
Shark Cartilage-Derived Anti-Angiogenic Peptide Inhibits Corneal Neovascularization

 and
and
Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Proliferation Assay
2.2. Cell Migration Assay
2.3. Transwell Assay
2.4. Matrigel Tube Formation Assay
2.5. Matrigel Plug Assay
2.6. Corneal Alkali Injury Model
2.7. Immunohistochemistry
2.8. Real-Time Fluorescence Quantitative PCR
2.9. Statistical Analysis
3. Results
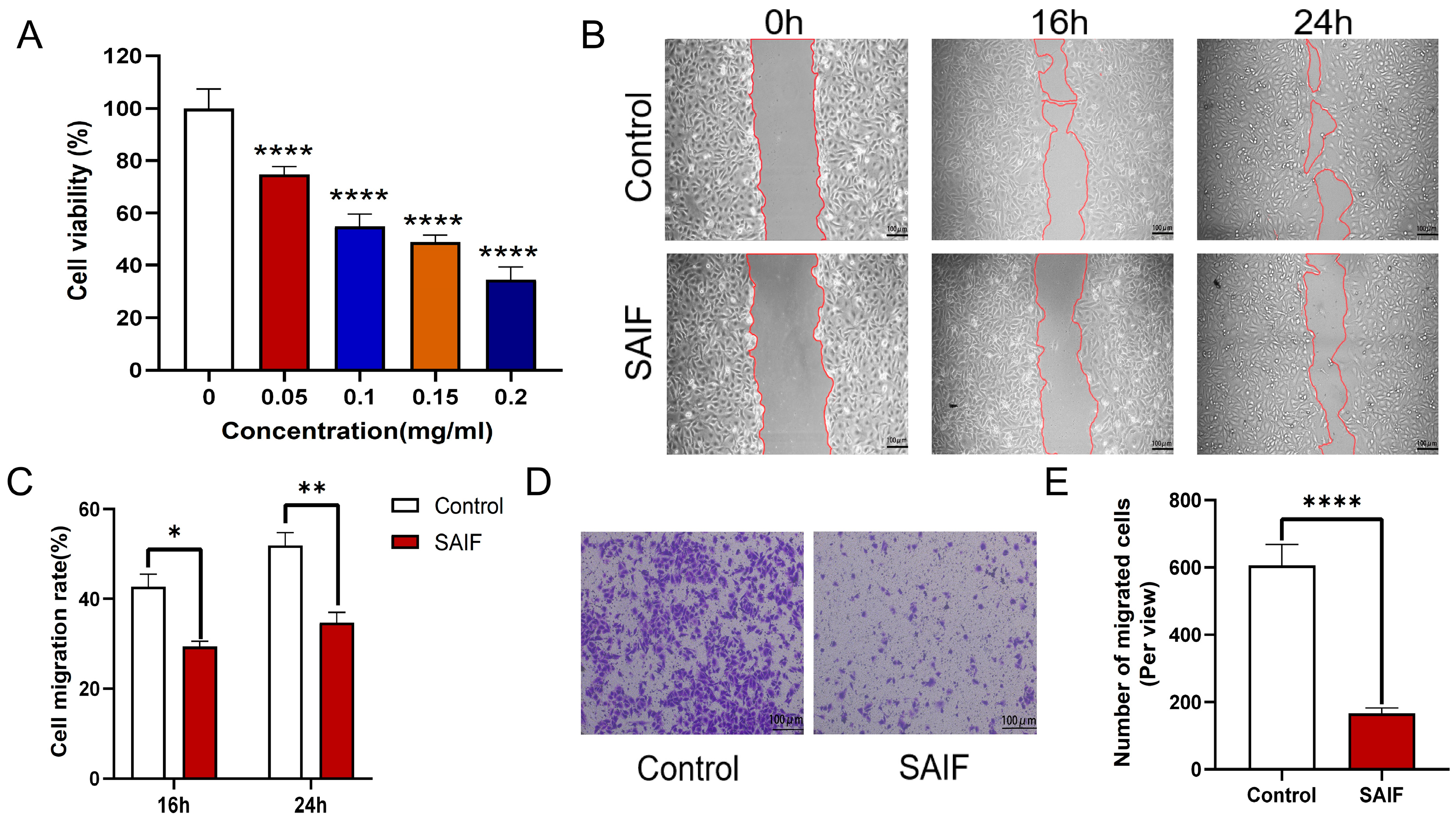
3.1. SAIF Inhibits the Proliferation and Migration of HUVEC Cells
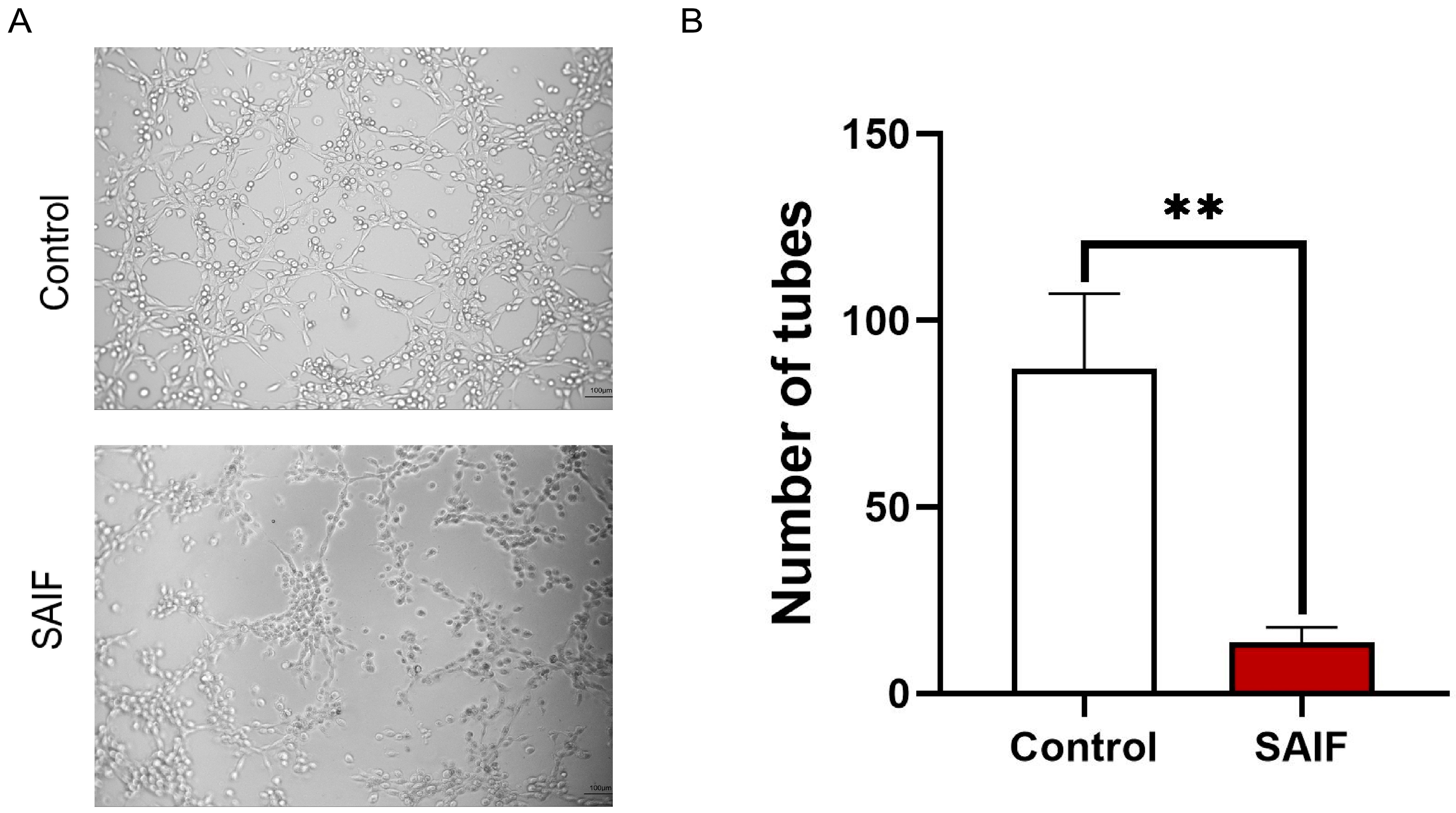
3.2. SAIF Inhibits the Tube Formation of HUVEC Cells
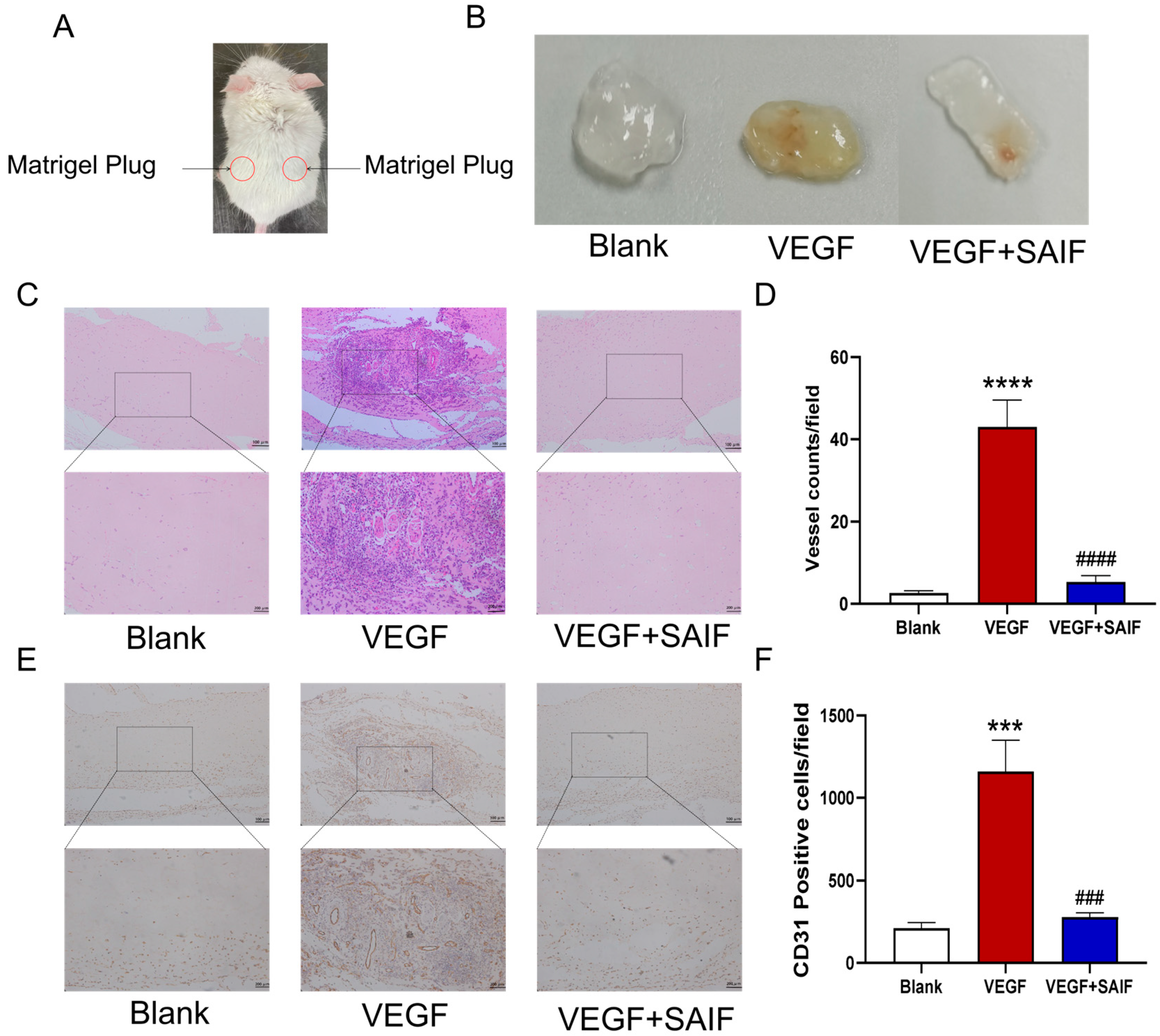
3.3. SAIF Inhibits Vascular Neovascularization of Matrigel Plugs in Mice
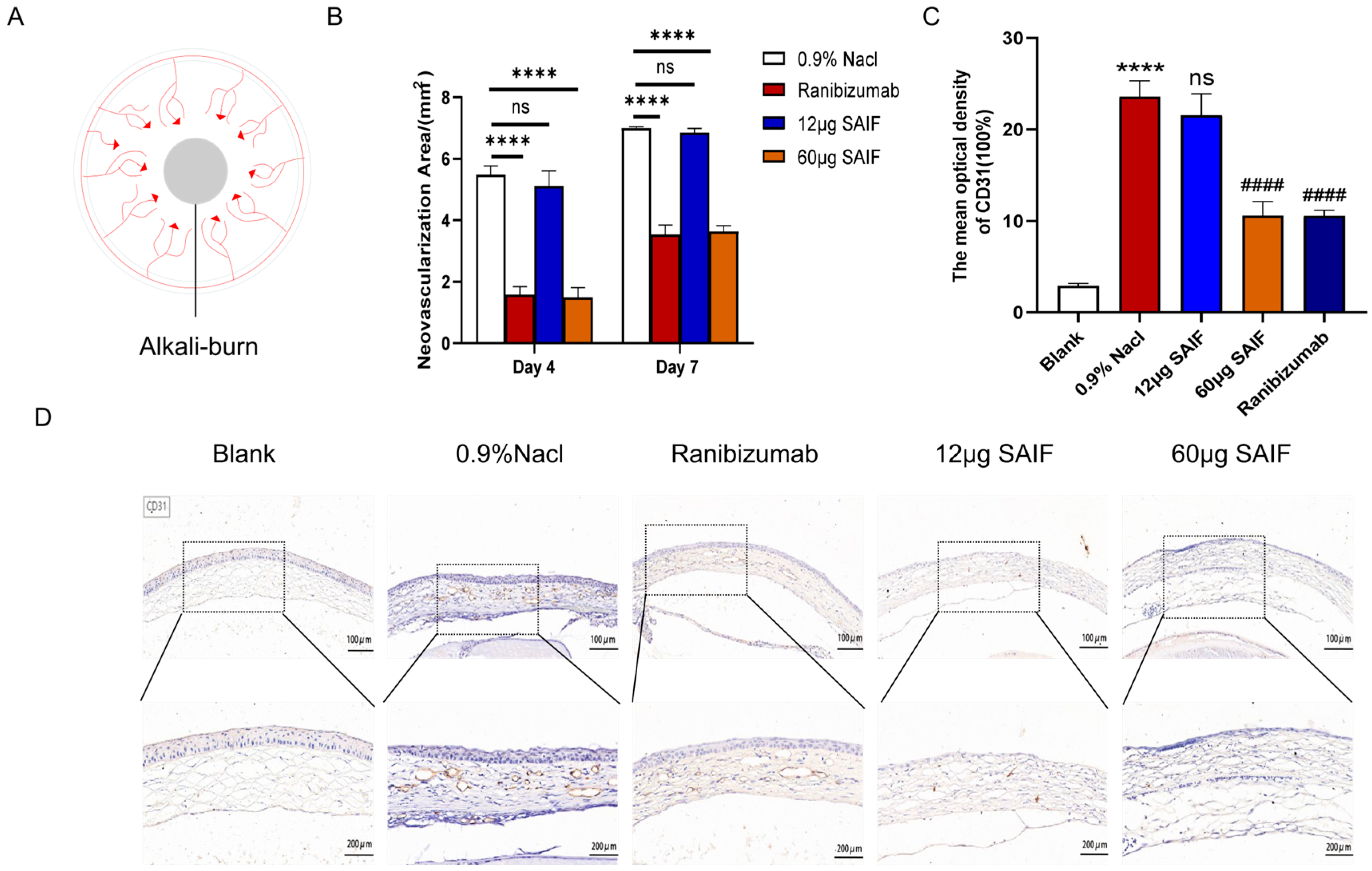
3.4. SAIF Inhibits Corneal Neovascularization in Mice
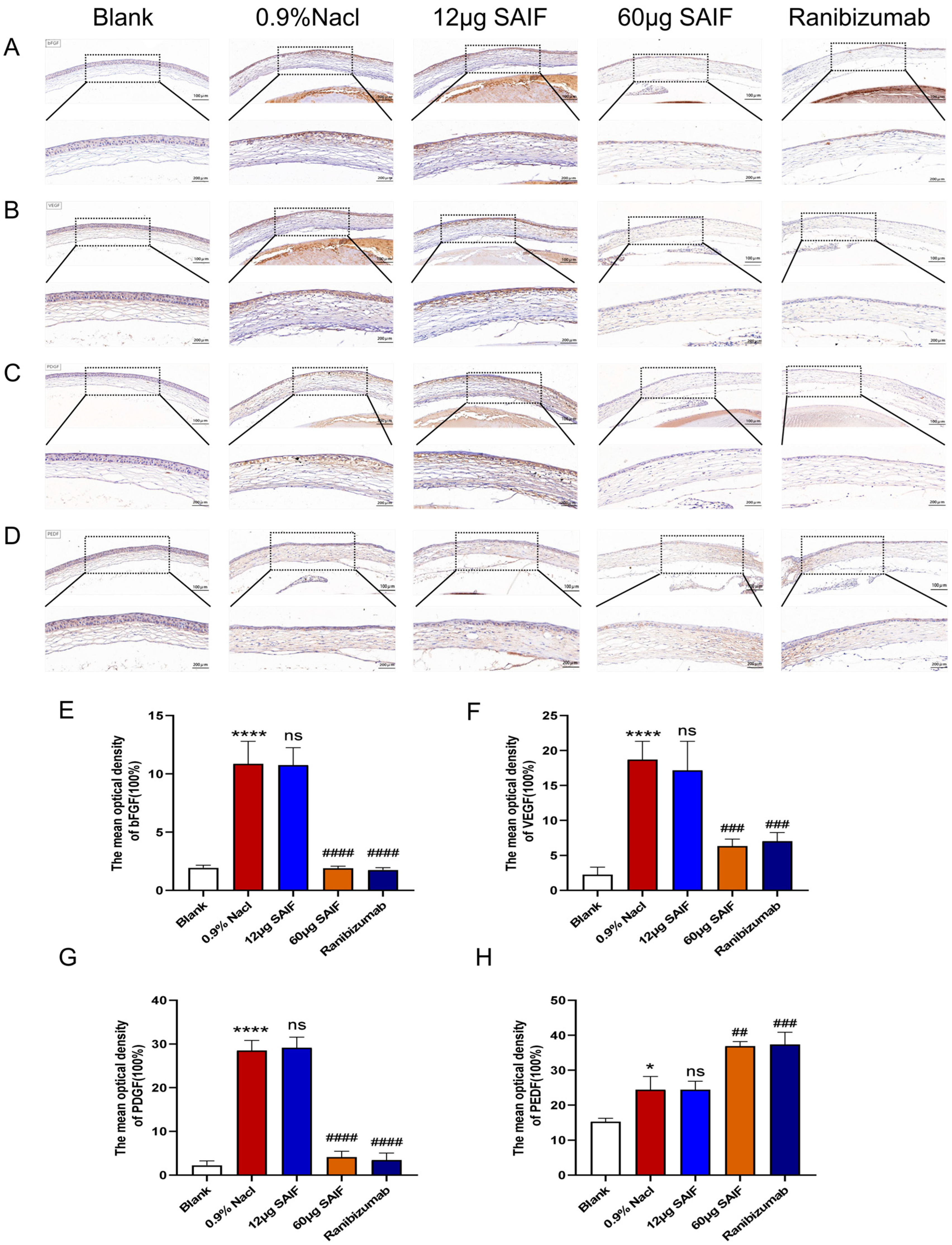
3.5. SAIF Affects the Expression of Angiogenic Factor in Mouse Corneal
3.6. SAIF Affects the Expression of MMPs in Mouse Corneal
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nicholas, M.P.; Mysore, N. Corneal neovascularization. Exp. Eye Res. 2021, 202, 108363. [Google Scholar] [CrossRef] [PubMed]
- Azar, D.T.; Chang, J.-H.; Han, K.Y. Wound healing after keratorefractive surgery: Review of biological and optical considerations. Cornea 2012, 31, S9. [Google Scholar] [CrossRef]
- Bock, F.; Cursiefen, C. Corneal Angiogenesis and Lymphangiogenesis. Found. Corneal Dis. Past Present Future 2020, 17, 249–262. [Google Scholar]
