The Membrane-Active Phytopeptide Cycloviolacin O2 Simultaneously Targets HIV-1-infected Cells and Infectious Viral Particles to Potentiate the Efficacy of Antiretroviral Drugs

 , , ,
, , , 
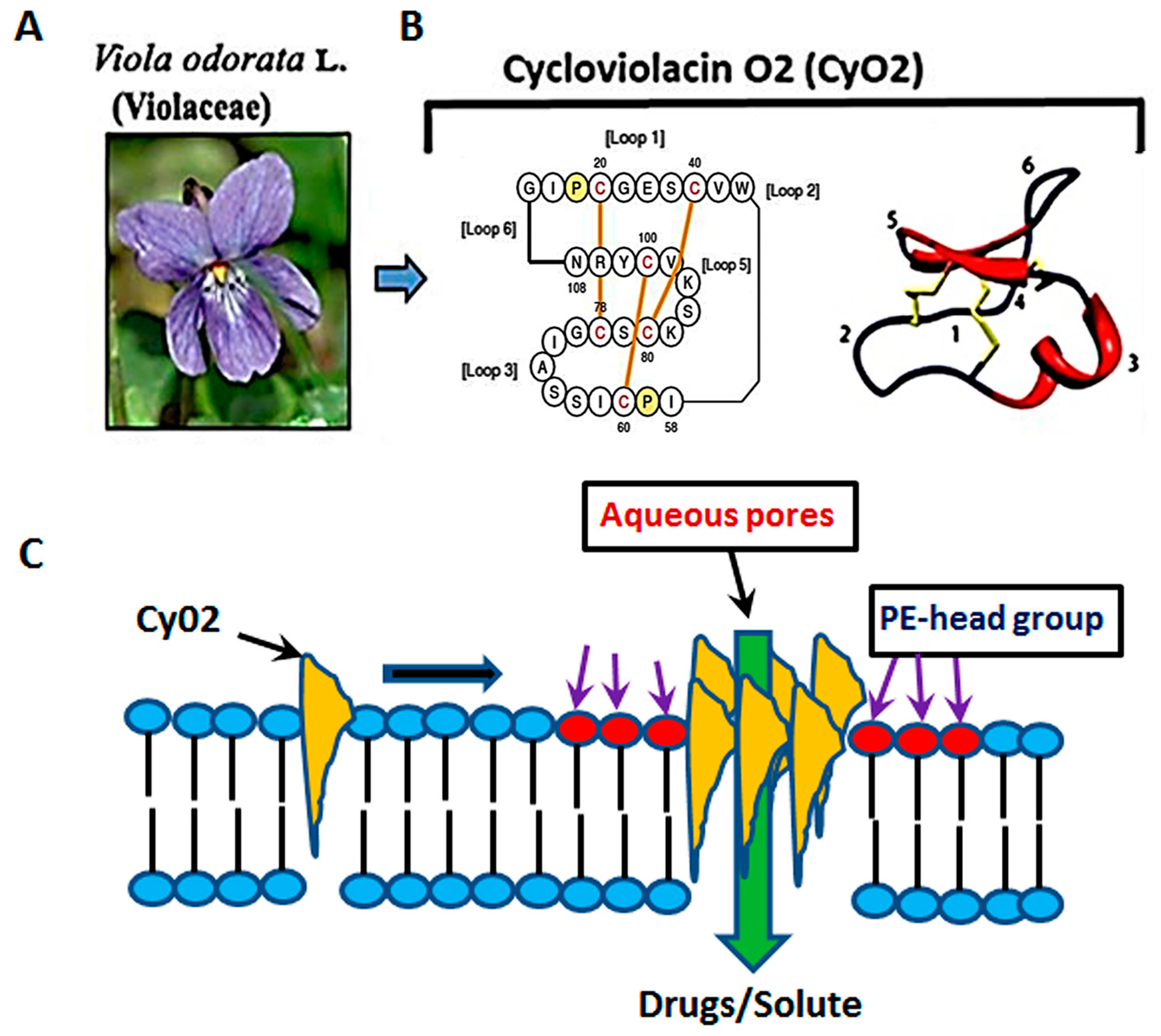{kind=link}
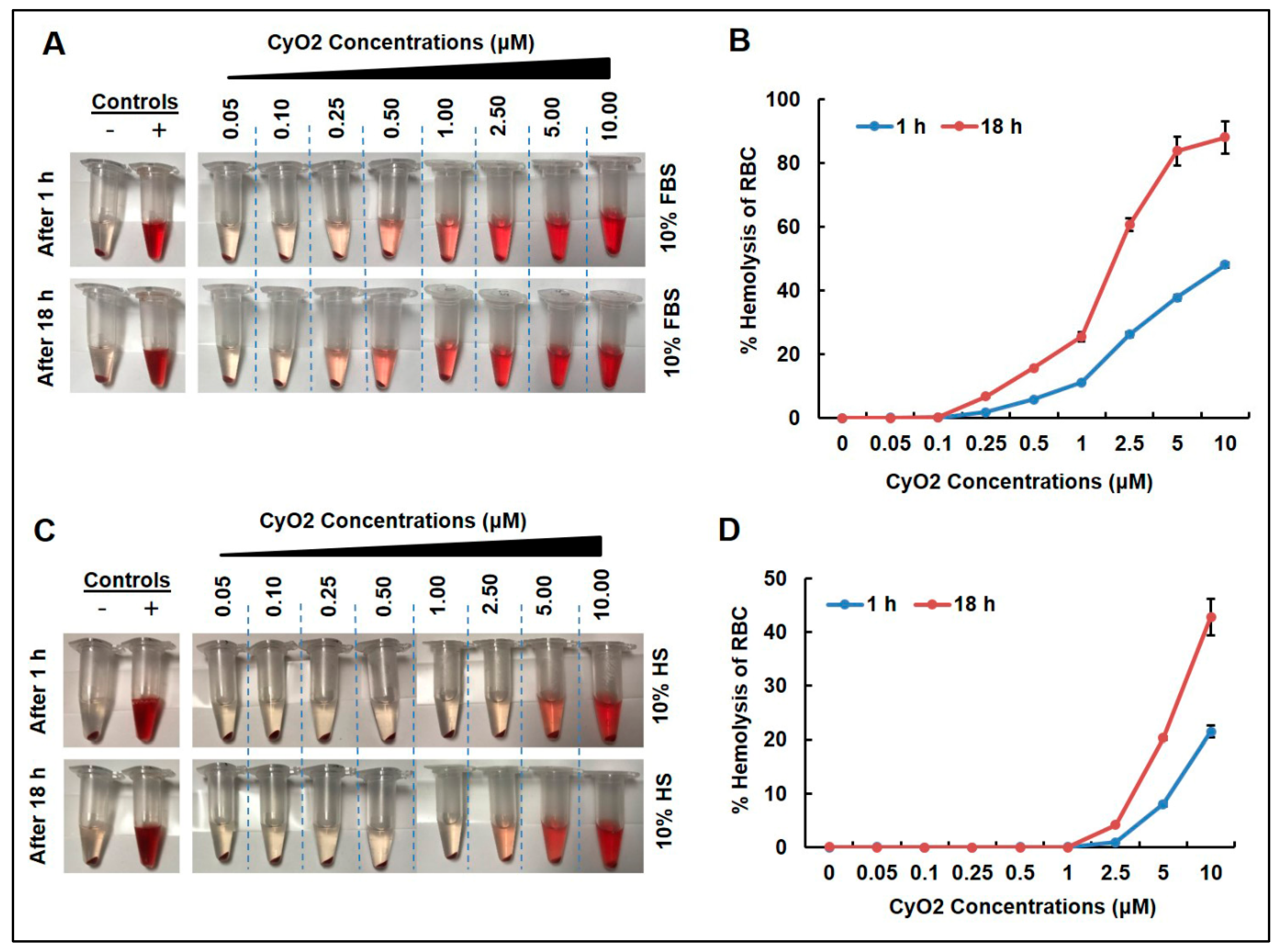{kind=link}
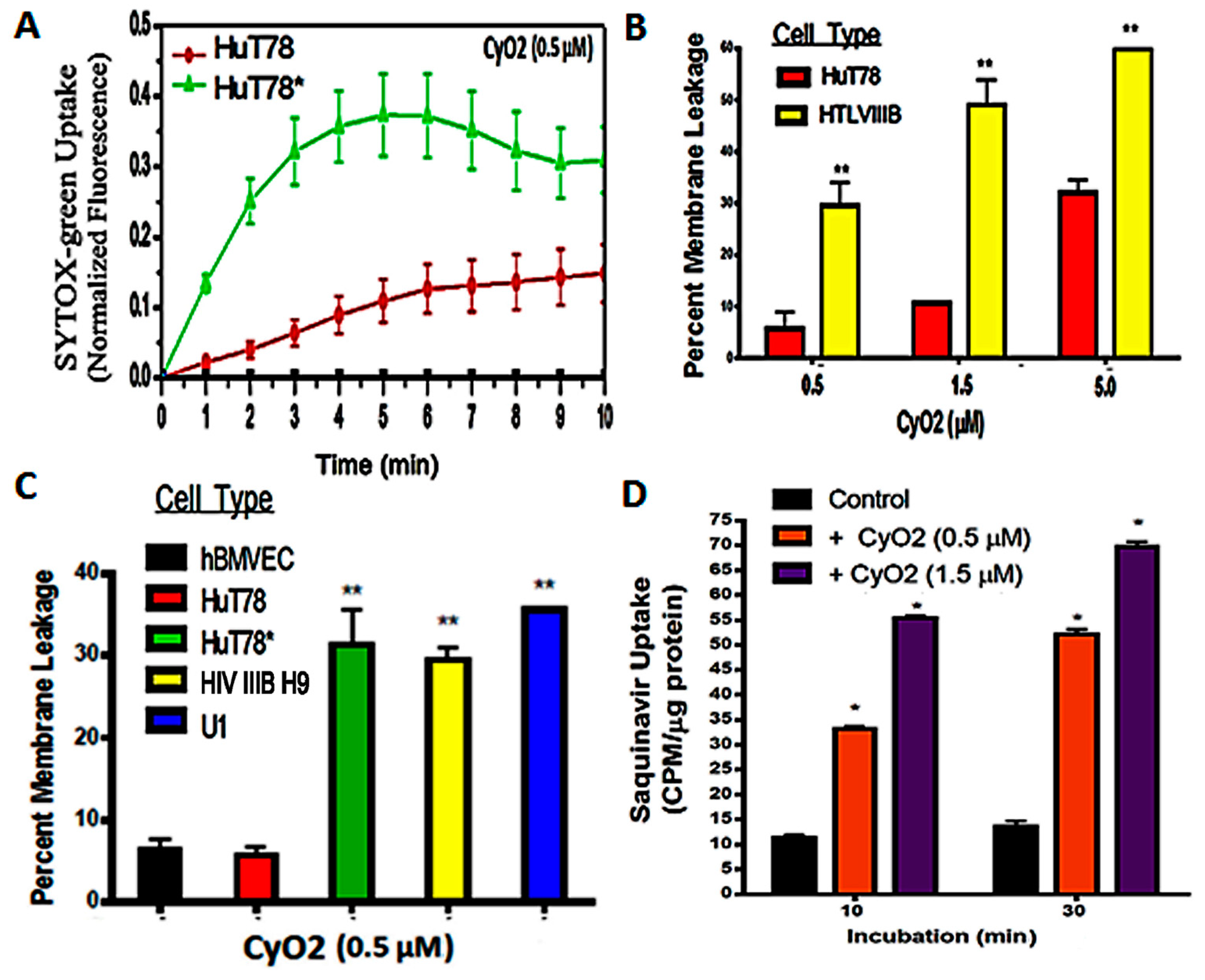{kind=link}
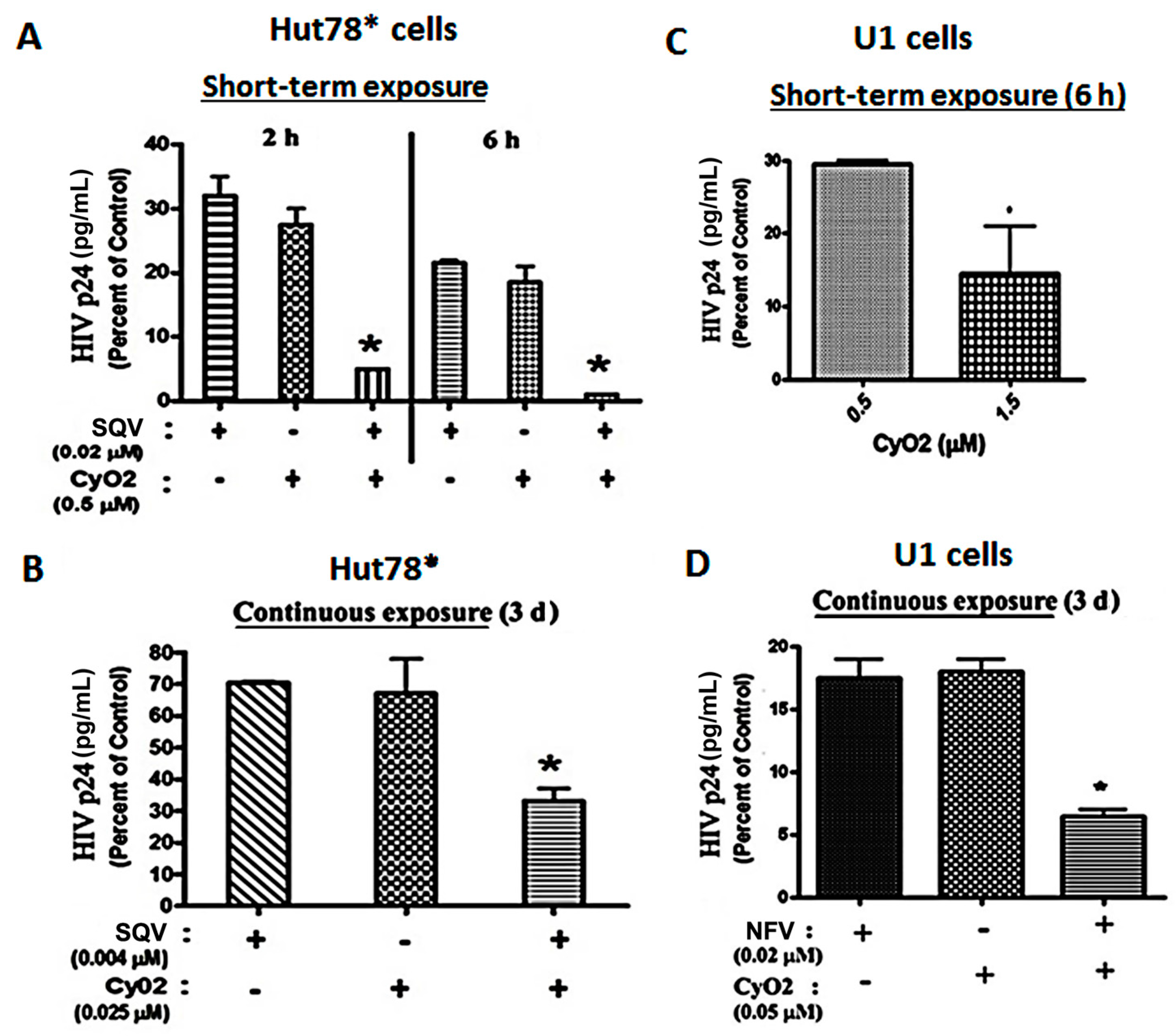{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue Culture
2.2. Reagents
2.3. Preparation of CFV and VLP Stocks
2.4. HIV-1 Infection and Treatment Protocols
2.5. Hemolytic Assays
2.6. SYTOX Green Uptake Assays
2.7. 3H-SQV Uptake Assays
2.8. Enzyme-Linked Immunosorbent Assays
2.9. HIV-1 LTR Reporter Assays
2.10. Infectivity Assays Using TZM-bl Cells
2.11. Statistical Analysis
3. Results
3.1. Low CyO2 Concentrations Do Not Cause Significant Hemolytic Activity on Human Red Blood Cells
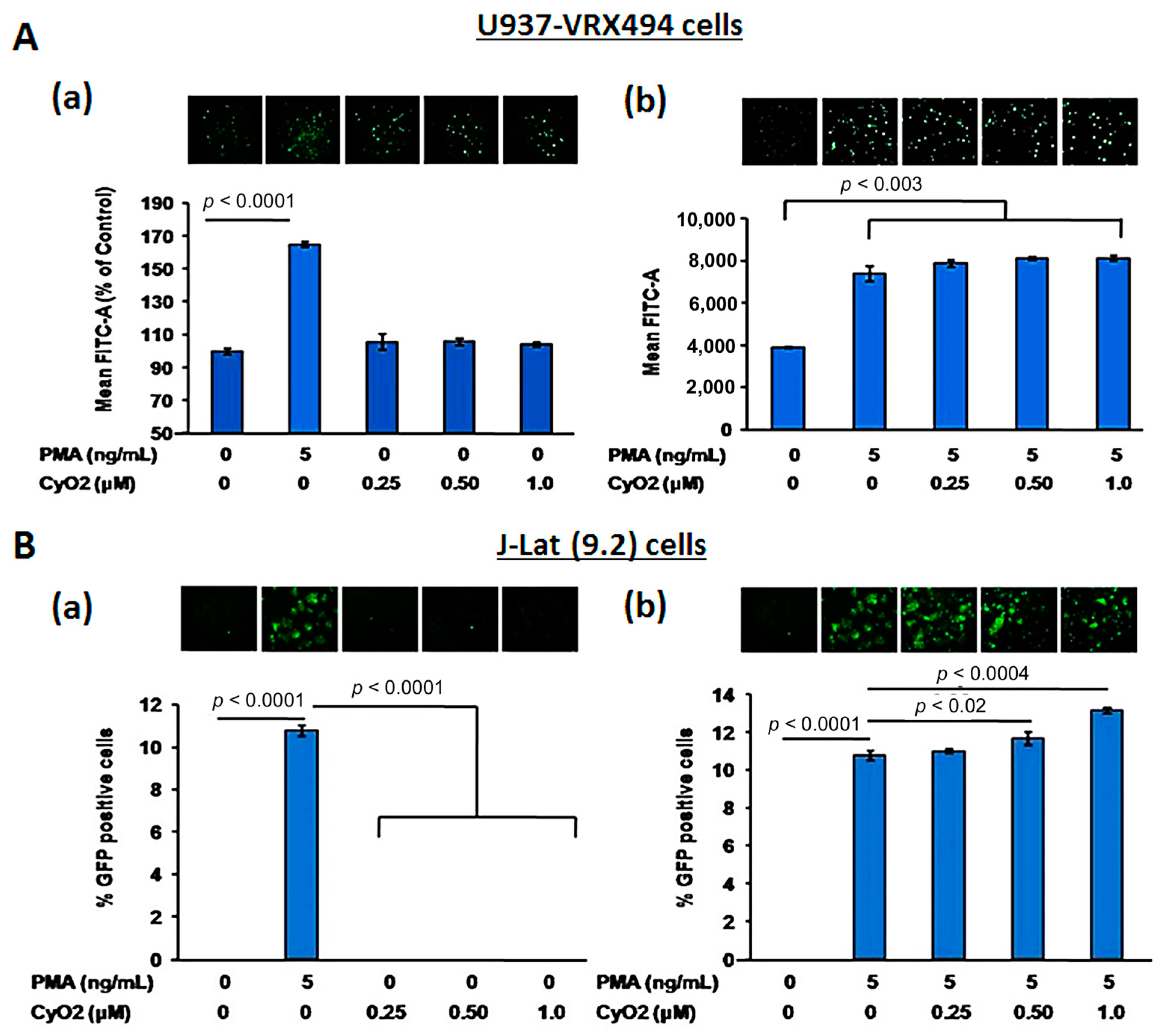
3.2. Non-Hemolytic Concentrations of CyO2 Retain Its Pore-Forming Ability and Induce Greater Pore-Formation in HIV-Infected Cells
3.3. CyO2 Facilitates Intracellular Uptake of Radiolabeled Saquinavir
3.4. Co-Exposure to CyO2 Impacts the Anti-HIV Efficacy of Multiple Protease Inhibitors
3.5. CyO2 Does Not Alter HIV-1 LTR Function
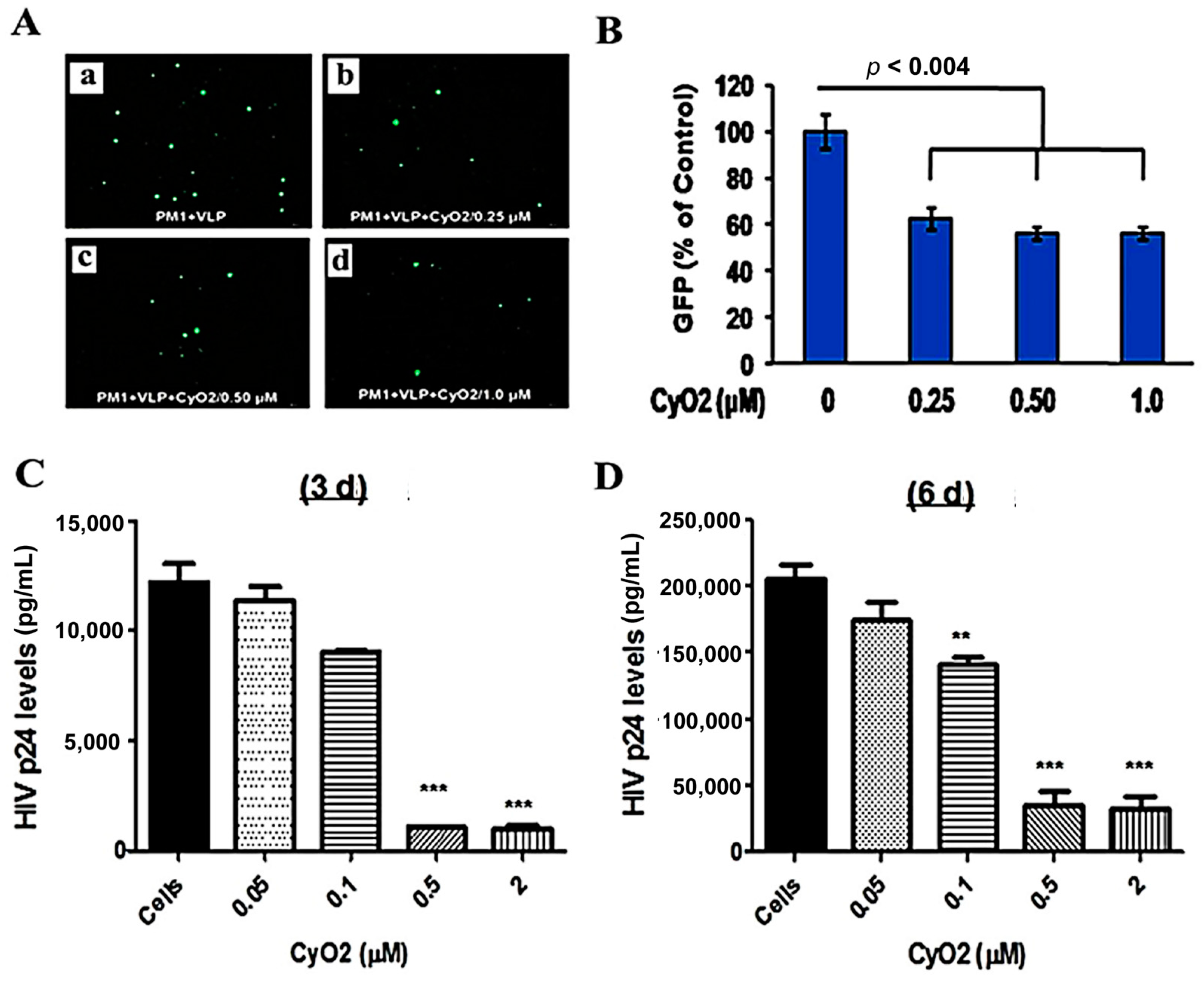
3.6. CyO2 Suppresses the Infectivity of Viral Particles
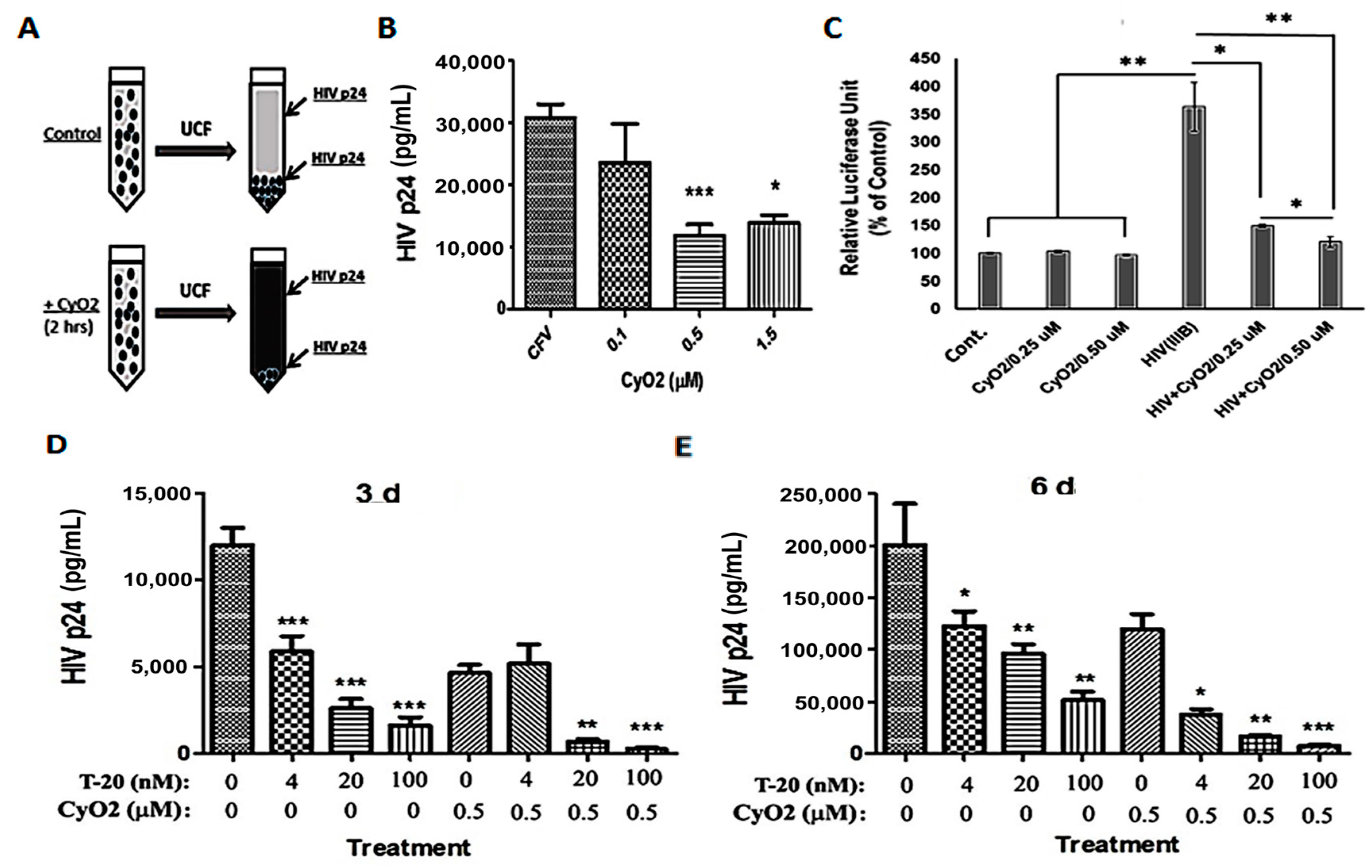
3.7. CyO2 Disrupts Viral Particles, Decreases Their Infectivity in the TZM-bl Model, and Increases the Anti-HIV Efficacy of Enfuvirtide
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- HIV/AIDS Basic Statistics. Available online: https://www.cdc.gov/hiv/basics/statistics.html (accessed on 19 November 2018).
- Song, R.; Hall, H.I.; Green, T.A.; Szwarcwald, C.L.; Pantazis, N. Using CD4 Data to Estimate HIV Incidence, Prevalence, and Percent of Undiagnosed Infections in the United States. J. Acquir. Immune Defic. Syndr. 2017, 74, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Bärnighausen, T.; Bloom, D.E.; Humair, S. Human Resources for Treating HIV/AIDS: Are the Preventive Effects of Antiretroviral Treatment a Game Changer? PLoS ONE 2016, 11, e0163960. [Google Scholar] [CrossRef] [PubMed]
- Kurapati, K.R.; Atluri, V.S.; Samikkannu, T.; Garcia, G.; Nair, M.P. Natural Products as Anti-HIV Agents and Role in HIV-Associated Neurocognitive Disorders (HAND): A Brief Overview. Front. Microbiol. 2016, 6, 1444. [Google Scholar] [CrossRef] [PubMed]
