
Anti-Aging and Antioxidant Potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans Indicate a New Utilization for Roasted Seeds of Guarana

 , , , and
, , , and 
Abstract
:1. Introduction
2. Material and Methods
2.1. Plant Material and Extraction
2.2. Chemical Characterization of the Extract
2.2.1. Characterization of Alkaloids
2.2.2. Characterization of Catechins (ESI-MS/MS)
2.2.3. Total Phenol Content
2.3. Antioxidant Activity in Vitro (DPPH Assay)
2.4. Caenorhabditis Elegans: Strains and Culture
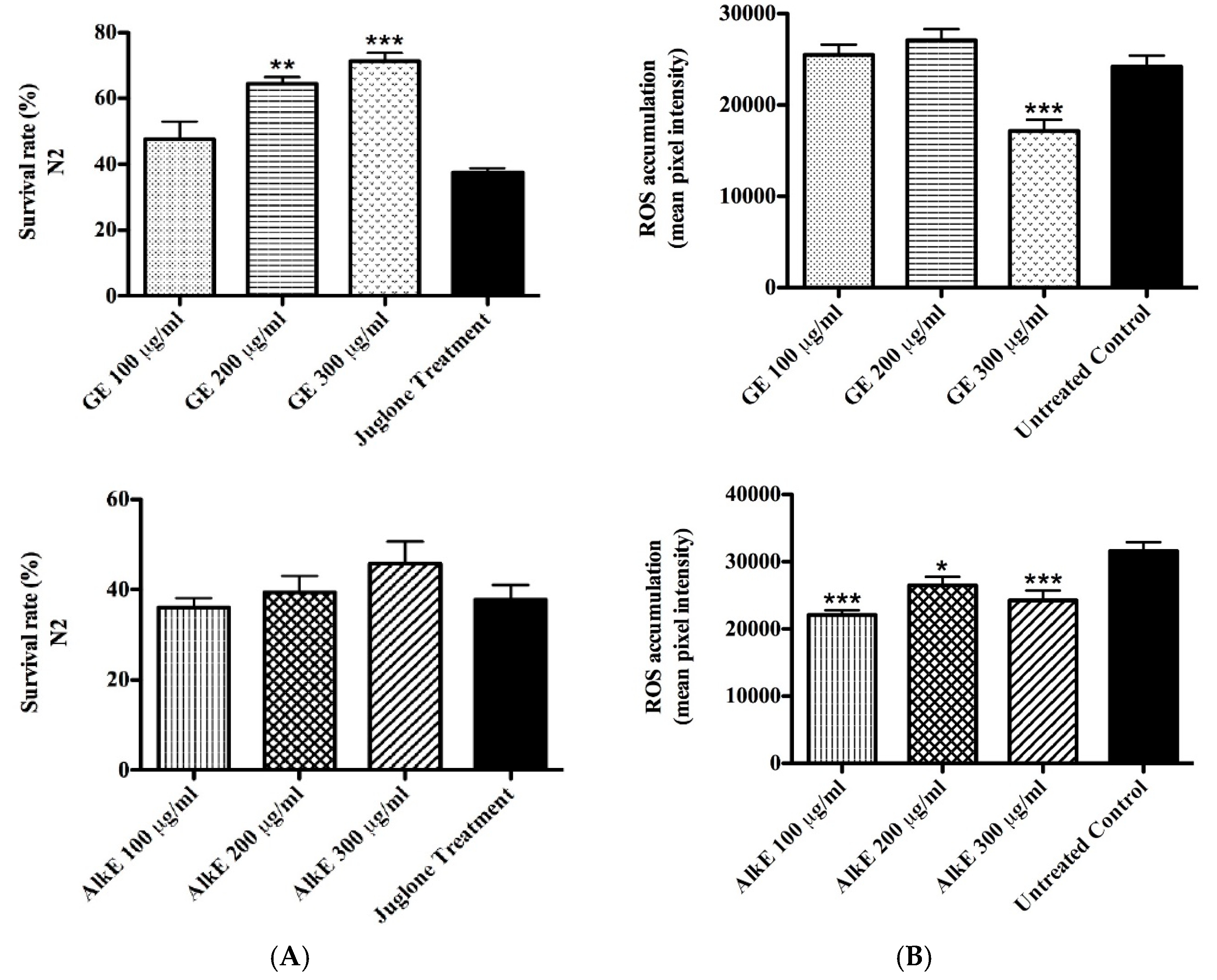
2.5. Antioxidant Activity in Vivo
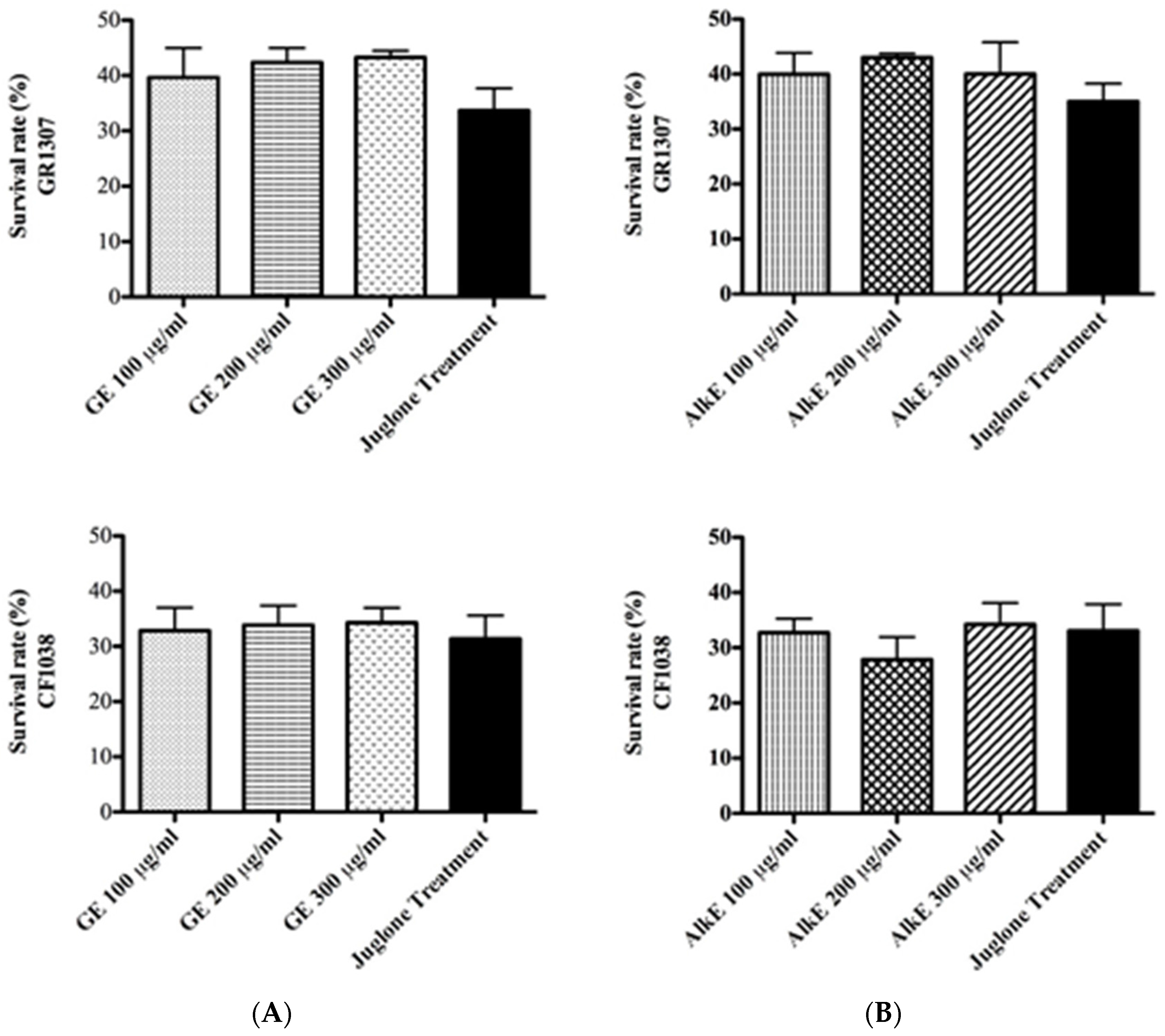
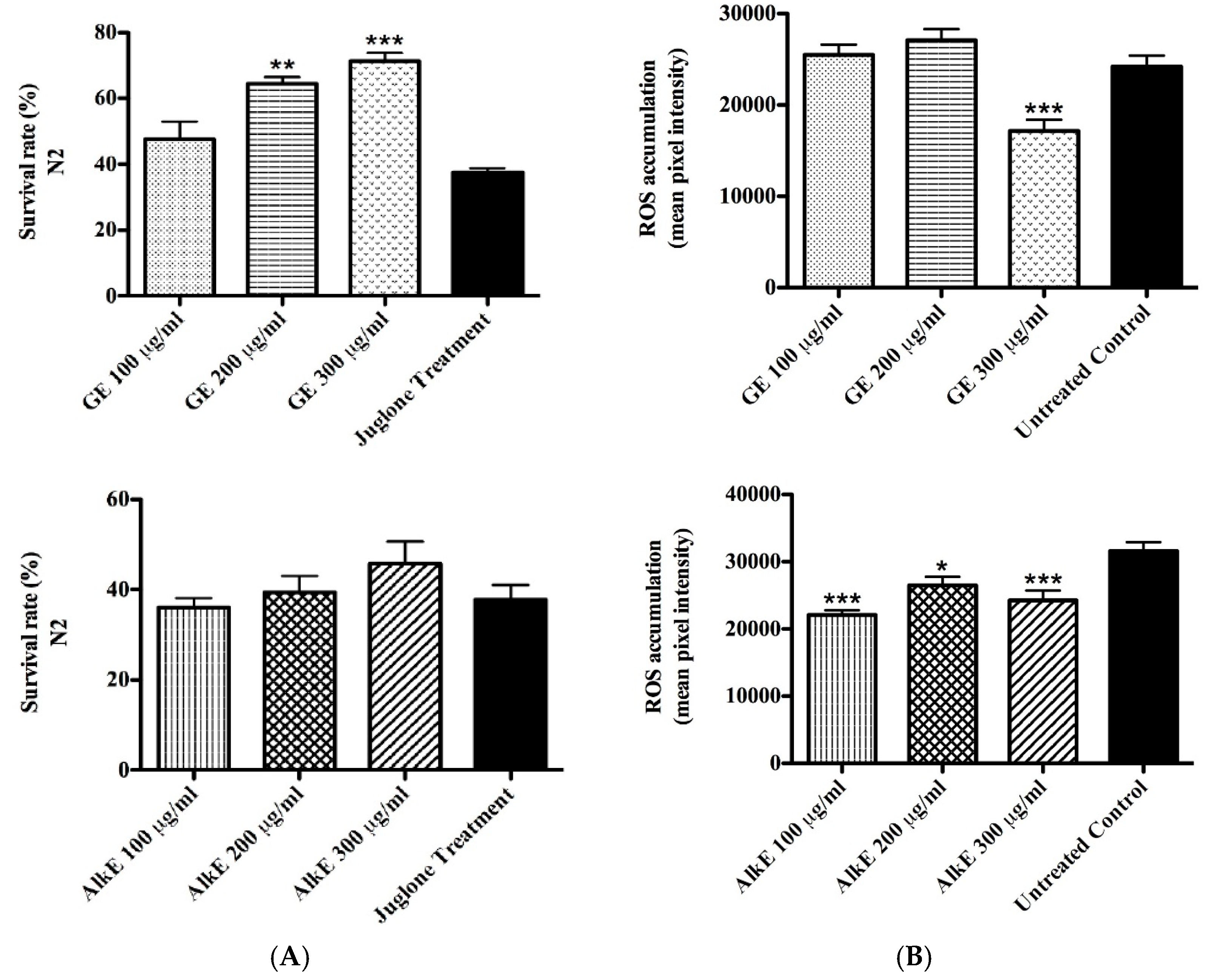
2.5.1. Survival Assay under Oxidative Stress
2.5.2. Intracellular ROS Accumulation
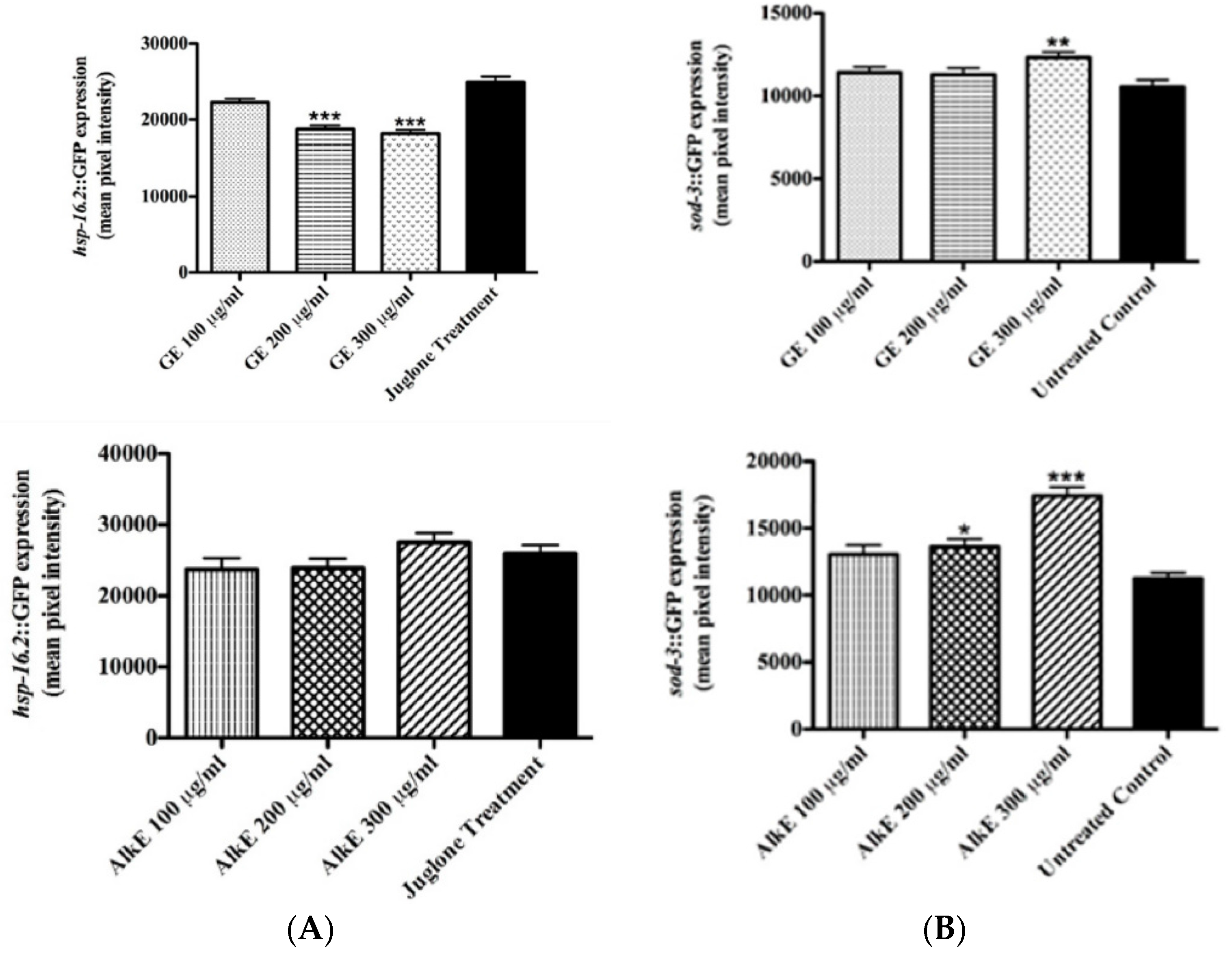
2.6. Expression of Stress Response Genes
2.6.1. Quantification of hsp-16.2::GFP Expression
2.6.2. Quantification of Sod-3::GFP Expression
2.7. Aging Markers
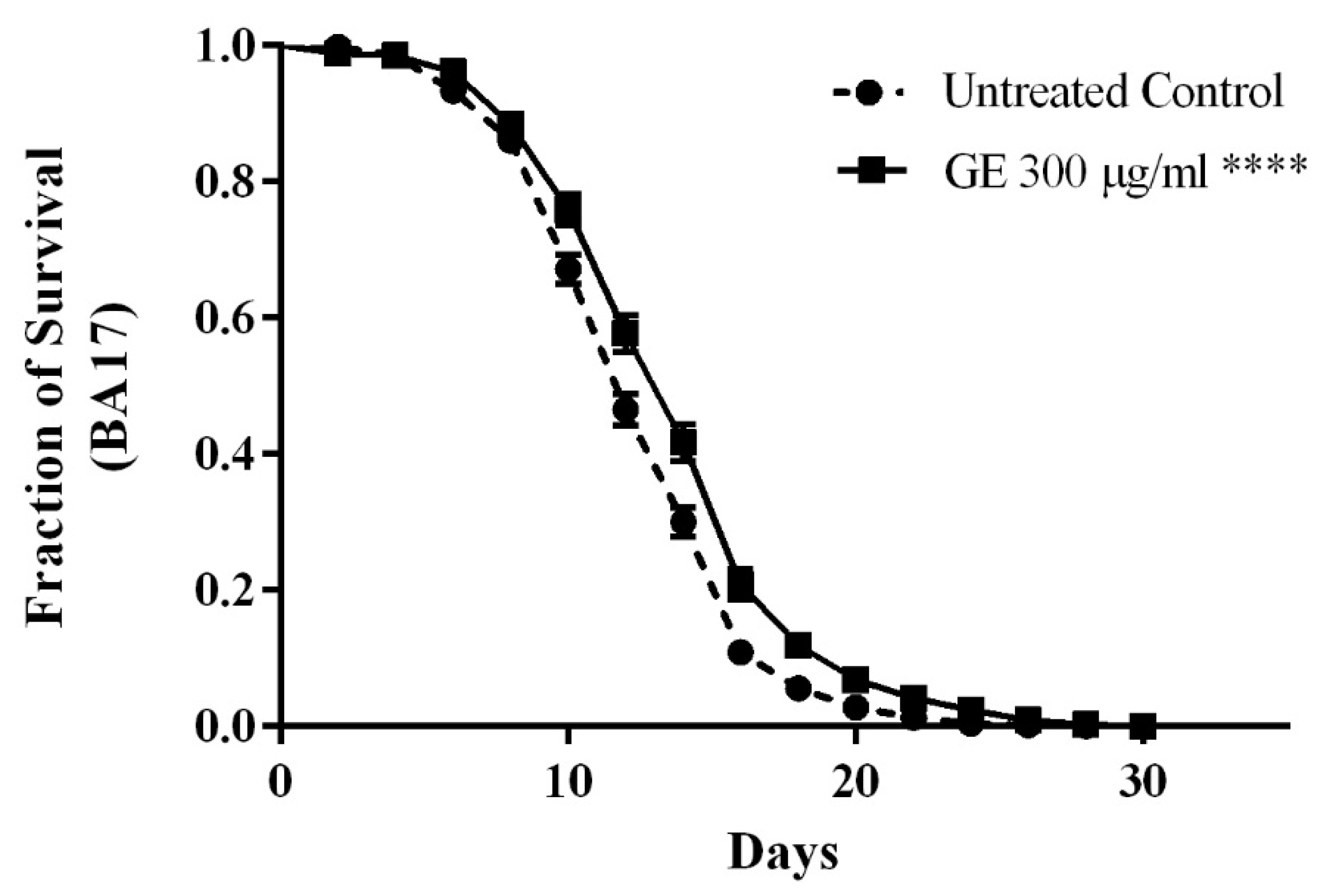
2.7.1. Longevity Assay
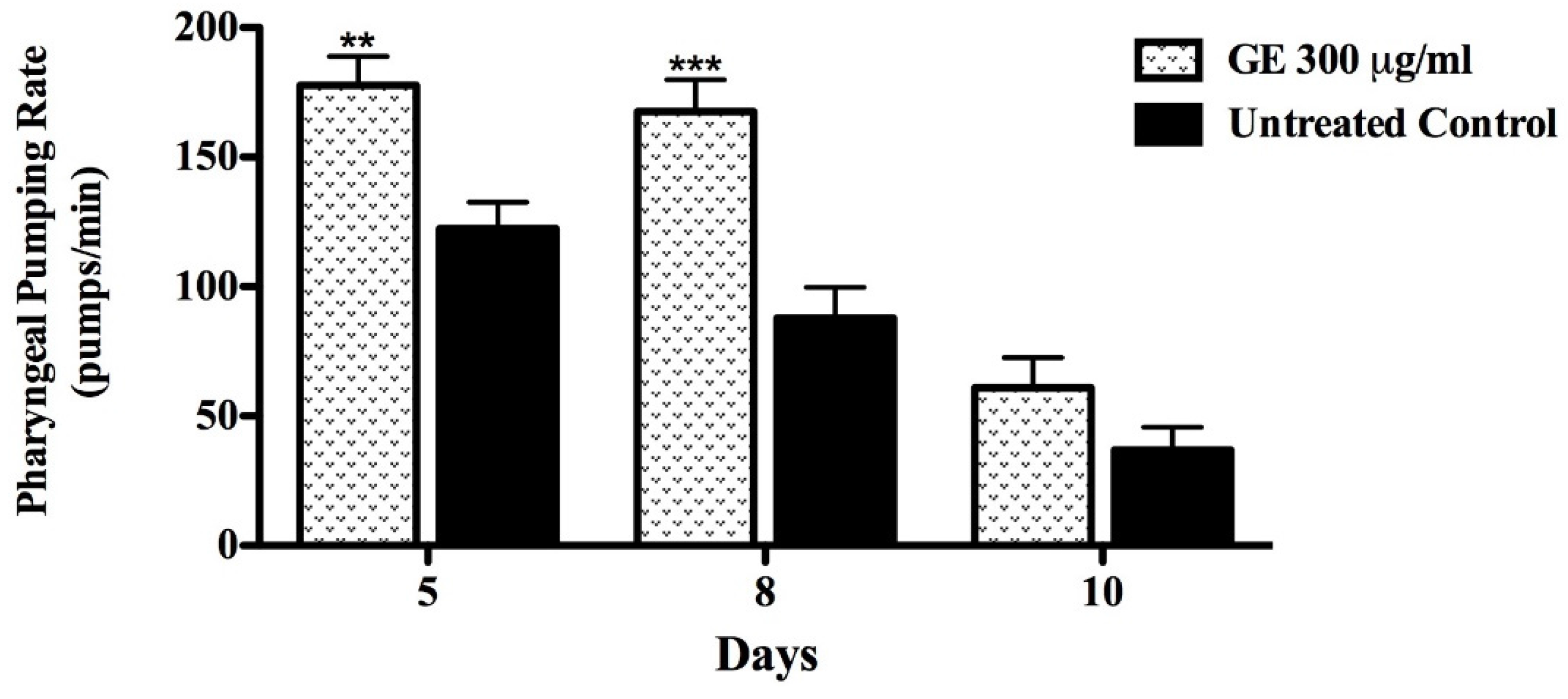
2.7.2. Pharyngeal Pumping Rate
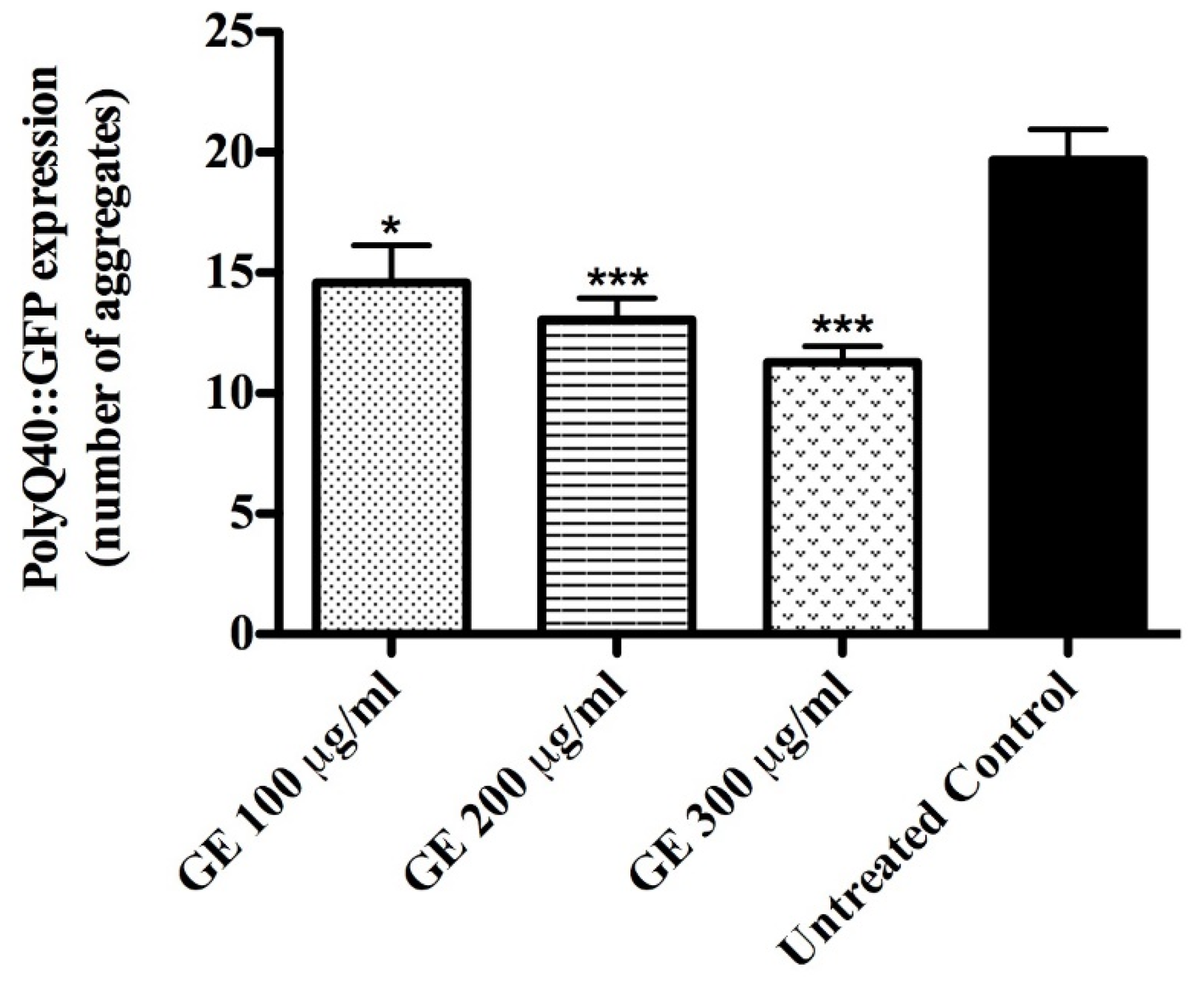
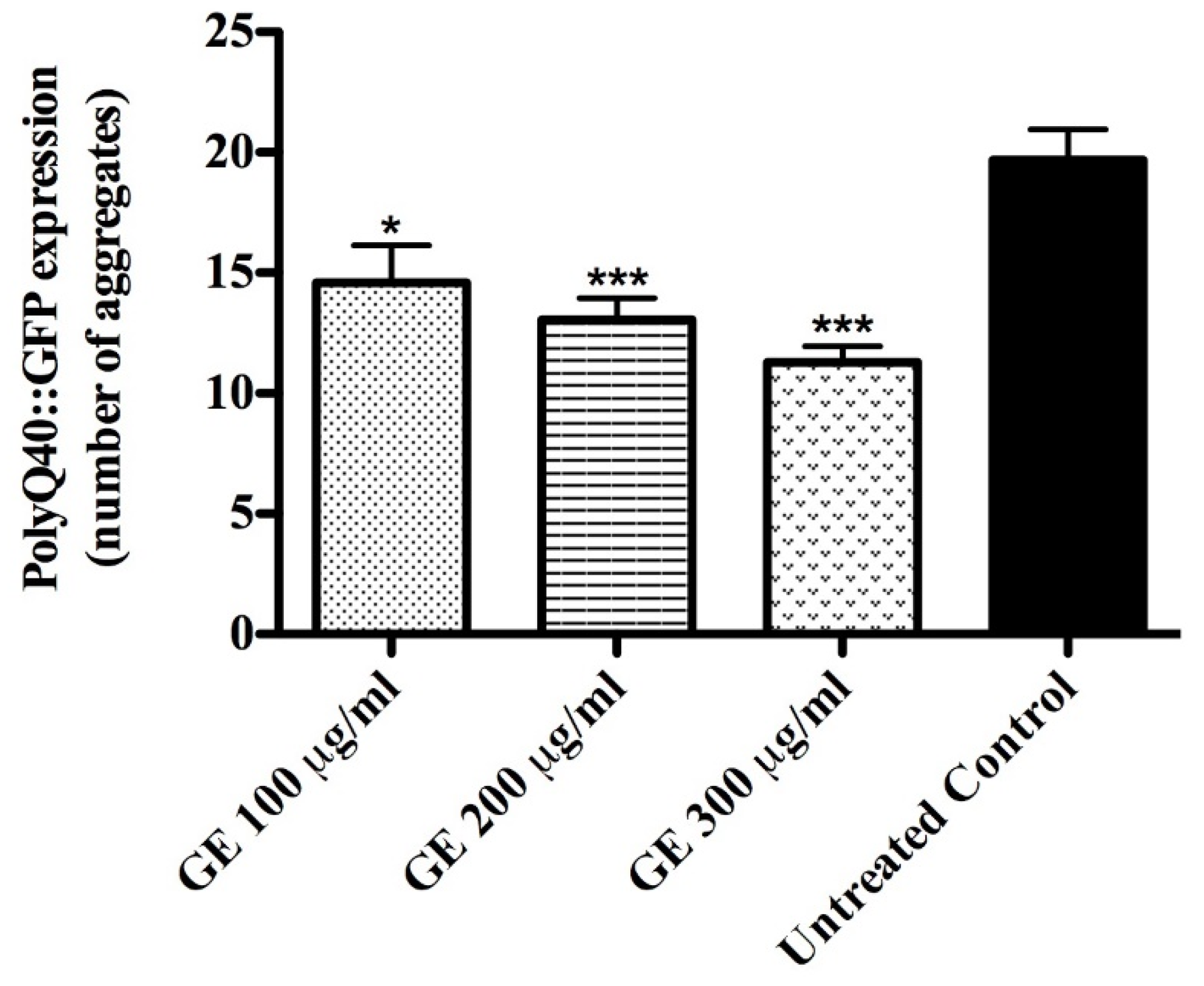
2.7.3. Quantification of PolyQ40::YFP Aggregates
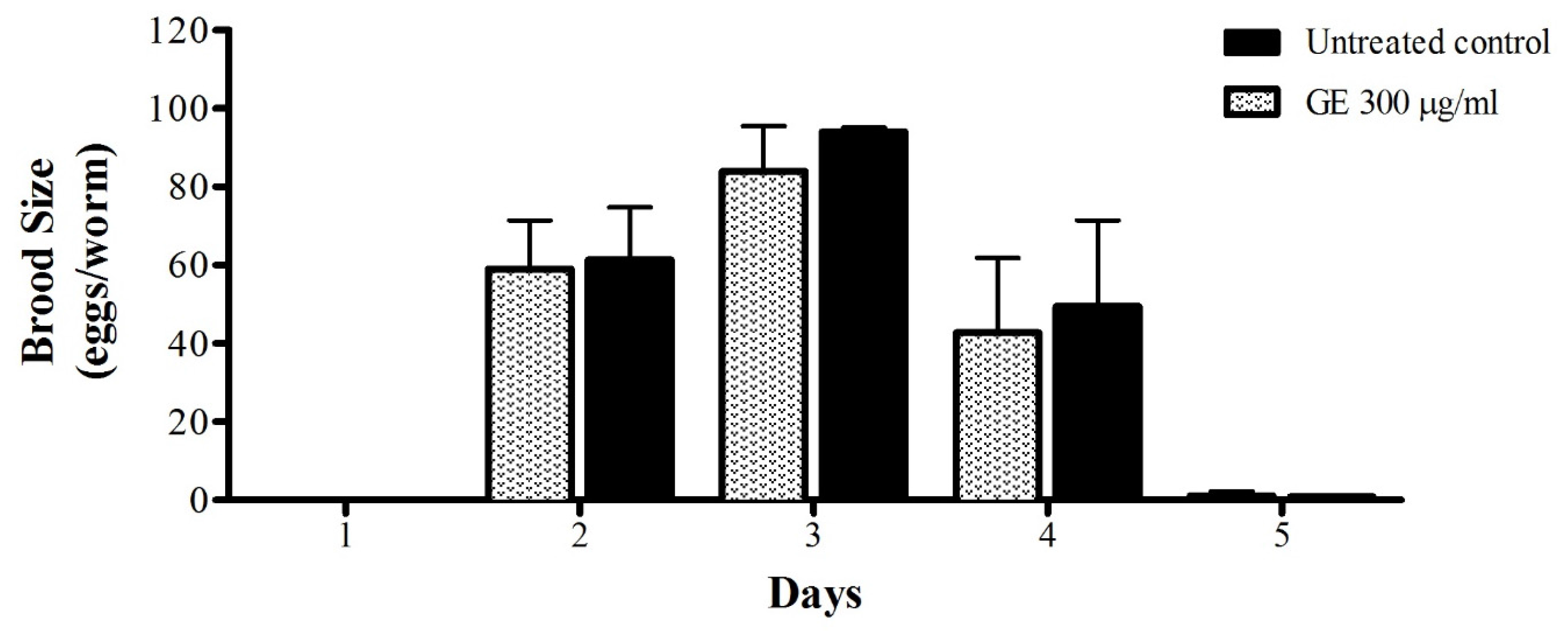
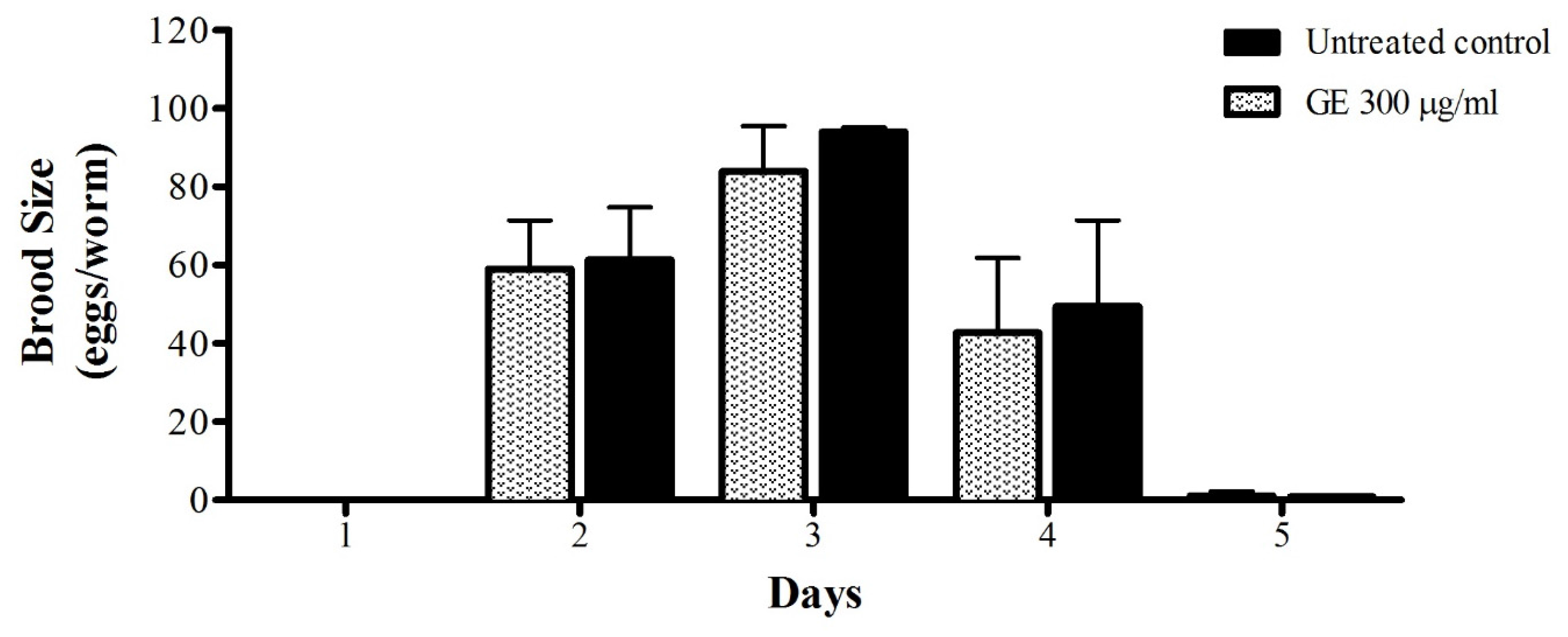