- Bock, F.; Maruyama, K.; Regenfuss, B.; Hos, D.; Steven, P.; Heindl, L.M.; Cursiefen, C. Novel anti (lymph) angiogenic treatment strategies for corneal and ocular surface diseases. Prog. Retin. Eye Res. 2013, 34, 89–124. [Google Scholar] [CrossRef] [PubMed]
- Maddula, S.; Davis, D.K.; Maddula, S.; Burrow, M.K.; Ambati, B.K. Horizons in therapy for corneal angiogenesis. Ophthalmology 2011, 118, 591–599. [Google Scholar] [CrossRef]
- Feizi, S.; Azari, A.A.; Safapour, S. Therapeutic approaches for corneal neovascularization. Eye Vis. 2017, 4, 1–10. [Google Scholar] [CrossRef]
- Papathanassiou, M.; Theodoropoulou, S.; Analitis, A.; Tzonou, A.; Theodossiadis, P.G. Vascular endothelial growth factor inhibitors for treatment of corneal neovascularization: A meta-analysis. Cornea 2013, 32, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Ellenberg, D.; Azar, D.T.; Hallak, J.A.; Tobaigy, F.; Han, K.Y.; Jain, S.; Zhou, Z.; Chang, J.-H. Novel aspects of corneal angiogenic and lymphangiogenic privilege. Prog. Retin. Eye Res. 2010, 29, 208–248. [Google Scholar] [CrossRef]
- Sulochana, K.; Ge, R. Developing antiangiogenic peptide drugs for angiogenesis-related diseases. Curr. Pharm. Des. 2007, 13, 2074–2086. [Google Scholar] [CrossRef]
- Kazemnejad, S.; Khanmohammadi, M.; Baheiraei, N.; Arasteh, S. Current state of cartilage tissue engineering using nanofibrous scaffolds and stem cells. Avicenna J. Med. Biotechnol. 2017, 9, 50. [Google Scholar]
