Optimizing the Amino Acid Sequence Enhances the Productivity and Bioefficacy of the RBP-Albumin Fusion Protein



Abstract

1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Isolation of Mouse Hepatic Stellate Cells (HSCs)
2.3. Transfection
2.4. SDS-PAGE and Western Blot Analysis
2.5. Quantitative Real-Time PCR
2.6. Statistical Analysis
3. Results
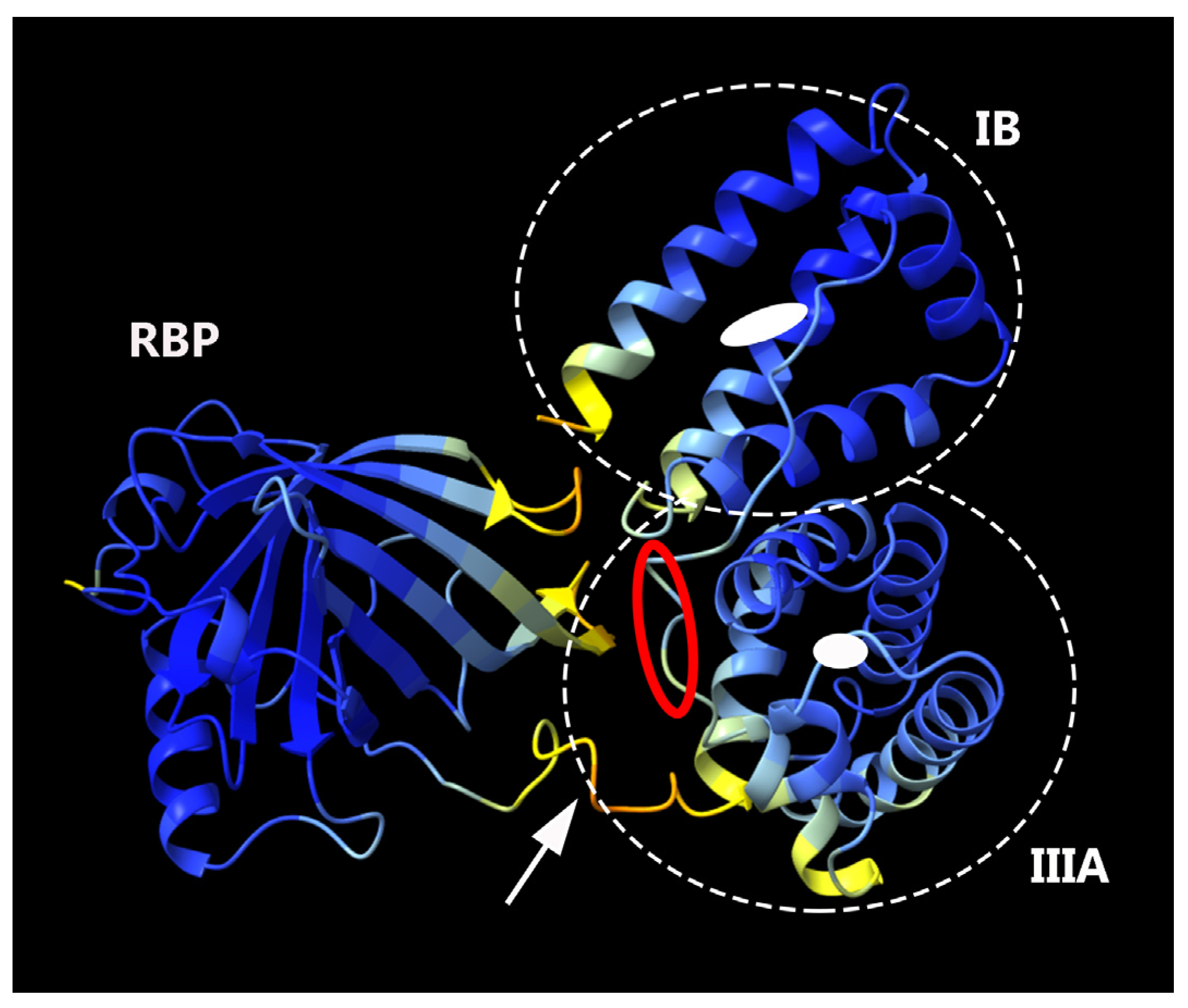
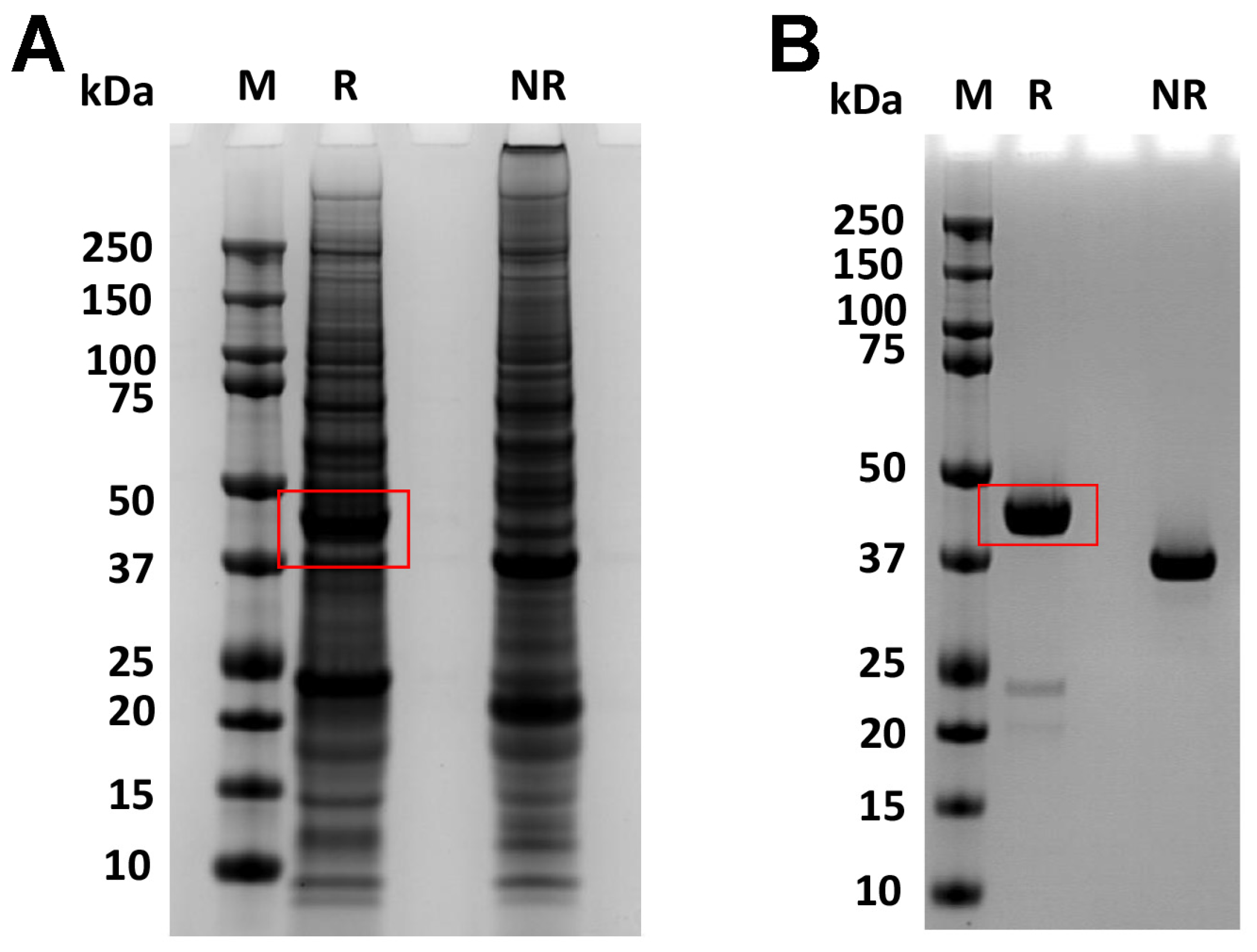
3.1. The Productivity of the Fusion Protein, RBP-IIIA-IB, Was Low
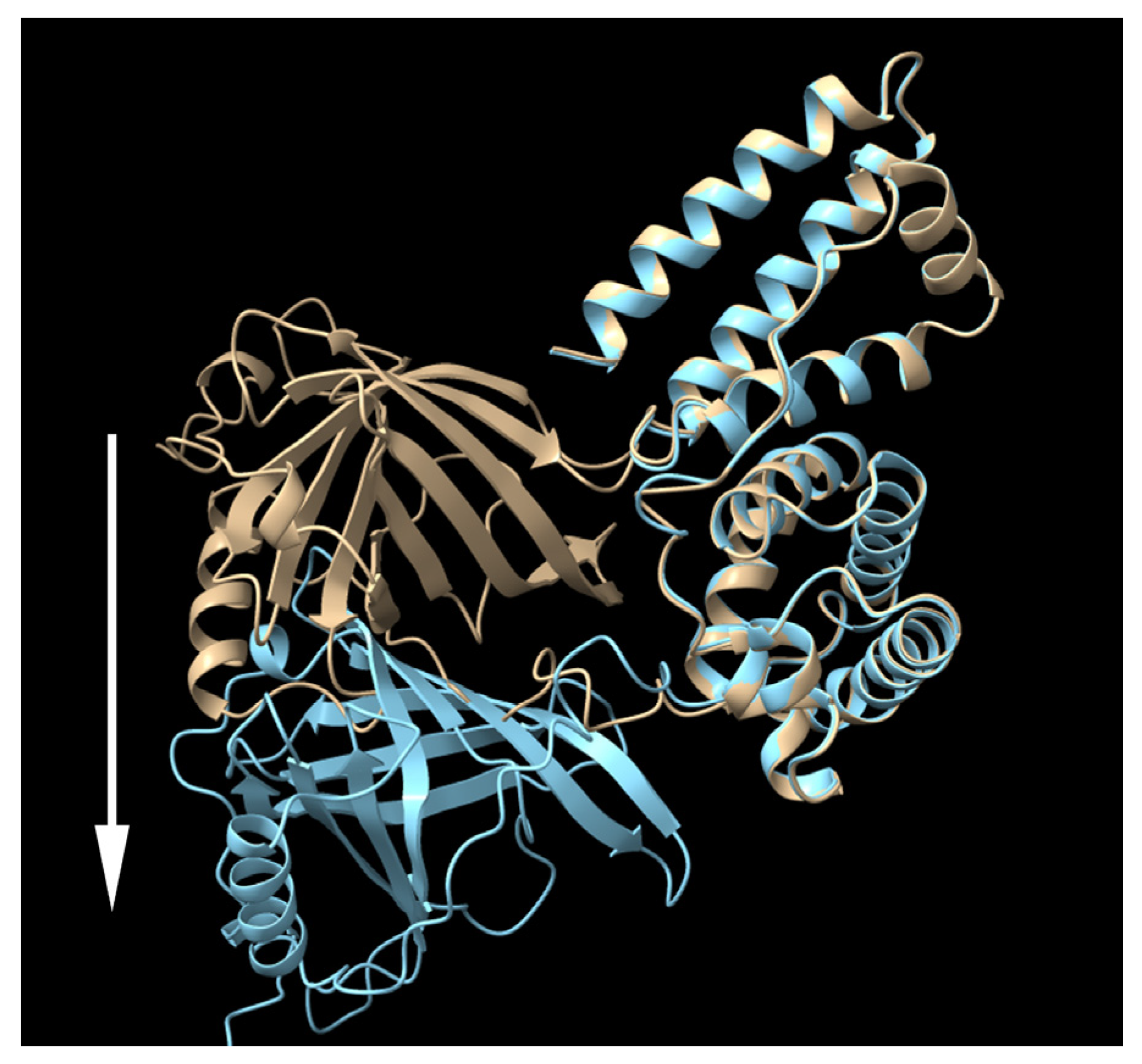
3.2. Altering the Linking Sequence between IIIA and IB Enhanced the Productivity and Bioactivity of RBP-IIIA-IB
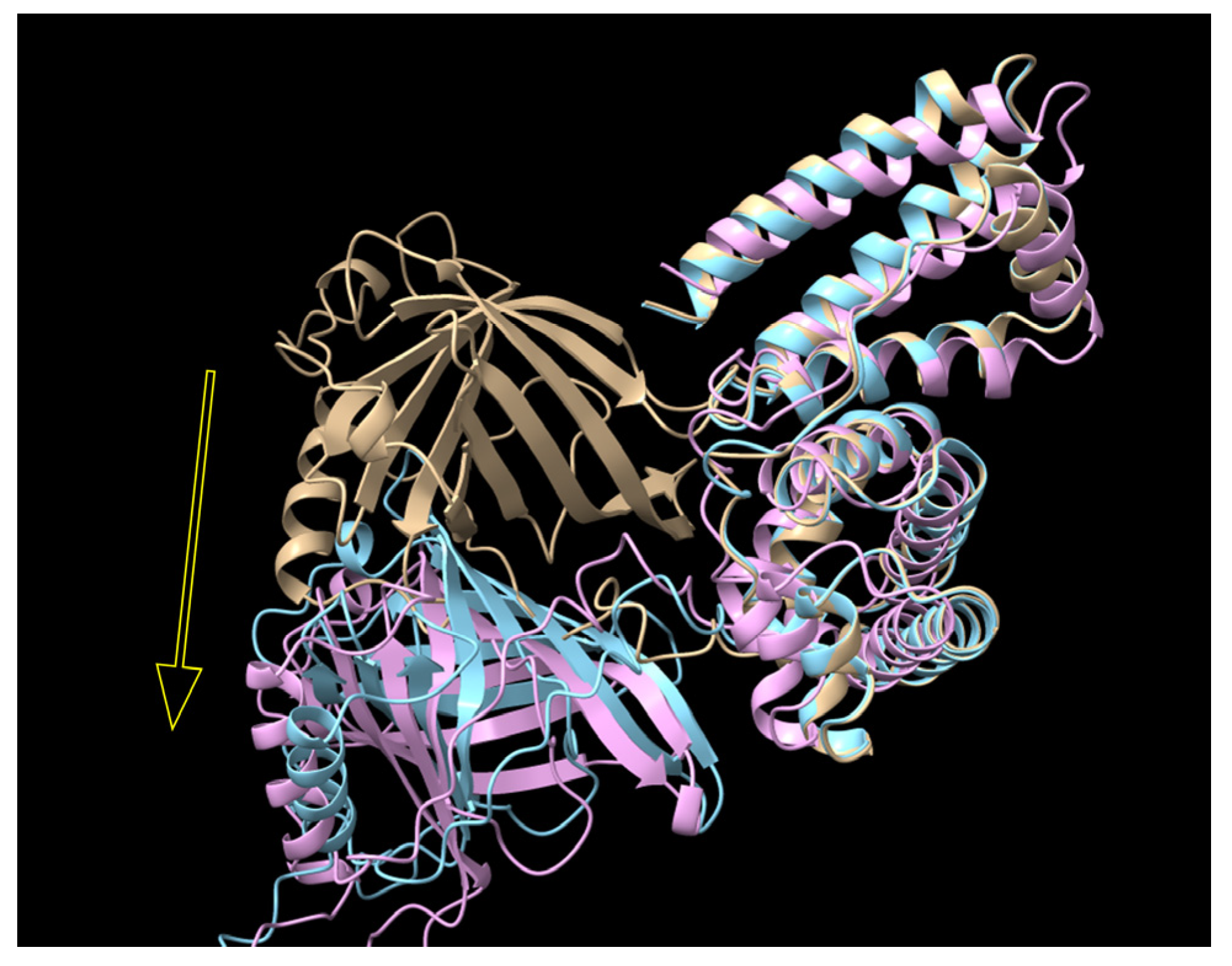
3.3. The Productivity of RBP-IIIA-AAAA-IB Was Further Enhanced by the Insertion of an Additional Disulfide Bond
3.4. The Fusion Protein RBP-IIIA-AAAA-IB_C453-480 Showed Improved Anti-Fibrotic Activity
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weiskirchen, R.; Weiskirchen, S.; Tacke, F. Organ and tissue fibrosis: Molecular signals, cellular mechanisms and translational implications. Mol. Asp. Med. 2019, 65, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Khomich, O.; Ivanov, A.V.; Bartosch, B. Metabolic Hallmarks of Hepatic Stellate Cells in Liver Fibrosis. Cells 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed]
- Kamm, D.R.; McCommis, K.S. Hepatic stellate cells in physiology and pathology. J. Physiol. 2022, 600, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Tsuchida, T.; Friedman, S.L. Mechanisms of hepatic stellate cell activation. Nat. Rev. Gastroenterol. Hepatol. 2017, 14, 397–411. [Google Scholar] [CrossRef] [PubMed]