2.8. Toxicity Assay: C. Elegans Brood Size
3. Results and Discussion
3.1. Chemical Characterization of Guarana Extract
3.2. In Vitro Antioxidant Activity of Guarana
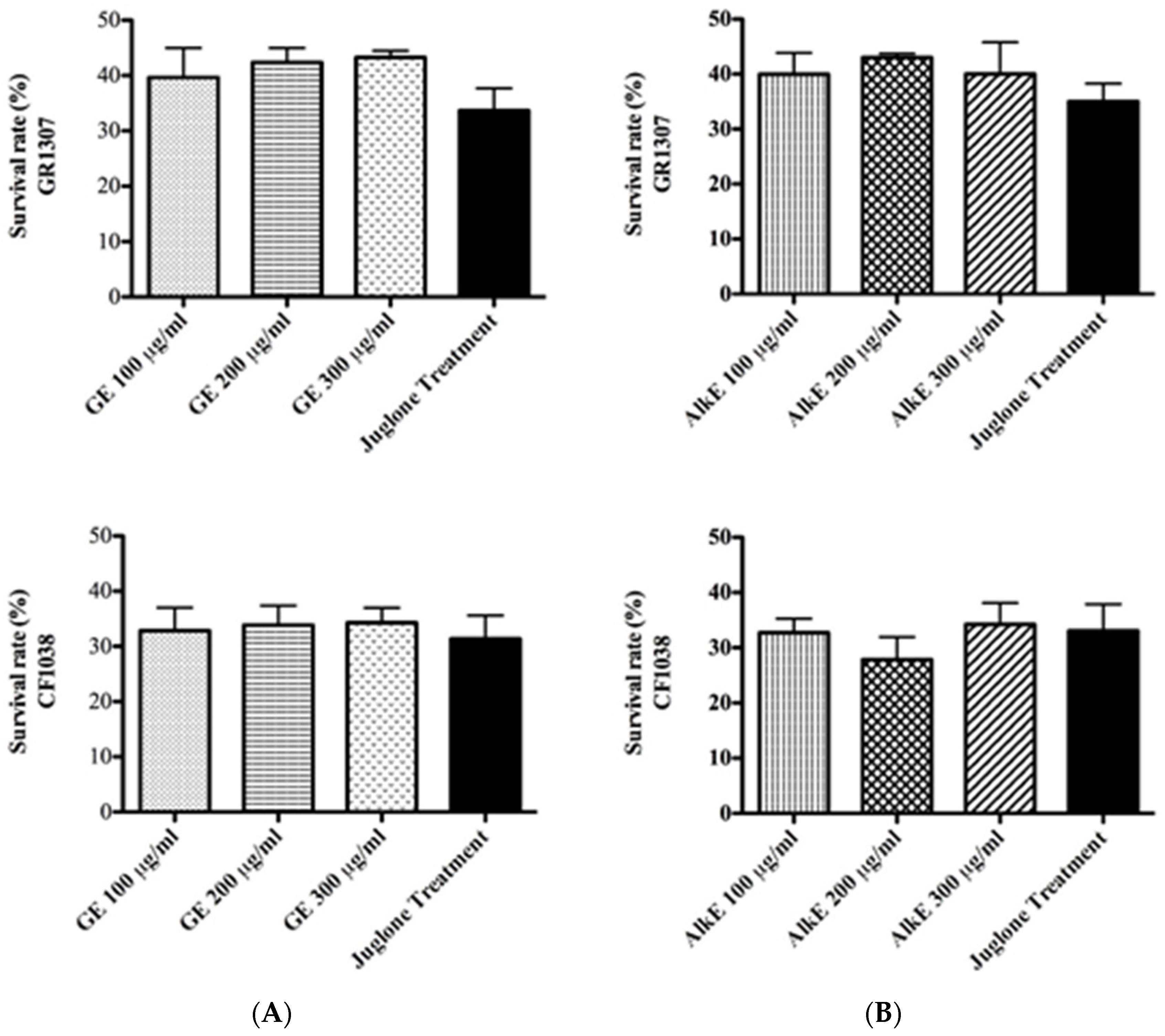
3.3. In Vivo Antioxidant Activity of Guarana
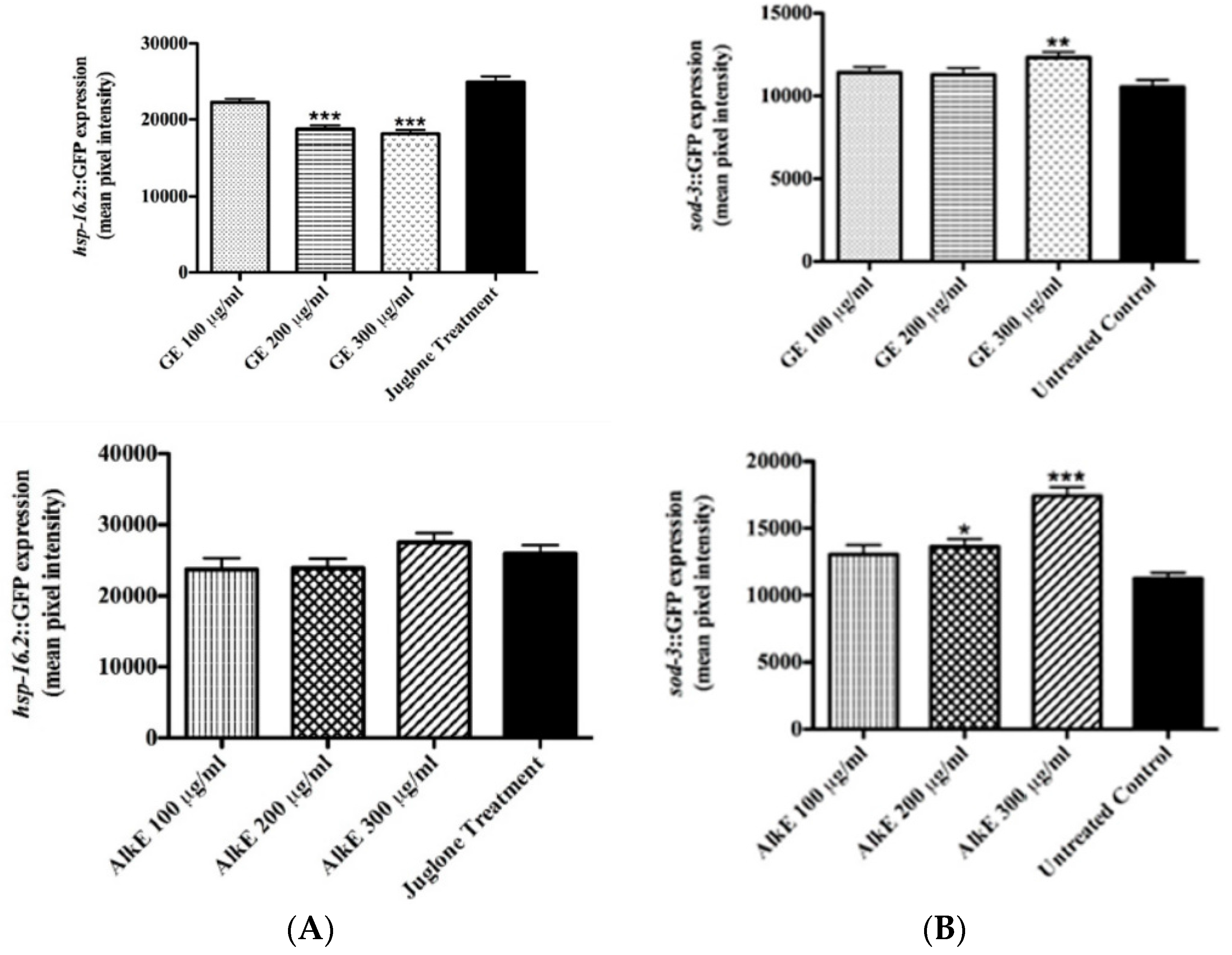
3.4. Quantification of the Expression of Stress Response Genes after Guarana Treatment
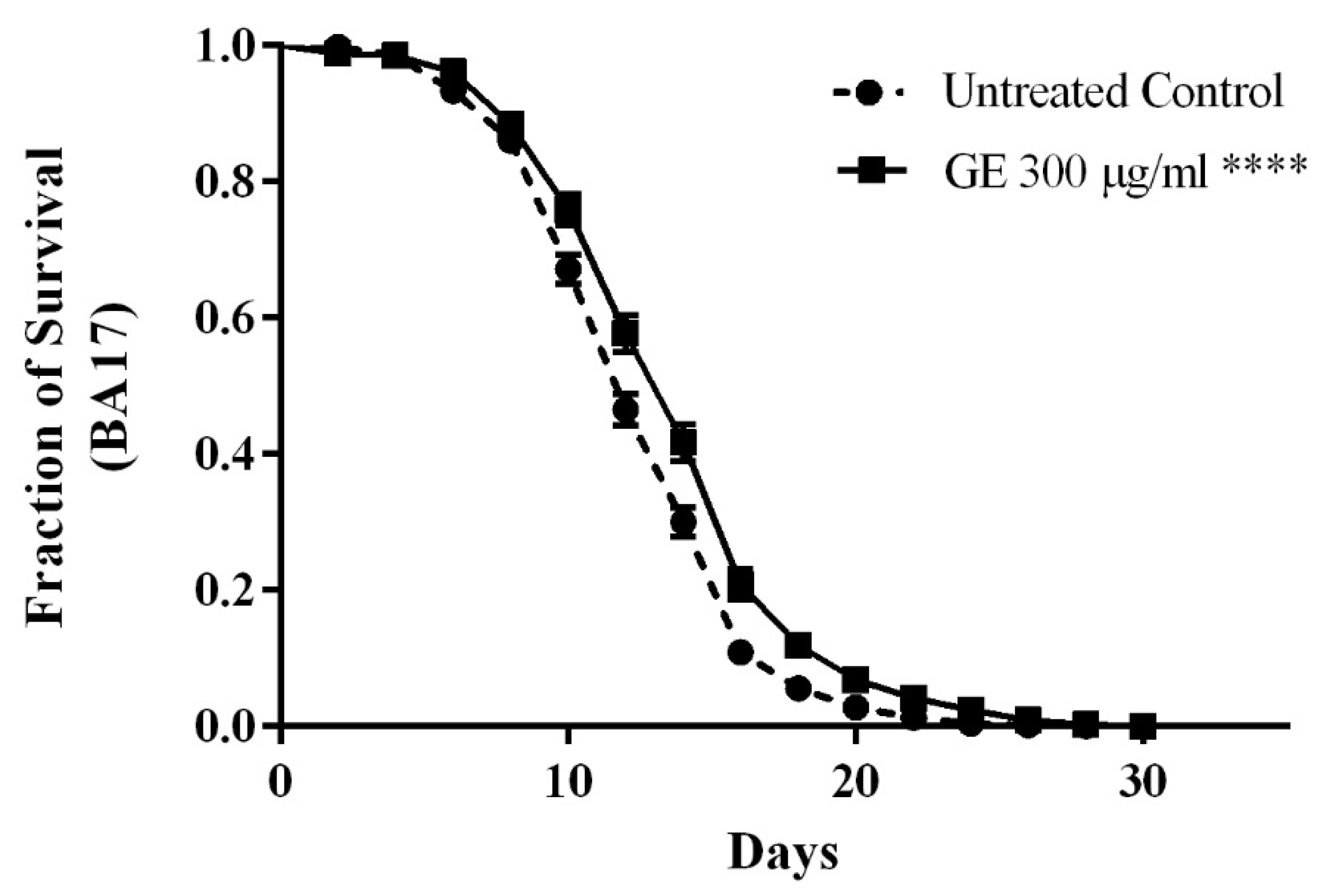
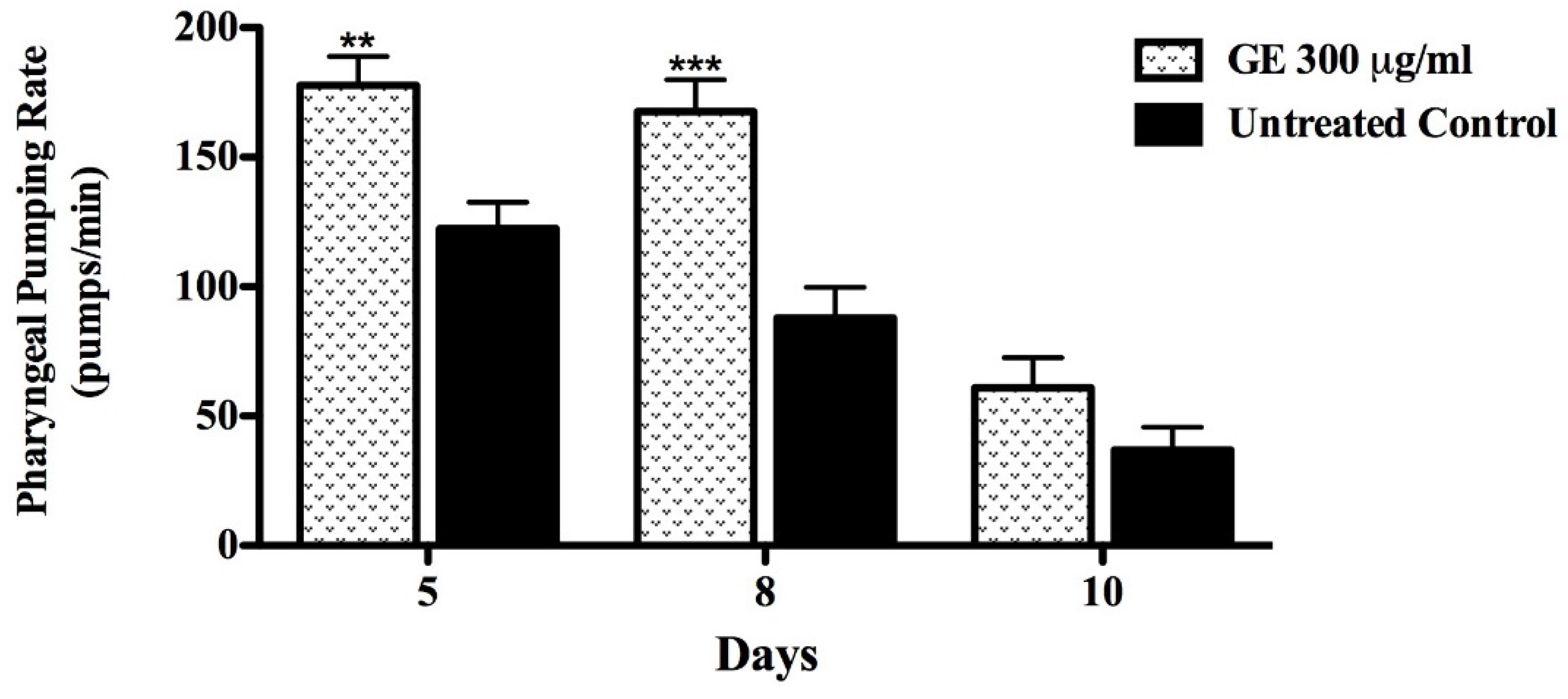
3.5. Anti-Aging effects
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Satoskar, R.; Rege, N.; Bhandarkar, S. Pharmacology and Pharmacotherapeutics; Elsevier Health Sciences APAC: Berlin, Germany, 2015. [Google Scholar]
- Wink, M.; Van Wyk, B.-E. Mind-Altering and Poisonous Plants of the World; Timber Press: Portland, OR, USA, 2008. [Google Scholar]
- Lüde, S.; Vecchio, S.; Sinno-Tellier, S.; Dopter, A.; Mustonen, H.; Vucinic, S.; Jonsson, B.; Müller, D.; Veras Gimenez Fruchtengarten, L.; Hruby, K. Adverse effects of plant food supplements and plants consumed as food: Results from the poisons centres-based plantlibra study. Phytother. Res. 2016, 30, 988–996. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.; Atroch, A.L. Guarana’s journey from regional tonic to aphrodisiac and global energy drink. Evid. Based Complement. Altern. Med. 2010, 7, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Henman, A.R. Guarana (Paullinia cupana var. sorbilis): Ecological and social perspectives on an economic plant of the central Amazon basin. J. Ethnopharmacol. 1982, 6, 311–338. [Google Scholar] [PubMed]
- Espinola, E.; Dias, R.; Mattei, R.; Carlini, E. Pharmacological activity of guarana (Paullinia cupana Mart.) in laboratory animals. J. Ethnopharmacol. 1997, 55, 223–229. [Google Scholar] [CrossRef]
- Otobone, F.J.; Sanches, A.C.; Nagae, R.L.; Martins, J.V.C.; Obici, S.; Mello, J.C.P.D.; Audi, E.A. Effect of crude extract and its semi purified constituents from guaraná seeds [Paullinia cupana var. sorbilis (Mart.) lucke] on cognitive performance in Morris water maze in rats. Braz. Arch. Biol. Technol. 2005, 48, 723–728. [Google Scholar]
- Otobone, F.; Sanches, A.; Nagae, R.; Martins, J.; Sela, V.; De Mello, J.; Audi, E. Effect of lyophilized extracts from guaraná seeds [Paullinia cupana var. sorbilis (Mart.) ducke] on behavioral profiles in rats. Phytother. Res. 2007, 21, 531–535. [Google Scholar] [PubMed]
- Rangel, M.P.; de Mello, J.C.P.; Audi, E.A. Evaluation of neurotransmitters involved in the anxiolytic and panicolytic effect of the aqueous fraction of Paullinia cupana (guaraná) in elevated t maze. Rev. Bras. Farmacogn. 2013, 23, 358–365. [Google Scholar] [CrossRef]