- Smith, L.J.; Elliott, D.M. Formation of lamellar cross bridges in the annulus fibrosus of the intervertebral disc is a consequence of vascular regression. Matrix Biol. 2011, 30, 267–274. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Isolation of a cartilage factor that inhibits tumor neovascularisation. Science 1976, 193, 70–71. [Google Scholar]
- Cho, J.; Kim, Y. Sharks: A potential source of antiangiogenic factors and tumor treatments. Mar. Biotechnol. 2002, 4, 521–525. [Google Scholar] [CrossRef]
- Walker, M. Why shark cartilage should succeed against cancer and other pathologies. Townsend Lett. Dr. 1991, 5, 847–854. [Google Scholar]
- Sheu, J.R.; Fu, C.C.; Tsai, M.L.; Chung, W.J. Effect of U-995, a potent shark cartilage-derived angiogenesis inhibitor, on anti-angiogenesis and anti-tumor activities. Anticancer Res. 1998, 18, 4435–4441. [Google Scholar] [PubMed]
- Gonzalez, R.P.; Soares, F.d.S.D.; Farias, R.F.; Pessoa, C.; Leyva, A.; de Barros Viana, G.S.; Moraes, M.O. Demonstration of inhibitory effect of oral shark cartilage on basic fibroblast growth factor-induced angiogenesis in the rabbit cornea. Biol. Pharm. Bull. 2001, 24, 151–154. [Google Scholar] [CrossRef] [PubMed]
- Xie, Q.; Yao, S.; Chen, X.; Xu, L.; Peng, W.; Zhang, L.; Zhang, Q.; Liang, X.-F.; Hong, A. A polypeptide from shark troponin I can inhibit angiogenesis and tumor growth. Mol. Biol. Rep. 2012, 39, 1493–1501. [Google Scholar] [CrossRef] [PubMed]