- Zofou, D.; Ntie-Kang, F.; Sippl, W.; Efange, S.M. Bioactive natural products derived from the Central African flora against neglected tropical diseases and HIV. Nat. Prod. Rep. 2013, 30, 109. [Google Scholar] [CrossRef] [PubMed]
- Grant, P.M.; Zolopa, A.R. Optimal antiretroviral therapy: HIV-1 treatment strategies to avoid and overcome drug resistance. Curr. Opin. Investig. Drugs 2010, 11, 901–910. [Google Scholar] [PubMed]
- Roy, U.; Bulot, C.; Honer zu Bentrup, K.; Mondal, D. Specific increase in MDR1 mediated drug-efflux in human brain endothelial cells following co-exposure to HIV-1 and saquinavir. PLoS ONE 2013, 8, e75374. [Google Scholar] [CrossRef] [PubMed]
- Okoye, A.A.; Picker, L.J. CD4(+) T-cell depletion in HIV infection: Mechanisms of immunological failure. Immunol. Rev. 2013, 254, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Cassol, E.; Alfano, M.; Biswas, P.; Poli, G. Monocyte-derived macrophages and myeloid cell lines as targets of HIV-1 replication and persistence. J. Leukoc. Biol. 2006, 80, 1018–1030. [Google Scholar] [CrossRef] [PubMed]
- Fumakia, M.; Yang, S.; Gu, J.; Ho, E.A. Protein/peptide-based entry/fusion inhibitors as anti-HIV therapies: Challenges and future direction. Rev. Med. Virol. 2016, 26, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Le Grice, S.F. Human immunodeficiency virus reverse transcriptase: 25 years of research, drug discovery, and promise. J. Biol. Chem. 2012, 287, 40850–40857. [Google Scholar] [CrossRef] [PubMed]
- Mesplède, T.; Quashie, P.K.; Zanichelli, V.; Wainberg, M.A. Integrase strand transfer inhibitors in the management of HIV-positive individuals. Ann. Med. 2014, 46, 123–129. [Google Scholar] [CrossRef] [PubMed]
- Kilareski, E.M.; Shah, S.; Nonnemacher, M.R.; Wigdahl, B. Regulation of HIV-1 transcription in cells of the monocyte-macrophage lineage. Retrovirology 2009, 6, 118. [Google Scholar] [CrossRef] [PubMed]
- Tejerina, F.; Bernaldo de Quirós, J.C. Protease inhibitors as preferred initial regimen for antiretroviral-naive HIV patients. AIDS Rev. 2011, 13, 227–233. [Google Scholar] [PubMed]
- Campbell, S.M.; Crowe, S.M.; Mak, J. Lipid rafts and HIV-1: From viral entry to assembly of progeny virions. J. Clin. Virol. 2001, 22, 217–227. [Google Scholar] [CrossRef]
