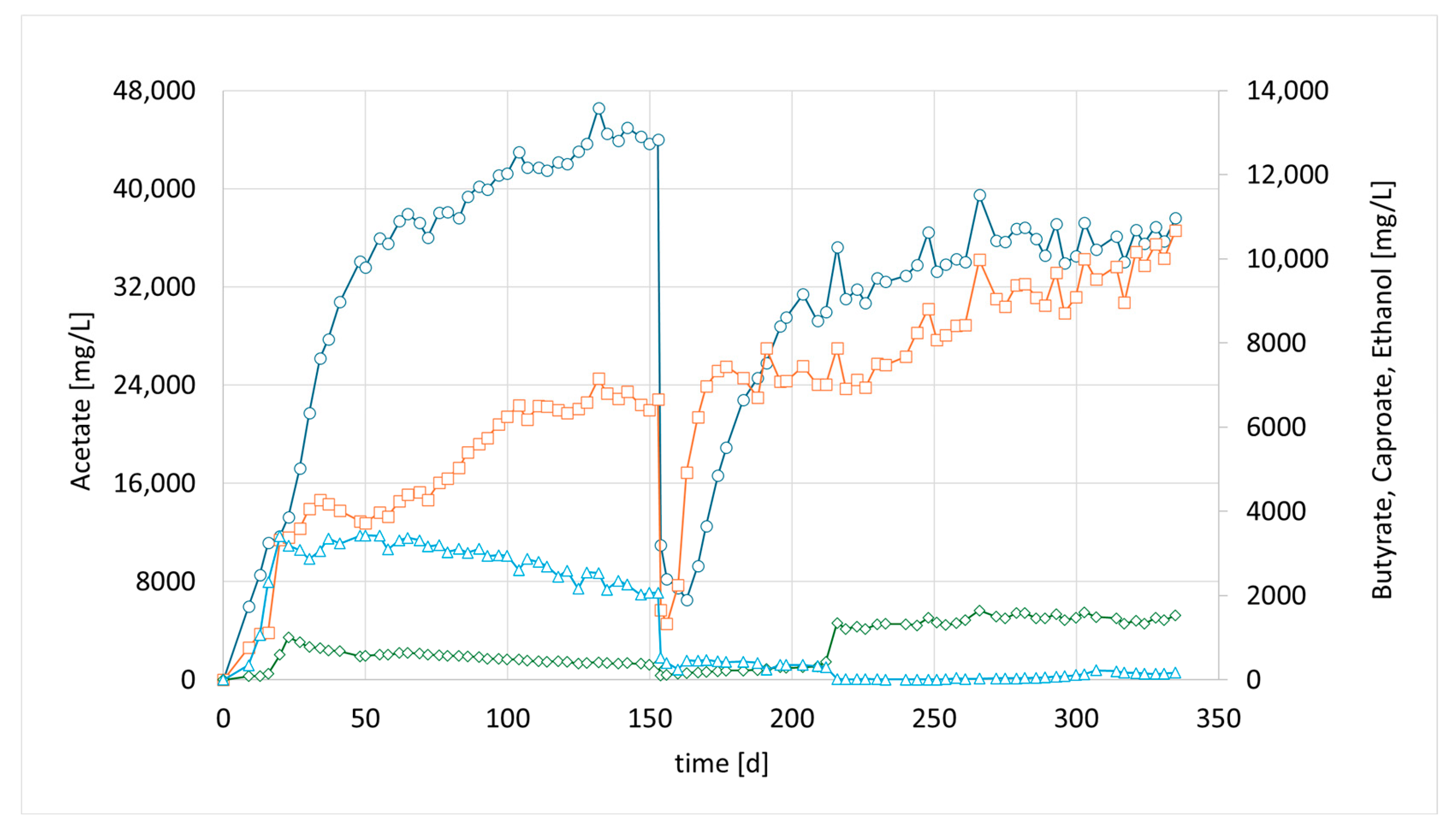
The final attempt, CHER IV; examined the augmentation of a second strain, the well-studied autotrophic species
C. carboxidivorans. Similar to CHER II & III, this reactor was inoculated with a mixed culture from CHER I. After 7 days, 50 mL of a separately cultivated
C. carboxidivorans pre-culture was added. The goal was to determine whether alcohol formation, particularly butanol production, could be enhanced. Several investigations have reported that the augmented strain is capable of producing carboxylic acids up to C6 and reducing them to their corresponding alcohols, including n-hexanol [
23]. The formed products resulted in the concentrations of 36.1 g/L acetate, 12.1 g/L butyrate, and 1.0 g/L caproate (
Figure 5, day 255). Alcoholic compounds were below 100 mg/L.