- Kern, B.E.; Balcom, J.H.; Antoniu, B.A.; Warshaw, A.L.; Castillo, C.F.-d. Troponin I peptide (Glu94-Leu123), a cartilage-derived angiogenesis inhibitor: In vitro and in vivo effects on human endothelial cells and on pancreatic cancer. J. Gastrointest. Surg. 2003, 7, 961–969. [Google Scholar] [CrossRef] [PubMed]
- Oikawa, T.; Ashino-Fuse, H.; Shimamura, M.; Koide, U.; Iwaguchi, T. A novel angiogenic inhibitor derived from Japanese shark cartilage (I). Extraction and estimation of inhibitory activities toward tumor and embryonic angiogenesis. Cancer Lett. 1990, 51, 181–186. [Google Scholar] [CrossRef]
- Xie, Q.L.; Wang, Y.Y.; Chen, X.J.; Li, G.G.; Zhang, L.; Liang, X.F. Molecular cloning, expression and anti-angiogenesis activity of TnI from whitespotted bambooshark (Chiloscyllium plagiosum Bennett). Chin. J. Pharm. Biotechnol. 2013, 20, 110–114. [Google Scholar]
- Zuazo-Gaztelu, I.; Casanovas, O.J.F.I.O. Unraveling the role of angiogenesis in cancer ecosystems. Front. Oncol. 2018, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- Page-McCaw, A.; Ewald, A.J.; Werb, Z. Matrix metalloproteinases and the regulation of tissue remodelling. Nat. Rev. Mol. Cell Biol. 2007, 8, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.-C.; Chang, J.-H.; Jain, S.; Gabison, E.E.; Kure, T.; Kato, T.; Fukai, N.; Azar, D.T. Matrilysin cleavage of corneal collagen type XVIII NC1 domain and generation of a 28-kDa fragment. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2517–2524. [Google Scholar]