- Yang, S.T.; Kiessling, V.; Simmons, J.A.; White, J.M.; Tamm, L.K. HIV gp41-mediated membrane fusion occurs at edges of cholesterol-rich lipid domains. Nat. Chem. Biol. 2015, 11, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Tang, M.W.; Shafer, R.W. HIV-1 antiretroviral resistance: Scientific principles and clinical applications. Drugs 2012, 72, e1–e25. [Google Scholar] [CrossRef] [PubMed]
- Zha, W.; Zha, B.S.; Zhou, F.; Zhou, H.; Wang, G. The cellular pharmacokinetics of HIV protease inhibitors: Current knowledge and future perspectives. Curr. Drug Metab. 2012, 13, 1174–1183. [Google Scholar] [CrossRef] [PubMed]
- Taiwo, B.; Hicks, C.; Eron, J. Unmet therapeutic needs in the new era of combination antiretroviral therapy for HIV-1. J. Antimicrob. Chemother. 2010, 65, 1100–1107. [Google Scholar] [CrossRef] [PubMed]
- Roy, U.; Barber, P.; Tse-Dinh, Y.C.; Batrakova, E.V.; Mondal, D.; Nair, M. Role of MRP transporters in regulating antimicrobial drug inefficacy and oxidative stress-induced pathogenesis during HIV-1 and TB infections. Front. Microbiol. 2015, 6, 948. [Google Scholar] [CrossRef] [PubMed]
- Eilers, M.; Roy, U.; Mondal, D. MRP (ABCC) transporters-mediated efflux of anti-HIV drugs, saquinavir and zidovudine, from human endothelial cells. Exp. Biol. Med. 2008, 233, 1149–1160. [Google Scholar] [CrossRef] [PubMed]
- Chandra, S.; Mondal, D.; Agrawal, K.C. HIV-1 protease inhibitor induced oxidative stress suppresses glucose stimulated insulin release: Protection with thymoquinone. Exp. Biol. Med. 2009, 234, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Blankson, J.N. Control of HIV-1 replication in elite suppressors. Discov. Med. 2010, 9, 261–266. [Google Scholar] [PubMed]
- Langford, S.E.; Ananworanich, J.; Cooper, D.A. Predictors of disease progression in HIV infection: A review. AIDS Res. Ther. 2007, 14, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Cos, P.; Maes, L.; Vanden Berghe, D.; Hermans, N.; Pieters, L.; Vlietinck, A. Plant substances as anti-HIV agents selected according to their putative mechanism of action. J. Nat. Prod. 2004, 67, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Asres, K.; Seyoum, A.; Veeresham, C.; Bucar, F.; Gibbons, S. Naturally derived anti-HIV agents. Phytother. Res. 2005, 19, 557–581. [Google Scholar] [CrossRef] [PubMed]