- Roehlen, N.; Crouchet, E.; Baumert, T.F. Liver Fibrosis: Mechanistic Concepts and Therapeutic Perspectives. Cells 2020, 9, 875. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.; Yoo, W.; Lee, J.; Kim, H.; Lee, H.; Kim, Y.S.; Kim, D.U.; Oh, J. Formation of vitamin A lipid droplets in pancreatic stellate cells requires albumin. Gut 2009, 58, 1382–1390. [Google Scholar] [CrossRef] [PubMed]
- Curry, S.; Mandelkow, H.; Brick, P.; Franks, N. Crystal structure of human serum albumin complexed with fatty acid reveals an asymmetric distribution of binding sites. Nat. Struct. Biol. 1998, 5, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Park, S.; Kim, S.; Lim, C.; Kim, J.; Cha, D.R.; Oh, J. Recombinant fusion protein of albumin-retinol binding protein inactivates stellate cells. Biochem. Biophys. Res. Commun. 2012, 418, 191–197. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, R.; Yu, J.; Honda, J.; Hu, J.; Whitelegge, J.; Ping, P.; Wiita, P.; Bok, D.; Sun, H. A membrane receptor for retinol binding protein mediates cellular uptake of vitamin A. Science 2007, 315, 820–825. [Google Scholar] [CrossRef]
- Park, J.H.; Kim, J.; Choi, S.Y.; Lee, B.; Lee, J.E.; Park, H.; Moon, J.W.; Park, S.H.; Lee, J.M.; Lee, H.S.; et al. Albumin inhibits the nuclear translocation of Smad3 via interleukin-1beta signaling in hepatic stellate cells. Sci. Rep. 2021, 11, 3196. [Google Scholar] [CrossRef]
