
Probiotic Enterococcus Faecium Attenuated Atherosclerosis by Improving SCFAs Associated with Gut Microbiota in ApoE−/− Mice


Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria
2.2. Animals
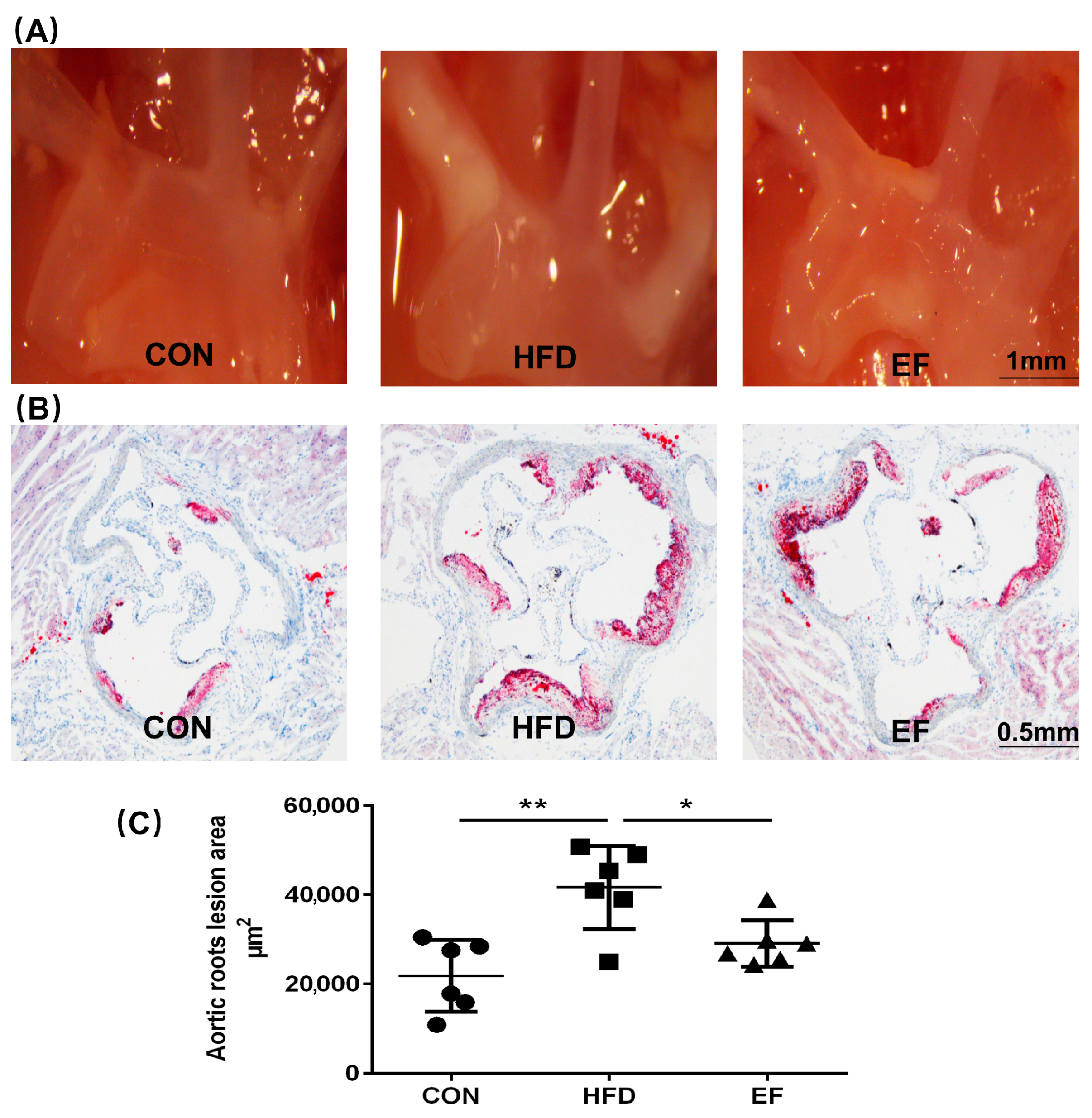
2.3. Atherosclerotic Lesions Analysis
2.4. Serum Inflammatory Factor Assays
2.5. Performing RNA Extraction and qRT-PCR Analysis
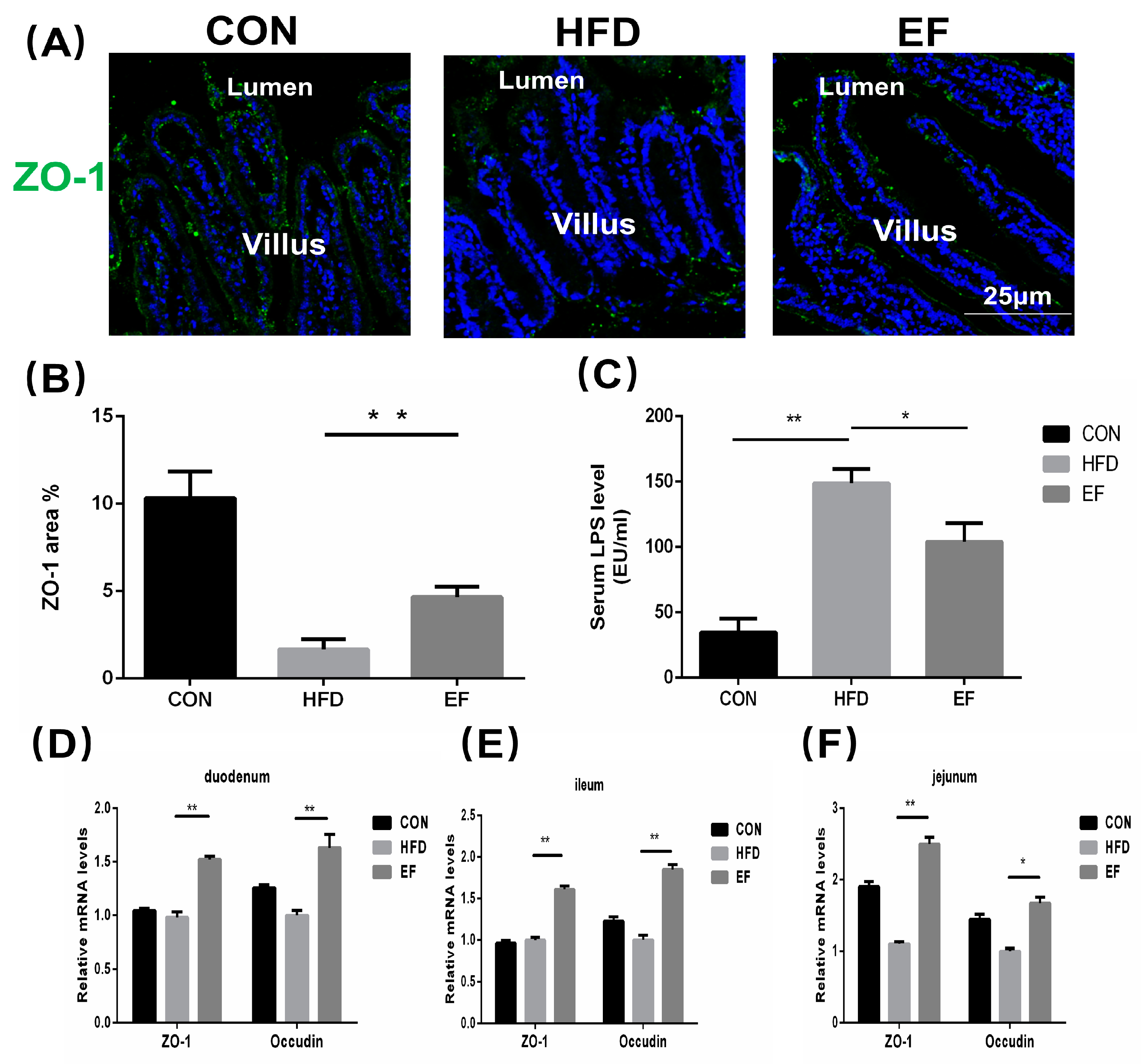
2.6. Histology and Immunofluorescent Staining
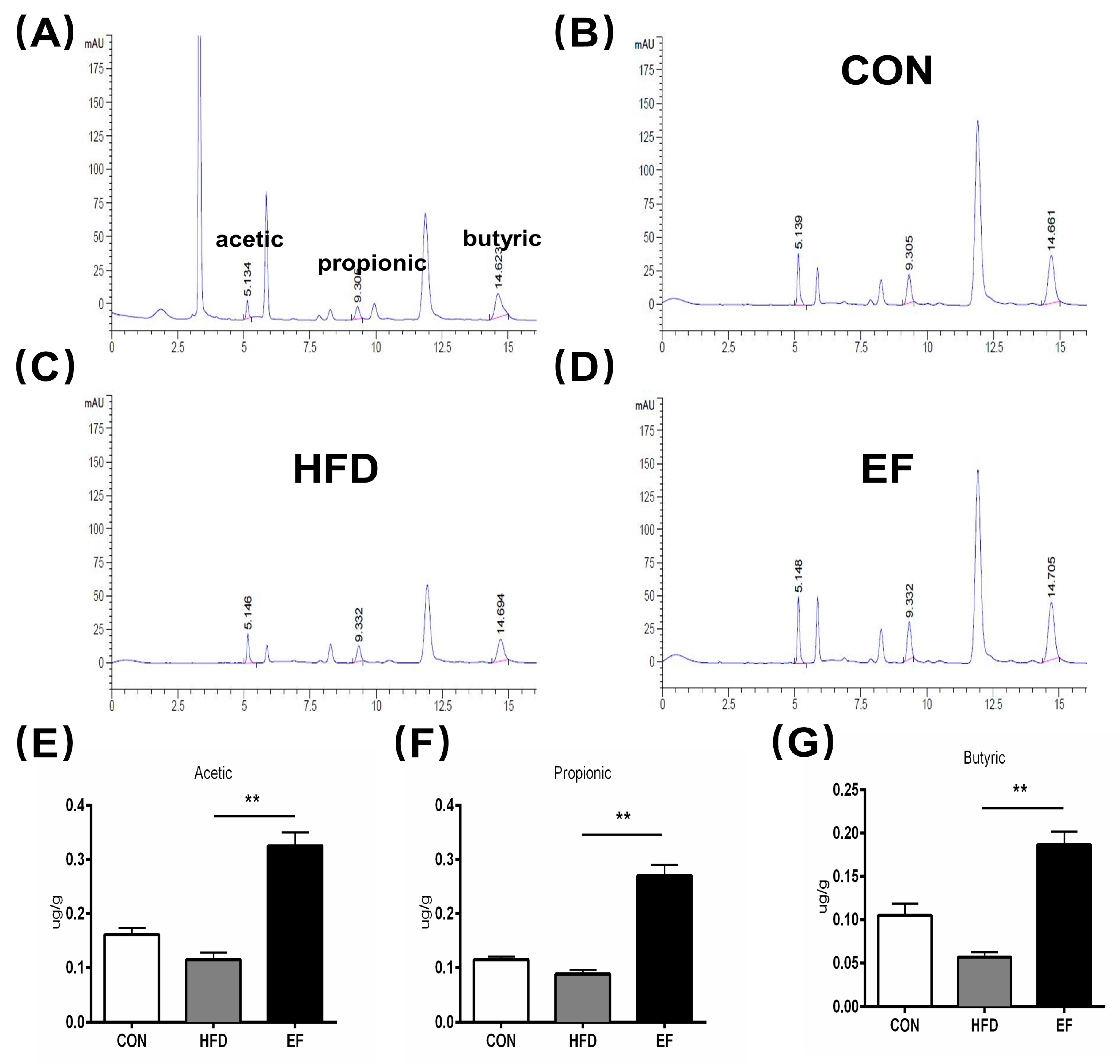
2.7. SCFAs Analysis
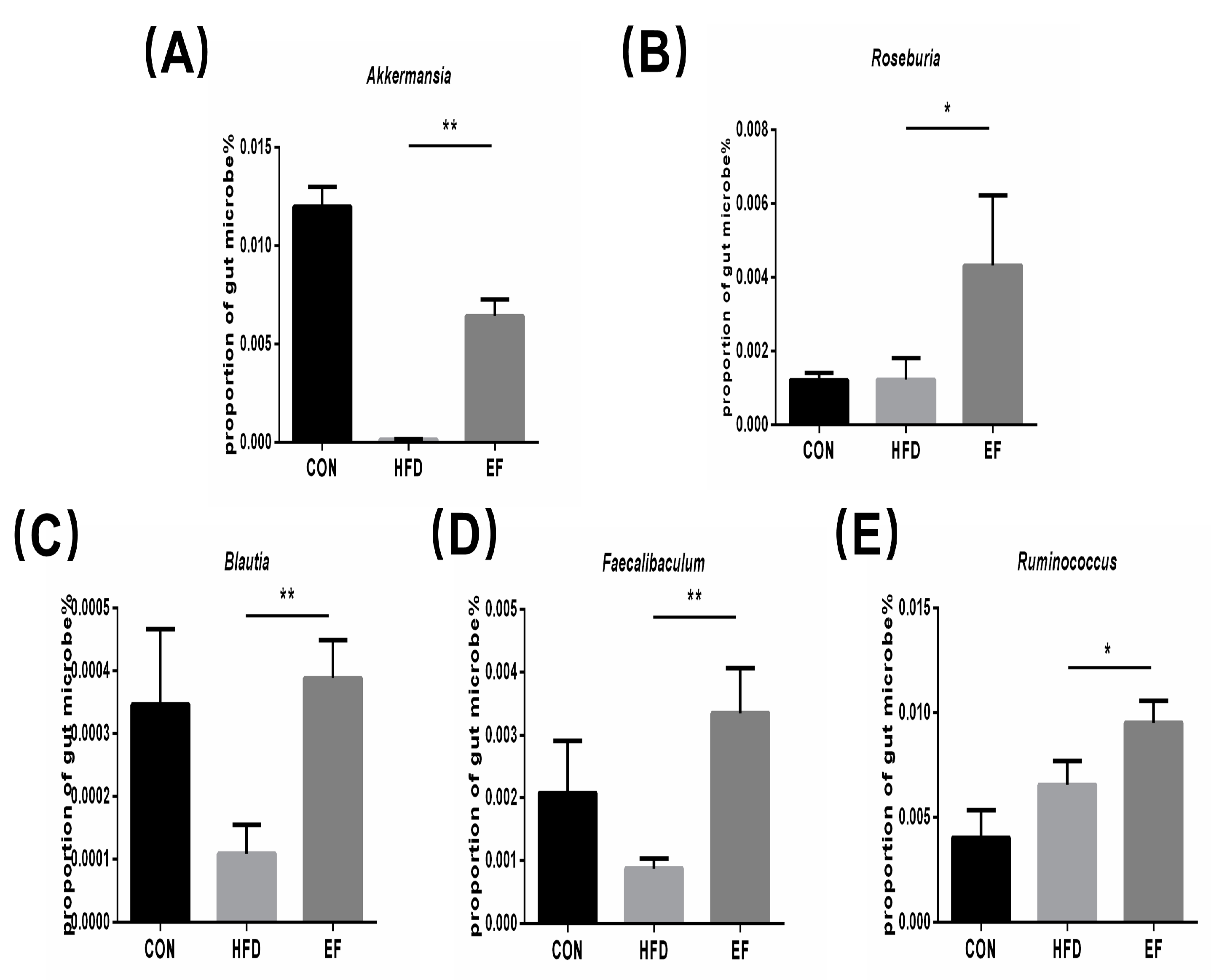
2.8. Gut Microbiota Analysis
2.9. Statistical Analysis
3. Results
3.1. Atherosclerotic Lesion Development Was Diminished by E. Faecium NCIMB11508
3.2. Enterococcus Faecium NCIMB11508 Alleviated the Inflammation of the Aorta
3.3. Enterococcus Faecium NCIMB11508 Decreased Intestinal Permeability for Lipopolysaccharides
3.4. The Quantity of SCFAs Was Amplified by Enterococcus Faecium NCIMB11508
3.5. The Gut Microbiota’s Composition Was Altered by Enterococcus Faecium NCIMB11508
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soppert, J.; Lehrke, M.; Marx, N.; Jankowski, J.; Noels, H. Lipoproteins and Lipids in Cardiovascular Disease: From Mechanistic Insights to Therapeutic Targeting. Adv. Drug Deliv. Rev. 2020, 159, 4–33. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Q.; Tian, F.; Xu, J.; Du, X.; Zhang, S.; Liu, L. New Insight into Dyslipidemia-Induced Cellular Senescence Atherosclerosis. Biol. Rev. Camb. Philos. Soc. 2022, 97, 1844–1867. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Donners, M.M.P.C.; Karel, J.; de Boer, H.; van Zonneveld, A.J.; den Ruijter, H.; Jukema, J.W.; Kraaijeveld, A.; Kuiper, J.; Pasterkamp, G.; et al. Sex-Specific Differences in Cytokine Signaling Pathways in Circulating Monocytes of Cardiovascular Disease Patients. Atherosclerosis 2023, 384, 117123. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.; Shapiro, M.D. Apolipoproteins in Vascular Biology and Atherosclerotic Disease. Nat. Rev. Cardiol. 2022, 19, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Clyburn, C.; Birren, S.J. Crosstalk between Nerves, Immune Cells and Plaques Drives Atherosclerosis. Nature 2022, 605, 32–34. [Google Scholar] [CrossRef]
- Gholami, H.; Chmiel, J.A.; Burton, J.P.; Maleki Vareki, S. The Role of Microbiota-Derived Vitamins in Immune Homeostasis and Enhancing Cancer Immunotherapy. Cancers 2023, 15, 1300. [Google Scholar] [CrossRef]
- Abernathy-Close, L.; Barron, M.R.; George, J.M.; Dieterle, M.G.; Vendrov, K.C.; Bergin, I.L.; Young, V.B. Intestinal Inflammation and Altered Gut Microbiota Associated with Inflammatory Bowel Disease Render Mice Susceptible to Clostridioides Difficile Colonization and Infection. mBio 2021, 12, e0273320. [Google Scholar] [CrossRef]
- TET1s Deficiency Exacerbates Oscillatory Shear Flow-Induced Atherosclerosis. Available online: https://pubmed.ncbi.nlm.nih.gov/35342333/ (accessed on 2 July 2024).