During this period, it was necessary to develop an analytical method for the detection and quantification of longer chain carboxylic acids/alcohols, as HPLC analysis revealed the presence of additional peaks. Due to the long retention time at the given analytical conditions, it was not feasible to retain this analysis for such products. Alternatively, the GC-MS method provided in Materials and Methods was developed. In fact, the formation of n-hexanol in CHER IV was discovered during a later stage of this fermentation (10 mg/L, around day 270). However, the concentrations of bio-alcohols in the media remained very low. As discussed further down, this low formation of hexanol cannot be clearly attributed to the augmentation of C. carboxidivorans, since the metataxonomic analysis showed no significant increase of the clostridia family. Overall, the metabolite profile closely resembled the performance of CHER II or CHER III, and bioaugmentation had no significant impact on the product spectrum. The new analytical method was also used to reanalyze the caprylic acid concentration in individual frozen samples taken from the reactors. The caprylic acid concentration reached 0.5 g/L in CHER I and 0.1 g/L in CHER II and CHER III, while no caprylic acid was detected in CHER IV.
Metataxonomic Data
Figure 6 shows the composition of the consortium that was used as a starter culture for the reactors (data from Steger et al. [
20]). The genus
Megasphaera could not be detected in the inoculum, as demonstrated in
Figure 6 and
Supplementary Figure S1.
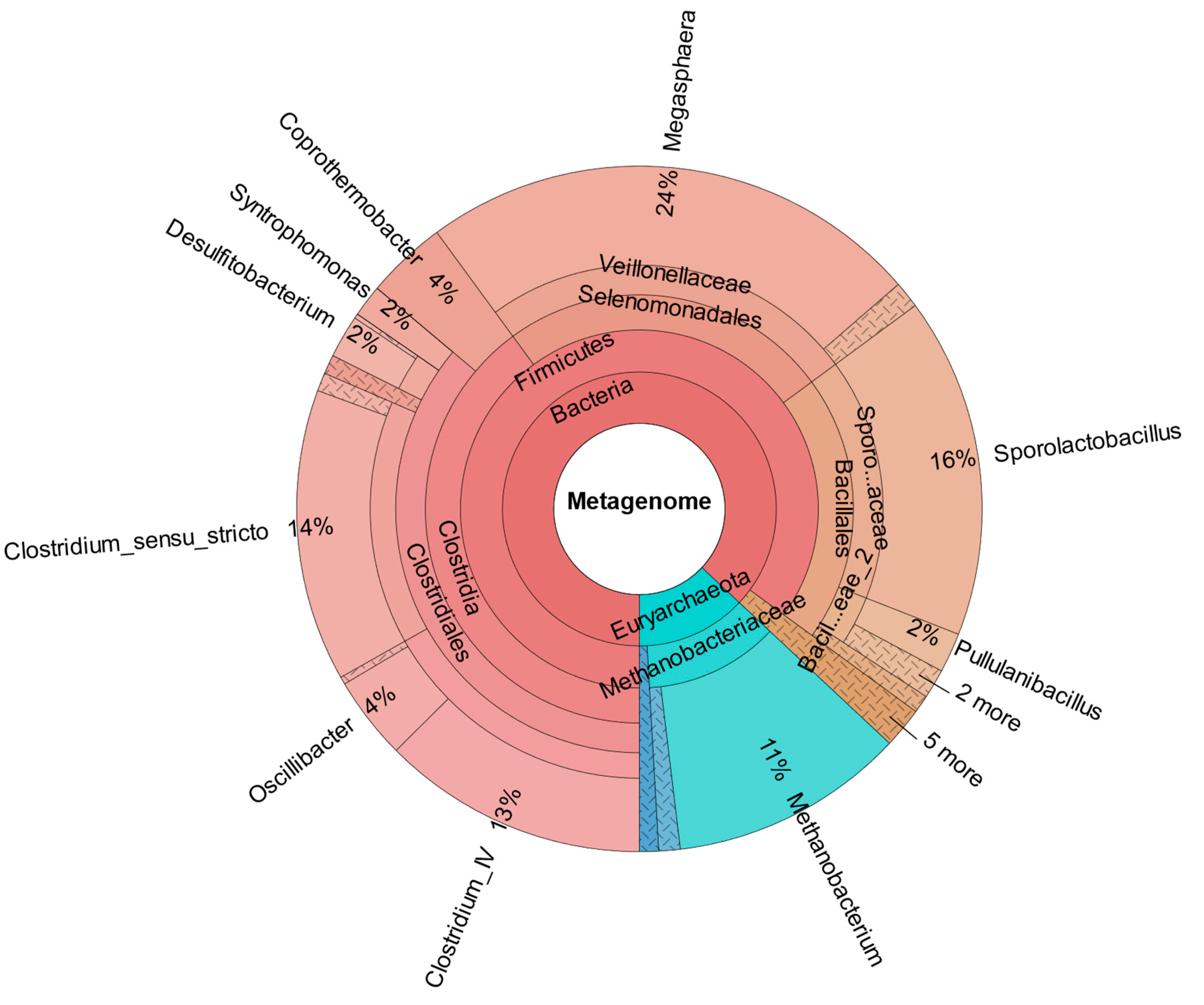
In comparison,
Figure 7 shows the metataxonomic results from CHER I. The
Supplementary Information provide insight in the generation of metataxonomic data for CHER I-IV, including used primer sequence and quality score. The sample was taken at day 315. As shown,
M. sueciensis is present in a much higher share (7%) and is also the dominant member of
Veillonellales, even though there was a time span of 144 days between inoculation with this
M. sueciensis and sampling. This result proves that it was possible to establish the introduced species as a permanent microbial member of the mixed population. It should be underlined that all reactors were operated under non-sterile auto-selective conditions with one notable exception: the addition of the inhibitor BES in order to suppress the growth of methanogenic archaea.