- Belatik, A.; Hotchandani, S.; Bariyanga, J.; Tajmir-Riahi, H.A. Binding sites of retinol and retinoic acid with serum albumins. Eur. J. Med. Chem. 2012, 48, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Jeong, H.; Park, S.; Yoo, W.; Choi, S.; Choi, K.; Lee, M.G.; Lee, M.; Cha, D.; Kim, Y.S.; et al. Fusion protein of retinol-binding protein and albumin domain III reduces liver fibrosis. EMBO Mol. Med. 2015, 7, 819–830. [Google Scholar] [CrossRef] [PubMed]
- Cha, J.J.; Mandal, C.; Ghee, J.Y.; Yoo, J.A.; Lee, M.J.; Kang, Y.S.; Hyun, Y.Y.; Lee, J.E.; Kim, H.W.; Han, S.Y.; et al. Inhibition of Renal Stellate Cell Activation Reduces Renal Fibrosis. Biomedicines 2020, 8, 431. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Son, Y.; Moon, J.W.; Park, D.W.; Kim, Y.S.; Oh, J. Fusion Protein of RBP and Albumin Domain III Reduces Lung Fibrosis by Inactivating Lung Stellate Cells. Biomedicines 2023, 11, 2007. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zaro, J.L.; Shen, W.C. Fusion protein linkers: Property, design and functionality. Adv. Drug Deliv. Rev. 2013, 65, 1357–1369. [Google Scholar] [CrossRef] [PubMed]
- Arai, R.; Ueda, H.; Kitayama, A.; Kamiya, N.; Nagamune, T. Design of the linkers which effectively separate domains of a bifunctional fusion protein. Protein Eng. 2001, 14, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Mederacke, I.; Dapito, D.H.; Affo, S.; Uchinami, H.; Schwabe, R.F. High-yield and high-purity isolation of hepatic stellate cells from normal and fibrotic mouse livers. Nat. Protoc. 