- Piewngam, P.; Khongthong, S.; Roekngam, N.; Theapparat, Y.; Sunpaweravong, S.; Faroongsarng, D.; Otto, M. Probiotic for Pathogen-Specific Staphylococcus Aureus Decolonisation in Thailand: A Phase 2, Double-Blind, Randomised, Placebo-Controlled Trial. Lancet Microbe 2023, 4, e75–e83. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, T.; Din, A.U.; Hassan, A.; Wang, Y.; Wang, G. Beneficial Effects of Enterococcus Faecalis in Hypercholesterolemic Mice on Cholesterol Transportation and Gut Microbiota. Appl. Microbiol. Biotechnol. 2019, 103, 3181–3191. [Google Scholar] [CrossRef]
- Hassan, A.; Din, A.U.; Zhu, Y.; Zhang, K.; Li, T.; Wang, Y.; Xu, S.; Lei, H.; Yu, X.; Wang, G. Anti-Atherosclerotic Effects of Lactobacillus Plantarum ATCC 14917 in ApoE−/− Mice through Modulation of Proinflammatory Cytokines and Oxidative Stress. Appl. Microbiol. Biotechnol. 2020, 104, 6337–6350. [Google Scholar] [CrossRef]
- Kiousi, D.E.; Karapetsas, A.; Karolidou, K.; Panayiotidis, M.I.; Pappa, A.; Galanis, A. Probiotics in Extraintestinal Diseases: Current Trends and New Directions. Nutrients 2019, 11, 788. [Google Scholar] [CrossRef] [PubMed]
- Guerra, N.P.; Fajardo, P.; Fuciños, C.; Amado, I.R.; Alonso, E.; Torrado, A.; Pastrana, L. Modelling the Biphasic Growth and Product Formation by Enterococcus Faecium CECT 410 in Realkalized Fed-Batch Fermentations in Whey. J. Biomed. Biotechnol. 2010, 2010, 290286. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, R.; Khanal, S.; Mathias, A.; Gupta, S.; Lallo, J.; Sahu, S.; Ohanyan, V.; Patel, A.; Storm, K.; Datta, S.; et al. TSP-1 (Thrombospondin-1) Deficiency Protects ApoE-/- Mice Against Leptin-Induced Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2021, 41, e112–e127. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Lan, B.; Zheng, T.; Yang, L.; Zhang, X.; Cheng, L.; Tuerhongjiang, G.; Yuan, Z.; Wu, Y. GSDME-Mediated Pyroptosis Promotes the Progression and Associated Inflammation of Atherosclerosis. Nat. Commun. 2023, 14, 929. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-W.; Ivanov, S.; Williams, J.W. Monocyte Recruitment, Specification, and Function in Atherosclerosis. Cells 2020, 10, 15. [Google Scholar] [CrossRef]
- Wang, W.; Liu, W.; Fidler, T.; Wang, Y.; Tang, Y.; Woods, B.; Welch, C.; Cai, B.; Silvestre-Roig, C.; Ai, D.; et al. Macrophage Inflammation, Erythrophagocytosis, and Accelerated Atherosclerosis in Jak2 V617F Mice. Circ. Res. 2018, 123, e35–e47. [Google Scholar] [CrossRef]
- Dong, Y.; Wang, B.; Cui, K.; Cai, X.; Bhattacharjee, S.; Wong, S.; Cowan, D.B.; Chen, H. Epsins Negatively Regulate Aortic Endothelial Cell Function by Augmenting Inflammatory Signaling. Cells 2021, 10, 1918. [Google Scholar] [CrossRef]
- Verhaar, B.J.H.; Prodan, A.; Nieuwdorp, M.; Muller, M. Gut Microbiota in Hypertension and Atherosclerosis: A Review. Nutrients 2020, 12, 2982. [Google Scholar] [CrossRef]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-Chain Fatty Acid Propionate Protects From Hypertensive Cardiovascular Damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef]
- Fang, F.; Ni, Y.; Yu, H.; Yin, H.; Yang, F.; Li, C.; Sun, D.; Pei, T.; Ma, J.; Deng, L.; et al. Inflammatory Endothelium-Targeted and Cathepsin Responsive Nanoparticles Are Effective against Atherosclerosis. Theranostics 2022, 12, 4200–4220. [Google Scholar] [CrossRef]