- Nagase, H.; Visse, R.; Murphy, G. Structure and function of matrix metalloproteinases and TIMPs. Cardiovasc. Res. 2006, 69, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Whitcher, J.P.; Srinivasan, M.; Upadhyay, M.P. Corneal blindness: A global perspective. Bull. World Health Organ. 2001, 79, 214–221. [Google Scholar] [PubMed]
- Keating, A.M.; Jacobs, D.S. Anti-VEGF treatment of corneal neovascularization. Ocul. Surf. 2011, 9, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Parker, B.C.; Engels, M.; Annala, M.; Zhang, W. Emergence of FGFR family gene fusions as therapeutic targets in a wide spectrum of solid tumours. J. Pathol. 2014, 232, 4–15. [Google Scholar] [CrossRef]
- Gan, L.; Fagerholm, P.; Palmblad, J. Expression of basic fibroblast growth factor in rabbit corneal alkali wounds in the presence and absence of granulocytes. Acta Ophthalmol. Scand. 2005, 83, 374–378. [Google Scholar] [CrossRef]
- Chen, L.; Wu, H.; Ren, C.; Liu, G.; Zhang, W.; Liu, W.; Lu, P. Inhibition of PDGF-BB reduces alkali-induced corneal neovascularization in mice. Mol. Med. Rep. 2021, 23, 1–11. [Google Scholar] [CrossRef]
- Marçola, M.; Rodrigues, C.E. Endothelial progenitor cells in tumor angiogenesis: Another brick in the wall. Stem Cells Int. 2015, 2015, 832649. [Google Scholar] [CrossRef]
- Cook, K.M.; Figg, W.D. Angiogenesis inhibitors: Current strategies and future prospects. CA A Cancer J. Clin. 2010, 60, 222–243. [Google Scholar] [CrossRef] [PubMed]