2015, 10, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Friedman, S.L. Hepatic stellate cells: Protean, multifunctional, and enigmatic cells of the liver. Physiol. Rev. 2008, 88, 125–172. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Yang, C.; Gao, X.; Li, X.; Zhang, Z.; Gong, R. Comprehensive elucidation of the structural and functional roles of engineered disulfide bonds in antibody Fc fragment. J. Biol. Chem. 2018, 293, 19127–19135. [Google Scholar] [CrossRef]
- Murphy, R.M.; Roberts, C.J. Protein misfolding and aggregation research: Some thoughts on improving quality and utility. Biotechnol. Prog. 2013, 29, 1109–1115. [Google Scholar] [CrossRef]
- Patel, D.K.; Menon, D.V.; Patel, D.H.; Dave, G. Linkers: A synergistic way for the synthesis of chimeric proteins. Protein Expr. Purif. 2022, 191, 106012. [Google Scholar] [CrossRef]
- Zhao, H.L.; Yao, X.Q.; Xue, C.; Wang, Y.; Xiong, X.H.; Liu, Z.M. Increasing the homogeneity, stability and activity of human serum albumin and interferon-alpha2b fusion protein by linker engineering. Protein Expr. Purif. 2008, 61, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Shen, W.C. Improving the oral efficacy of recombinant granulocyte colony-stimulating factor and transferrin fusion protein by spacer optimization. Pharm. Res. 2006, 23, 2116–2121. [Google Scholar] [CrossRef] [PubMed]