- Chen, L.; Liu, W.; Li, Y.; Luo, S.; Liu, Q.; Zhong, Y.; Jian, Z.; Bao, M. Lactobacillus Acidophilus ATCC 4356 Attenuates the Atherosclerotic Progression through Modulation of Oxidative Stress and Inflammatory Process. Int. Immunopharmacol. 2013, 17, 108–115. [Google Scholar] [CrossRef]
- Mencarelli, A.; Cipriani, S.; Renga, B.; Bruno, A.; D’Amore, C.; Distrutti, E.; Fiorucci, S. VSL#3 Resets Insulin Signaling and Protects against NASH and Atherosclerosis in a Model of Genetic Dyslipidemia and Intestinal Inflammation. PLoS ONE 2012, 7, e45425. [Google Scholar] [CrossRef]
- Libby, P. Inflammation during the Life Cycle of the Atherosclerotic Plaque. Cardiovasc. Res. 2021, 117, 2525–2536. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.; Zhang, X.; Ma, Z.; Niu, J.; Ma, S.; Wenjie, W.; Chen, J. Combination of Tanshinone IIA and Astragaloside IV Attenuate Atherosclerotic Plaque Vulnerability in ApoE(-/-) Mice by Activating PI3K/AKT Signaling and Suppressing TRL4/NF-κB Signaling. Biomed. Pharmacother. 2020, 123, 109729. [Google Scholar] [CrossRef] [PubMed]
- Anez-Bustillos, L.; Dao, D.T.; Finkelstein, A.; Pan, A.; Cho, B.S.; Mitchell, P.D.; Gura, K.M.; Bistrian, B.R.; Puder, M. Metabolic and Inflammatory Effects of an ω-3 Fatty Acid-Based Eucaloric Ketogenic Diet in Mice With Endotoxemia. JPEN J. Parenter. Enteral Nutr. 2019, 43, 986–997. [Google Scholar] [CrossRef]