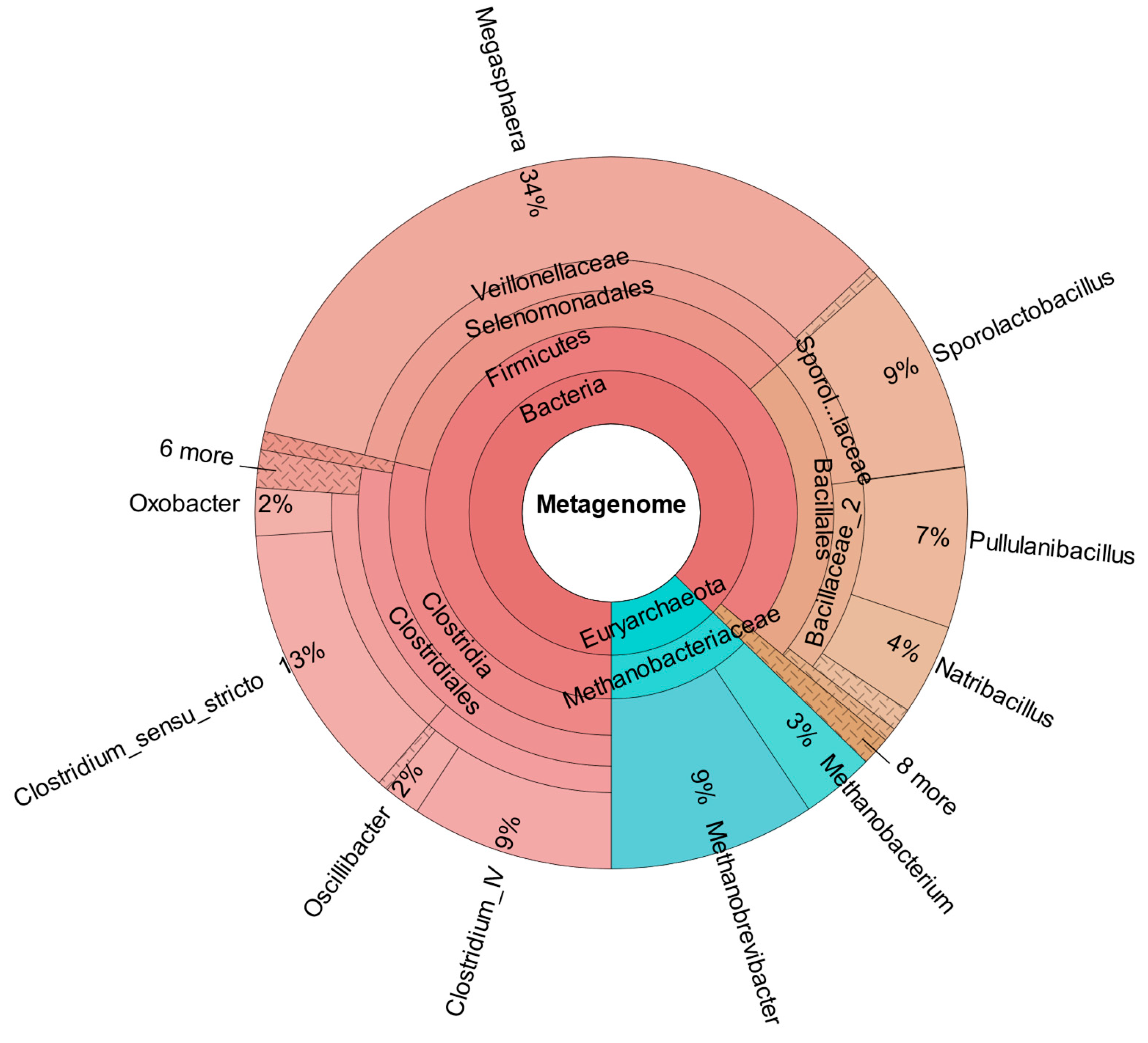
As confirmed by
Figure 8, which shows the metataxonomic data of CHER II, the establishment of
M. sueciensis is not only long lasting but it even increased in percentage proportion (24%). Unexpectedly, in this reactor, a certain share of archaeal species was also found. This is likely a remnant of the previous experiments on biomethanation of CO
2 and H
2. The bioreactors were flushed, and the medium was exchanged; however, sterilization was not possible. We expected that the exposure to O
2 during reactor manipulation to prepare for the new experiment in combination with the presence of BES was sufficient to eradicate methanogenic species. Also, the permanent operation at pH levels below 6.0 must be considered. It is assumed that methanogenic species survived in niches and deeper layers of the biofilm that were not removed. Later, metataxonomic analysis confirmed that archaeal organisms declined over time. In CHER III (
Figure 9), the reactor with the slightly lower pH, the proportion of
M. sueciensis was actually the highest, despite its reported growth optimum being at neutral pH. It constitutes 96% of the identified members of the
Veillonellaceae family.
In contrast to the results obtained, CHER IV’s metataxonomic data (
Figure 10) did not indicate the successful introduction of
C. carboxidivorans. There is a certain percentage of the parent genus
Clostridium senso stricto (13%), although it is not significantly different from the other reactors (CHER I: 5%; CHER II: 14%, CHER III: 11%). This aligns with the low impact on product formation of this second bioaugmentation attempt.
Generally, the concept of co-cultivation of two or more defined species already has some tradition in biotechnology, e.g., co-cultivation is an established strategy for the industrial-scale fermentation of Vitamin B12 [
33]. Co-cultivation has also been discussed for the enhancement of tertiary metabolite production [
34]. In addition, mixed-culture fermentations utilizing enriched natural communities are a well-known strategy. In addition to its traditional application in the food industry (e.g., sourdough, yoghurt) as well as in wastewater treatment or biogas production, mixed microbial cultures have been proposed for a variety of applications in industrial biotechnology. This includes bioethanol, biohydrogen, butanol and 1,4 propanediol [