- Ireland, D.C.; Wang, C.K.; Wilson, J.A.; Gustafson, K.R.; Craik, D.J. Cyclotides as natural anti-HIV agents. Biopolymers 2008, 90, 51–60. [Google Scholar] [CrossRef] [PubMed]
- Bokesch, H.R.; Pannell, L.K.; Cochran, P.K.; Sowder, R.C., 2nd; McKee, T.C.; Boyd, M.R. A novel anti-HIV macrocyclic peptide from Palicourea condensata. J. Nat. Prod. 2001, 64, 249–250. [Google Scholar] [CrossRef] [PubMed]
- Hallock, Y.F.; Sowder, R.C., 2nd; Pannell, L.K.; Hughes, C.B.; Johnson, D.G.; Gulakowski, R.; Cardellina, J.H.; Boyd, M.R. Cycloviolins A-D, anti-HIV macrocyclic peptides from Leonia cymosa. J. Org. Chem. 2000, 65, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Huang, Y.H.; Rosengren, K.J.; Franquelim, H.G.; Carvalho, F.A.; Johnson, A.; Sonza, S.; Tachedjian, G.; Castanho, M.A.R.B.; Daly, N.L.; et al. Decoding the membrane activity of the cyclotide kalata B1: The importance of phosphatidylethanolamine phospholipids and lipid organization on hemolytic and anti-HIV activities. J. Biol. Chem. 2011, 286, 24231–24241. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Huang, Y.H.; Castanho, M.A.; Bagatolli, L.A.; Sonza, S.; Tachedjian, G.; Daly, N.L.; Craik, D.J. Phosphatidylethanolamine binding is a conserved feature of cyclotide-membrane interactions. J. Biol. Chem. 2012, 287, 33629–33643. [Google Scholar] [CrossRef] [PubMed]
- Malik, S.Z.; Linkevicius, M.; Göransson, U.; Andersson, D.I. Resistance to the Cyclotide Cycloviolacin O2 in Salmonella enterica Caused by Different Mutations that Often Confer Cross-Resistance or Collateral Sensitivity to Other Antimicrobial Peptides. Antimicrob. Agents Chemother. 2017, 61, e00684-17. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.H.; Colgrave, M.L.; Daly, N.L.; Keleshian, A.; Martinac, B.; Craik, D.J. The biological activity of the prototypic cyclotide kalata b1 is modulated by the formation of multimeric pores. J. Biol. Chem. 2009, 284, 20699–20707. [Google Scholar] [CrossRef] [PubMed]
- Gustafson, K.R.; Sowder, R.C.; Henderson, L.E.; Parsons, I.C.; Kashman, Y.; Cardellina, J.H., II; McMahon, J.B.; Buckheit, R.W., Jr.; Pannell, L.K. Circulins A and B. Novel human immunodeficiency virus (HIV)-inhibitory macrocyclic peptides from the tropical tree Chassalia parvifolia. J. Am. Chem. Soc. 1994, 116, 9337–9338. [Google Scholar] [CrossRef]