- Da Costa Krewer, C.; Ribeiro, E.E.; Ribeiro, E.A.M.; Moresco, R.N.; da Rocha, M.I.U.M.; Montagner, G.F.F.S.; Machado, M.M.; Viegas, K.; Brito, E.; Da cruz, I. Habitual intake of guaraná and metabolic morbidities: An epidemiological study of an elderly amazonian population. Phytother. Res. 2011, 25, 1367–1374. [Google Scholar] [CrossRef] [PubMed]
- Ruchel, J.; Rezer, J.; Thorstenberg, M.; Santos, C.; Cabral, F.; Lopes, S.; Silva, C.; Machado, A.; Cruz, I.; Schetinger, M. Hypercholesterolemia and ecto-enzymes of purinergic system: Effects of Paullinia cupana. Phytother. Res. 2016, 30, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Hamerski, L.; Somner, G.V.; Tamaio, N. Paullinia cupana Kunth (Sapindaceae): A review of its ethnopharmacology, phytochemistry and pharmacology. J. Med. Plants Res. 2013, 7, 2221–2229. [Google Scholar]
- Do Nascimento Carvalho, L.V.; Cordeiro, M.F.; e Lins, T.U.L.; Sampaio, M.C.P.D.; de Mello, G.S.V.; da Costa, V.D.C.M.; Marques, L.L.M.; Klein, T.; de Mello, J.C.P.; Cavalcanti, I.M.F.; et al. Evaluation of antibacterial, antineoplastic, and immunomodulatory activity of Paullinia cupana seeds crude extract and ethyl-acetate fraction. Evid. Based Complement. Altern. Med. 2016, 2016. [Google Scholar] [CrossRef]