- Meyer, C.; Notari, L.; Becerra, S.P. Mapping the type I collagen-binding site on pigment epithelium-derived factor: Implications for its antiangiogenic activity. J. Biol. Chem. 2002, 277, 45400–45407. [Google Scholar] [CrossRef] [PubMed]
- Becerra, S.P.; Notario, V. The effects of PEDF on cancer biology: Mechanisms of action and therapeutic potential. Nat. Rev. Cancer 2013, 13, 258–271. [Google Scholar] [CrossRef] [PubMed]
- Sacks, D.; Baxter, B.; Campbell, B.C.V.; Carpenter, J.S.; Cognard, C.; Dippel, D.; Eesa, M.; Fischer, U.; Hausegger, K.; Hirsch, J.A.; et al. Multisociety Consensus Quality Improvement Revised Consensus Statement for Endovascular Therapy of Acute Ischemic Stroke. Int. J. Stroke Off. J. Int. Stroke Soc. 2018, 13, 612–632. [Google Scholar] [CrossRef] [PubMed]
- Azar, D.T. Corneal angiogenic privilege: Angiogenic and antiangiogenic factors in corneal avascularity, vasculogenesis, and wound healing (an American Ophthalmological Society thesis). Trans. Am. Ophthalmol. Soc. 2006, 104, 264–302. [Google Scholar]
- Tao, Y.; Bazan, H.E.; Bazan, N.G. Platelet-activating factor induces the expression of metalloproteinases-1 and -9, but not -2 or -3, in the corneal epithelium. Investig. Ophthalmol. Vis. Sci. 1995, 36, 345–354. [Google Scholar]