- Chen, B.; Colgrave, M.L.; Daly, N.L.; Rosengren, K.J.; Gustafson, K.R.; Craik, D.J. Isolation and characterization of novel cyclotides from Viola hederaceae: Solution structure and anti-HIV activity of vhl-1, a leaf-specific expressed cyclotide. J. Biol. Chem. 2005, 280, 22395–22405. [Google Scholar] [CrossRef] [PubMed]
- Daly, N.L.; Gustafson, K.R.; Craik, D.J. The role of the cyclic peptide backbone in the anti-HIV activity of the cyclotide kalata B1. FEBS Lett. 2004, 574, 69–72. [Google Scholar] [CrossRef] [PubMed]
- Daly, N.L.; Clark, R.J.; Plan, M.R.; Craik, D.J. Kalata B8, a novel antiviral circular protein, exhibits conformational flexibility in the cystine knot motif. Biochem. J. 2006, 393, 619–626. [Google Scholar] [CrossRef] [PubMed]
- Svangård, E.; Burman, R.; Gunasekera, S.; Lövborg, H.; Gullbo, J.; Göransson, U. Mechanism of action of cytotoxic cyclotides: Cycloviolacin O2 disrupts lipid membranes. J. Nat. Prod. 2007, 70, 643–647. [Google Scholar] [CrossRef] [PubMed]
- Pränting, M.; Lööv, C.; Burman, R.; Göransson, U.; Andersson, D.I. The cyclotide cycloviolacin O2 from Viola odorata has potent bactericidal activity against Gram-negative bacteria. J. Antimicrob. Chemother. 2010, 65, 1964–1971. [Google Scholar] [CrossRef] [PubMed]
- Burman, R.; Strömstedt, A.A.; Malmsten, M.; Göransson, U. Cyclotide-membrane interactions: Defining factors of membrane binding, depletion and disruption. Biochim. Biophys. Acta 2011, 11, 2665–2673. [Google Scholar] [CrossRef] [PubMed]
- Ghanam, R.H.; Samal, A.B.; Fernandez, T.F.; Saad, J.S. Role of the HIV-1 Matrix Protein in Gag Intracellular Trafficking and Targeting to the Plasma Membrane for Virus Assembly. Front. Microbiol. 2012, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Huarte, N.; Carravilla, P.; Cruz, A.; Lorizate, M.; Nieto-Garai, J.A.; Kräusslich, H.G.; Pérez-Gil, J.; Requejo-Isidro, J.; Nieva, J.L. Functional organization of the HIV lipid envelope. Sci. Rep. 2016, 6, 34190. [Google Scholar] [CrossRef] [PubMed]
- Leung, K.; Kim, J.O.; Ganesh, L.; Kabat, J.; Schwartz, O.; Nabel, G.J. HIV-1 assembly: Viral glycoproteins segregate quantally to lipid rafts that associate individually with HIV-1 capsids and virions. Cell Host Microbe 2008, 3, 285–292. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, S.L.; Yeshak, M.; Göransson, U.; Roy, U.; Izadpanah, R.; Mondal, D. Cycloviolacin O2 (CyO2) suppresses productive infection and augments the antiviral efficacy of nelfinavir in HIV-1 infected monocytic cells. Biopolymers 2013, 5, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, S.L.; Rathinakumar, R.; Chakravarty, G.; Göransson, U.; Wimley, W.C.; Darwin, S.P.; Mondal, D. Anticancer and chemosensitizing abilities of cycloviolacin 02 from Viola odorata and psyle cyclotides from Psychotria leptothyrsa. Biopolymers 2010, 94, 617–625. [Google Scholar] [CrossRef] [PubMed]
- Graham, F.L.; van der Eb, A.J. A new technique for the assay of infectivity of human adenovirus 5 DNA. Virology 1973, 52, 456–467. [Google Scholar] [CrossRef]
- Humeau, L.M.; Binder, G.K.; Lu, X.; Slepushkin, V.; Merling, R.; Echeagaray, P.; Pereira, M.; Slepushkina, T.; Barnett, S.; Dropulic, L.K.; et al. Efficient lentiviral vector-mediated control of HIV-1 replication in CD4 lymphocytes from diverse HIV+ infected patients grouped according to CD4 count and viral load. Mol. Ther. 2004, 9, 902–913. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Song, H.; Austin, J.L.; Cheng, W. Optimized Infectivity of the Cell-Free Single-Cycle Human Immunodeficiency Viruses Type 1 (HIV-1) and Its Restriction by Host Cells. PLoS ONE 2013, 8, e67170. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, A.; Svangård, E.; Claeson, P.; Gullbo, J.; Bohlin, L.; Göransson, U. Key role of glutamic acid for the cytotoxic activity of the cyclotide cycloviolacin O2. Cell. Mol. Life Sci. 2006, 63, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Huang, Y.H.; Chaousis, S.; Wang, C.K.; Craik, D.J. Anticancer and toxic properties of cyclotides are dependent on phosphatidylethanolamine phospholipid targeting. ChemBioChem 2014, 15, 1956–1965. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, J.; Peters, P.J.; Clapham, P.R. Human immunodeficiency virus type 1 envelope glycoproteins that lack cytoplasmic domain cysteines: Impact on association with membrane lipid rafts and incorporation onto budding virus particles. J. Virol. 2004, 78, 5500–5506. [Google Scholar] [CrossRef] [PubMed]
- Mathur, A.; Abd Elmageed, Z.Y.; Liu, X.; Kostochka, M.L.; Zhang, H.; Abdel-Mageed, A.B.; Mondal, D. Subverting ER-stress towards apoptosis by nelfinavir and curcumin coexposure augments docetaxel efficacy in castration resistant prostate cancer cells. PLoS ONE 2014, 9, e103109. [Google Scholar] [CrossRef] [PubMed]