35]. Several laboratory studies confirm the value of mixed microbial consortia in syngas fermentation [
26,
36,
37]. Moreover, the authors of the abovementioned investigations [
21], which closely resemble the experiments conducted here, emphasize the robustness of the system and high reproducibility of the results obtained with trickle-bed reactors.
Bioaugmentation integrates both approaches, enhancing a natural and well-adapted consortium by introducing species that provide complementary functions. Most full-scale applications are found in agriculture (particularly the introduction of nitrogen-fixing microorganisms) and soil remediation (amplifying the degradation of specific pollutants) [
38]. However, there are also several examples where bioaugmentation has been used to enhance product formation. For example, Pozytek et al. list several different examples where strain addition has been applied to improve anaerobic digestion reactor stability [
39] and/or biogas formation rate [
40]. The generated data support the validity of this approach. The addition of
M. sueciensis to an existing acetogenic mixed culture shifted the product spectrum from almost exclusively acetate into higher value compounds, such as butyrate, caproate and caprylate. The experiment most closely related to the current investigation, as reported in the literature, involves the bioaugmentation of a mixed bacterial culture with
Clostridium kluyveri for the microbial production of caproate from mixed carbon sources [
41]. However, in the mentioned study, the addition of
C. kluyveri, an efficient caproic acid producer, failed to positively impact product formation. In a similar manner, our attempt to introduce a new
Clostridium species was unsuccessful. It should be emphasized that the cultivation of the pure culture was conducted in the same growth medium and under the same gas composition as used in the trickle-bed reactor, and no indication of impaired growth was observed. One potential explanation for the failure is the presence of a relatively high number of closely related and likely well-adapted clostridial species, as revealed by the taxonomic analysis. These organisms presumably occupy the same ecological niche, which may hinder the establishment of a bacterium with a similar genetic makeup.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}