- Sang, Q.X.A. Complex role of matrix metalloproteinases in angiogenesis. Cell Res. 1998, 8, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Saghizadeh, M.; Brown, D.J.; Castellon, R.; Chwa, M.; Huang, G.H.; Ljubimova, J.Y.; Rosenberg, S.; Spirin, K.S.; Stolitenko, R.B.; Adachi, W. Overexpression of matrix metalloproteinase-10 and matrix metalloproteinase-3 in human diabetic corneas: A possible mechanism of basement membrane and integrin alterations. Am. J. Pathol. 2001, 158, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Zhou, D.; Fan, M.; Wang, H.; Huang, C.; Zhang, Z.; Wu, Y.; Li, W.; Chen, Y.; Liu, Z. Keratocytes create stromal spaces to promote corneal neovascularization via MMP13 expression. Investig. Ophthalmol. Vis. Sci. 2014, 55, 6691–6703. [Google Scholar] [CrossRef]
- Mohan, V.; Talmi-Frank, D.; Arkadash, V.; Papo, N.; Sagi, I. Matrix metalloproteinase protein inhibitors: Highlighting a new beginning for metalloproteinases in medicine. Met. Med 2016, 3, 31–47. [Google Scholar]
- Chang, J.-H.; Han, K.Y.; Azar, D.T. Wound healing fibroblasts modulate corneal angiogenic privilege: Interplay of basic fibroblast growth factor and matrix metalloproteinases in corneal angiogenesis. Jpn. J. Ophthalmol. 2010, 54, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.-Z.; Pang, X.; Peng, X.-J. Changes of matrix metalloproteinases in the stroma after corneal cross-linking in rabbits. Int. J. Ophthalmol. 2021, 14, 26. [Google Scholar] [CrossRef] [PubMed]