- He, C.; Deng, J.; Hu, X.; Zhou, S.; Wu, J.; Xiao, D.; Darko, K.O.; Huang, Y.; Tao, T.; Peng, M.; et al. Vitamin A Inhibits the Action of LPS on the Intestinal Epithelial Barrier Function and Tight Junction Proteins. Food Funct. 2019, 10, 1235–1242. [Google Scholar] [CrossRef]
- Duttaroy, A.K. Role of Gut Microbiota and Their Metabolites on Atherosclerosis, Hypertension and Human Blood Platelet Function: A Review. Nutrients 2021, 13, 144. [Google Scholar] [CrossRef]
- den Besten, G.; Bleeker, A.; Gerding, A.; van Eunen, K.; Havinga, R.; van Dijk, T.H.; Oosterveer, M.H.; Jonker, J.W.; Groen, A.K.; Reijngoud, D.-J.; et al. Short-Chain Fatty Acids Protect Against High-Fat Diet-Induced Obesity via a PPARγ-Dependent Switch From Lipogenesis to Fat Oxidation. Diabetes 2015, 64, 2398–2408. [Google Scholar] [CrossRef]
- Zhou, M.-S.; Zhang, B.; Gao, Z.-L.; Zheng, R.-P.; Marcellin, D.F.H.M.; Saro, A.; Pan, J.; Chu, L.; Wang, T.-S.; Huang, J.-F. Altered Diversity and Composition of Gut Microbiota in Patients with Allergic Rhinitis. Microb. Pathog. 2021, 161, 105272. [Google Scholar] [CrossRef]
- Yu, G.; Ji, X.; Huang, J.; Liao, A.; Pan, L.; Hou, Y.; Hui, M.; Guo, W. Immunity Improvement and Gut Microbiota Remodeling of Mice by Wheat Germ Globulin. World J. Microbiol. Biotechnol. 2021, 37, 64. [Google Scholar] [CrossRef]
- Li, J.; Lin, S.; Vanhoutte, P.M.; Woo, C.W.; Xu, A. Akkermansia Muciniphila Protects Against Atherosclerosis by Preventing Metabolic Endotoxemia-Induced Inflammation in Apoe−/− Mice. Circulation 2016, 133, 2434–2446. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence |
|---|---|