- Alissa, E.M.; Ferns, G.A. Functional foods and nutraceuticals in the primary prevention of cardiovascular diseases. J. Nutr. Metab. 2012, 2012, 569486. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Liu, S.; Pan, R.; Li, G.; Tang, H.; Jiang, M.; Xing, Y.; Jin, F.; Lin, L.; Dong, J. Curcumin attenuates gp120-Induced microglial inflammation by inhibiting autophagy via the PI3K pathway. Cell Mol. Neurobiol. 2018, 38, 1465–1477. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Craik, D.J. Cyclotide Structure and Function: The Role of Membrane Binding and Permeation. Biochemistry 2017, 56, 669–682. [Google Scholar] [CrossRef] [PubMed]
- Mulvenna, J.P.; Wang, C.; Craik, D.J. CyBase: A database of cyclic protein sequence and structure. Nucleic Acids Res. 2006, 34, D192–D194. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, S.L.; Göransson, U.; Kaas, Q.; Craik, D.J.; Mondal, D.; Gruber, C.W. A systematic approach to document cyclotide distribution in plant species from genomic, transcriptomic, and peptidomic analysis. Biopolymers 2013, 100, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Craik, D.J.; Daly, N.L.; Bond, T.; Waine, C. Plant cyclotides: A unique family of cyclic and knotted proteins that defines the cyclic cystine knot structural motif. J. Mol. Biol. 1999, 294, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Chahroudi, A.; Silvestri, G. Animal models to achieve an HIV cure. Curr. Opin. HIV AIDS 2016, 11, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Burman, R.; Svedlund, E.; Felth, J.; Hassan, S.; Herrmann, A.; Clark, R.J.; Craik, D.J.; Bohlin, L.; Claeson, P.; Göransson, U.; et al. Evaluation of toxicity and antitumor activity of cycloviolacin O2 in mice. Biopolymers 2010, 94, 626–634. [Google Scholar] [CrossRef] [PubMed]
- Esmaeili, M.A.; Abagheri-Mahabadi, N.; Hashempour, H.; Farhadpour, M.; Gruber, C.W.; Ghassempour, A. Viola plant cyclotide vigno 5 induces mitochondria-mediated apoptosis via cytochrome C release and caspases activation in cervical cancer cells. Fitoterapia 2016, 109, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Henriques, S.T.; Huang, Y.H.; Chaousis, S.; Sani, M.A.; Poth, A.G.; Separovic, F.; Craik, D.J. The Prototypic Cyclotide Kalata B1 Has a Unique Mechanism of Entering Cells. Chem. Biol. 2015, 8, 1087–1097. [Google Scholar] [CrossRef] [PubMed]