- Marx, F. Analysis of guarana seeds ii.Studies on the composition of the tannin fraction. Zeitschrift für Lebensmittel-Untersuchung und Forschung 1990, 190, 429–431. [Google Scholar] [CrossRef]
- Meurer-Grimes, B.; Berkov, A.; Beck, H. Theobromine, theophylline, and caffeine in 42 samples and products of guarana (Paullinia cupana, Sapindaceae). Econ. Bot. 1998, 52, 293–301. [Google Scholar] [CrossRef]
- Van Wyk, B.-E.; Wink, M. Phytomedicines, Herbal Drugs, and Poisons; University of Chicago Press: Chicago, IL, USA, 2015. [Google Scholar]
- Peixoto, H.; Roxo, M.; Krstin, S.; Röhrig, T.; Richling, E.; Wink, M. An anthocyanin-rich extract of acai (Euterpe precatoria Mart.) increases stress resistance and retards aging-related markers in Caenorhabditis elegans. J. Agric. Food Chem. 2016, 64, 1283–1290. [Google Scholar] [PubMed]
- Abbas, S.; Wink, M. Green tea extract induces the resistance of Caenorhabditis elegans against oxidative stress. Antioxidants 2014, 3, 129–143. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Müller, D.; Richling, E.; Wink, M. Anthocyanin-rich purple wheat prolongs the life span of Caenorhabditis elegans probably by activating the daf-16/foxo transcription factor. J. Agric. Food Chem. 2013, 61, 3047–3053. [Google Scholar] [CrossRef] [PubMed]