| F4/80 | F:5′-CTTTGGCTATGGGCTTCCAGTC-3′ |
| R:5′-GCAAGGAGGACAGAGTTTATCGTG-3′ | |
| MCP-1 | F:5′-CCACTCACCTGCTGCTACTCA-3′ |
| R:5′-TGGTGATCCTCTTGTAGCTCTCC-3′ | |
| TNF-α | F:5′-ACGGCATGGATCTCAAAGAC-3′ |
| R:5′-AGATAGCAAATCGGCTGACG-3′ | |
| ZO-1 | F:5′-TTTTTGACAGGGGGAGTGG-3′ |
| R:5′- TGCTGCAGAGGTCAAAGTTCAAG-3′ | |
| Occlude | F:5′-ATGTCCGGCCGATGCTCTC-3′ |
| R:5′-TTTGGCTGCTCTTGGGTCTGTAT-3′ | |
| β-actin | F:5′-GTGGGCCGGTCTAGGCACCAA-3′ |
| R:5′-CGGTTGCCTTAGGGTTCAGG-3′ | |
| 341F | F:5′-CCTACGGGAGGCAGCAG-3′ |
| 806R | F:5′-GGACTACACGGGTATCTAAT-3′ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, Y.; Yin, C.; Wang, Y. Probiotic Enterococcus Faecium Attenuated Atherosclerosis by Improving SCFAs Associated with Gut Microbiota in ApoE−/− Mice. Bioengineering 2024, 11, 1033. https://doi.org/10.3390/bioengineering11101033
Zhu Y, Yin C, Wang Y. Probiotic Enterococcus Faecium Attenuated Atherosclerosis by Improving SCFAs Associated with Gut Microbiota in ApoE−/− Mice. Bioengineering. 2024; 11(10):1033. https://doi.org/10.3390/bioengineering11101033
Chicago/Turabian StyleZhu, Yuan, Chao Yin, and Yeqi Wang. 2024. "Probiotic Enterococcus Faecium Attenuated Atherosclerosis by Improving SCFAs Associated with Gut Microbiota in ApoE−/− Mice" Bioengineering 11, no. 10: 1033. https://doi.org/10.3390/bioengineering11101033
APA StyleZhu, Y., Yin, C., & Wang, Y. (2024). Probiotic Enterococcus Faecium Attenuated Atherosclerosis by Improving SCFAs Associated with Gut Microbiota in ApoE−/− Mice. Bioengineering, 11(10), 1033. https://doi.org/10.3390/bioengineering11101033