- Contreras, J.; Elnagar, A.Y.; Hamm-Alvarez, S.F.; Camarero, J.A. Cellular uptake of cyclotide MCoTI-I follows multiple endocytic pathways. J. Control. Release 2011, 55, 134–143. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.T.; Mischiati, C.; Ather, A.; Ohyama, T.; Dedachi, K.; Borgatti, M.; Kurita, N.; Gambari, R. Structure-based analysis of the molecular recognitions between HIV-1 TAR-RNA and transcription factor nuclear factor-kappaB (NFkB). Curr. Top. Med. Chem. 2012, 12, 814–827. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Darcis, G.; Van Lint, C.; Herbein, G. Epigenetic control of HIV-1 post integration latency: Implications for therapy. Clin. Epigenet. 2015, 7, 103. [Google Scholar] [CrossRef] [PubMed]








© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gerlach, S.L.; Chandra, P.K.; Roy, U.; Gunasekera, S.; Göransson, U.; Wimley, W.C.; Braun, S.E.; Mondal, D. The Membrane-Active Phytopeptide Cycloviolacin O2 Simultaneously Targets HIV-1-infected Cells and Infectious Viral Particles to Potentiate the Efficacy of Antiretroviral Drugs. Medicines 2019, 6, 33. https://doi.org/10.3390/medicines6010033
Gerlach SL, Chandra PK, Roy U, Gunasekera S, Göransson U, Wimley WC, Braun SE, Mondal D. The Membrane-Active Phytopeptide Cycloviolacin O2 Simultaneously Targets HIV-1-infected Cells and Infectious Viral Particles to Potentiate the Efficacy of Antiretroviral Drugs. Medicines. 2019; 6(1):33. https://doi.org/10.3390/medicines6010033
Chicago/Turabian StyleGerlach, Samantha L., Partha K. Chandra, Upal Roy, Sunithi Gunasekera, Ulf Göransson, William C. Wimley, Stephen E. Braun, and Debasis Mondal. 2019. "The Membrane-Active Phytopeptide Cycloviolacin O2 Simultaneously Targets HIV-1-infected Cells and Infectious Viral Particles to Potentiate the Efficacy of Antiretroviral Drugs" Medicines 6, no. 1: 33. https://doi.org/10.3390/medicines6010033
APA StyleGerlach, S. L., Chandra, P. K., Roy, U., Gunasekera, S., Göransson, U., Wimley, W. C., Braun, S. E., & Mondal, D. (2019). The Membrane-Active Phytopeptide Cycloviolacin O2 Simultaneously Targets HIV-1-infected Cells and Infectious Viral Particles to Potentiate the Efficacy of Antiretroviral Drugs. Medicines, 6(1), 33. https://doi.org/10.3390/medicines6010033