- Amet, N.; Lee, H.F.; Shen, W.C. Insertion of the designed helical linker led to increased expression of tf-based fusion proteins. Pharm. Res. 2009, 26, 523–528. [Google Scholar] [CrossRef] [PubMed]
- Argos, P. An investigation of oligopeptides linking domains in protein tertiary structures and possible candidates for general gene fusion. J. Mol. Biol. 1990, 211, 943–958. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Ueda, H.; Kazami, J.; Kawano, G.; Suzuki, E.; Nagamune, T. Engineering of functional chimeric protein G-Vargula luciferase. Anal. Biochem. 1997, 249, 147–152. [Google Scholar] [CrossRef] [PubMed]
- George, R.A.; Heringa, J. An analysis of protein domain linkers: Their classification and role in protein folding. Protein Eng. 2002, 15, 871–879. [Google Scholar] [CrossRef]
- Kassa, A.; Dey, A.K.; Sarkar, P.; Labranche, C.; Go, E.P.; Clark, D.F.; Sun, Y.; Nandi, A.; Hartog, K.; Desaire, H.; et al. Stabilizing exposure of conserved epitopes by structure guided insertion of disulfide bond in HIV-1 envelope glycoprotein. PLoS ONE 2013, 8, e76139. [Google Scholar] [CrossRef]
- Ning, J.; Ahmed, S.; Cheng, G.; Chen, T.; Wang, Y.; Peng, D.; Yuan, Z. Analysis of the stability and affinity of BlaR-CTD protein to β-lactam antibiotics based on docking and mutagenesis studies. J. Biol. Eng. 2019, 13, 27. [Google Scholar] [CrossRef]
- Xu, T.; Xu, Q.; Li, J. Toward the appropriate interpretation of Alphafold2. Front. Artif. Intell. 2023, 6, 1149748. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Amino Acid Sequence |
|---|---|