- Bonoli, M.; Pelillo, M.; Toschi, T.G.; Lercker, G. Analysis of green tea catechins: Comparative study between HPLC and HPCE. Food Chem. 2003, 81, 631–638. [Google Scholar] [CrossRef]
- Blois, M. Antioxidant determinations by the use of a stable free radical. Nature 1958, 26, 1199–1200. [Google Scholar] [CrossRef]
- Stiernagle, T. Maintenance of C. elegans; Wormbook, Ed.; The C. elegans Research Community, Wormbook: Pasadena, CA, USA, 2006.
- Strange, K.; Christensen, M.; Morrison, R. Primary culture of Caenorhabditis elegans developing embryo cells for electrophysiological, cell biological and molecular studies. Nat. Protoc. 2007, 2, 1003–1012. [Google Scholar] [CrossRef] [PubMed]
- Yamaguti-Sasaki, E.; Ito, L.A.; Canteli, V.C.D.; Ushirobira, T.M.A.; Ueda-Nakamura, T.; Nakamura, C.V.; Palazzo de Mello, J.C. Antioxidant capacity and in vitro prevention of dental plaque formation by extracts and condensed tannins of Paullinia cupana. Molecules 2007, 12, 1950–1963. [Google Scholar] [CrossRef] [PubMed]
- Hesp, K.; Smant, G.; Kammenga, J.E. Caenorhabditis elegans daf-16/foxo transcription factor and its mammalian homologs associate with age-related disease. Exp. Gerontol. 2015, 72, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Strayer, A.; Wu, Z.; Christen, Y.; Link, C.D.; Luo, Y. Expression of the small heat-shock protein hsp 16-2 in Caenorhabditis elegans is suppressed by Ginkgo biloba extract EGb 761. FASEB J. 2003, 17, 2305–2307. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Sudji, I.R.; Wang, E.; Joubert, E.; van Wyk, B.-E.; Wink, M. Ameliorative effect of aspalathin from rooibos (Aspalathus linearis) on acute oxidative stress in Caenorhabditis elegans. Phytomedicine 2013, 20, 380–386. [Google Scholar] [CrossRef] [PubMed]
- Fontana, L.; Partridge, L.; Longo, V.D. Extending healthy life span—From yeast to humans. Science 2010, 328, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Rothman, J.H.; Singson, A. Caenorhabditis elegans: Cell Biology and Physiology; Academic Press: Cambridge, MA, USA, 2012; Volume 107. [Google Scholar]
- Herndon, L.A.; Schmeissner, P.J.; Dudaronek, J.M.; Brown, P.A.; Listner, K.M.; Sakano, Y.; Paupard, M.C.; Hall, D.H.; Driscoll, M. Stochastic and genetic factors influence tissue-specific decline in ageing. C. elegans. Nature 2002, 419, 808–814. [Google Scholar] [CrossRef] [PubMed]
- Chow, D.K.; Glenn, C.F.; Johnston, J.L.; Goldberg, I.G.; Wolkow, C.A. Sarcopenia in the Caenorhabditis elegans pharynx correlates with muscle contraction rate over lifespan. Exp. Gerontol. 2006, 41, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.F.; Dues, D.J.; Spielbauer, K.K.; Machiela, E.; Senchuk, M.M.; Van Raamsdonk, J.M. Delaying aging is neuroprotective in Parkinson’s disease: A genetic analysis in C. elegans models. NPJ Parkinson Dis. 2015, 1. [Google Scholar] [CrossRef]