- Ma, D.H.-K.; Chen, J.-K.; Kim, W.-S.; Hao, Y.-X.; Wu, H.-C.; Tsai, R.J.-F.; Hwang, D.G.; Zhang, F. Expression of matrix metalloproteinases 2 and 9 and tissue inhibitors of metalloproteinase 1 and 2 in inflammation-induced corneal neovascularization. Ophthalmic Res. 2001, 33, 353–362. [Google Scholar] [CrossRef]
- Ebrahem, Q.; Qi, J.H.; Sugimoto, M.; Ali, M.; Sears, J.E.; Cutler, A.; Khokha, R.; Vasanji, A.; Anand-Apte, B. Increased neovascularization in mice lacking tissue inhibitor of metalloproteinases-3. Investig. Ophthalmol. Vis. Sci. 2011, 52, 6117–6123. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Chen, A.; Hong, A.; Xiong, S.; Chen, X.; Xie, Q. Shark Cartilage-Derived Anti-Angiogenic Peptide Inhibits Corneal Neovascularization. Bioengineering 2024, 11, 693. https://doi.org/10.3390/bioengineering11070693
Li Y, Chen A, Hong A, Xiong S, Chen X, Xie Q. Shark Cartilage-Derived Anti-Angiogenic Peptide Inhibits Corneal Neovascularization. Bioengineering. 2024; 11(7):693. https://doi.org/10.3390/bioengineering11070693
Chicago/Turabian StyleLi, Yunxian, Aoke Chen, An Hong, Sheng Xiong, Xiaojia Chen, and Qiuling Xie. 2024. "Shark Cartilage-Derived Anti-Angiogenic Peptide Inhibits Corneal Neovascularization" Bioengineering 11, no. 7: 693. https://doi.org/10.3390/bioengineering11070693
APA StyleLi, Y., Chen, A., Hong, A., Xiong, S., Chen, X., & Xie, Q. (2024). Shark Cartilage-Derived Anti-Angiogenic Peptide Inhibits Corneal Neovascularization. Bioengineering, 11(7), 693. https://doi.org/10.3390/bioengineering11070693