| RBP (1-193) | mkwvwallllaawaaaERDCRVSSFRVKENFDKARFSGTWY AMAKKDPEGLFLQDNIVAEFSVDETGQMSATAKGRVR LLNNWDVCADMVGTFTDTEDPAKFKMKYWGVASFLQ KGNDDHWIVDTDYDTYAVQYSCRLLNLDGTCADSYSF VFSRDPNGLPPEAQKIVRQRQEELCLARQYRLIVHNGY CDGR |
| albumin IIIA (404-517) | L VEEPQNLIKQNCELFEQLGE YKFQNALLVRYTKKVPQVST PTLVEVSRNLGKVGSKCCKH PEAKRMPCAEDYLSVVLNQL CVLHEKTPVSDRVTKCCTES LVNRRPCFSALEV |
| albumin IB (131-218) | DDNPNLPRLVRPEV DVMCTAFHDNEETFLKKYLY EIARRHPYFYAPELLFFAKR YKAAFTECCQAADKAACLLP KLDELRDEGKASSA hhhhhhhh |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.H.; Kwon, S.; Choi, S.-Y.; Kim, B.; Oh, J. Optimizing the Amino Acid Sequence Enhances the Productivity and Bioefficacy of the RBP-Albumin Fusion Protein. Bioengineering 2024, 11, 617. https://doi.org/10.3390/bioengineering11060617
Park JH, Kwon S, Choi S-Y, Kim B, Oh J. Optimizing the Amino Acid Sequence Enhances the Productivity and Bioefficacy of the RBP-Albumin Fusion Protein. Bioengineering. 2024; 11(6):617. https://doi.org/10.3390/bioengineering11060617
Chicago/Turabian StylePark, Ji Hoon, Sohyun Kwon, So-Young Choi, Bongcheol Kim, and Junseo Oh. 2024. "Optimizing the Amino Acid Sequence Enhances the Productivity and Bioefficacy of the RBP-Albumin Fusion Protein" Bioengineering 11, no. 6: 617. https://doi.org/10.3390/bioengineering11060617
APA StylePark, J. H., Kwon, S., Choi, S.-Y., Kim, B., & Oh, J. (2024). Optimizing the Amino Acid Sequence Enhances the Productivity and Bioefficacy of the RBP-Albumin Fusion Protein. Bioengineering, 11(6), 617. https://doi.org/10.3390/bioengineering11060617