- Bridi, J.C.; de Almeida Barros, A.G.; Sampaio, L.R.; Ferreira, J.C.D.; Soares, F.A.A.; Romano-Silva, M.A. Lifespan extension induced by caffeine in Caenorhabditis elegans is partially dependent on adenosine signaling. Front. Aging Neurosci. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Sutphin, G.L.; Bishop, E.; Yanos, M.E.; Moller, R.M.; Kaeberlein, M. Caffeine extends life span, improves healthspan, and delays age-associated pathology in Caenorhabditis elegans. Longev. Healthspan 2012, 1. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Concentration (mg/g) |
|---|---|
| Caffeine | 102.8 |
| Theophyline | 2.3 |
| Thebromine | 1.0 |
| Catechin | + |
| Epicatechin | + |
| Sample | DPPH * |
|---|---|
| GE | 40 |
| EGCG | 1.03 |
| Vitamin C | 2.12 |
| ALK | - |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peixoto, H.; Roxo, M.; Röhrig, T.; Richling, E.; Wang, X.; Wink, M. Anti-Aging and Antioxidant Potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans Indicate a New Utilization for Roasted Seeds of Guarana. Medicines 2017, 4, 61. https://doi.org/10.3390/medicines4030061
Peixoto H, Roxo M, Röhrig T, Richling E, Wang X, Wink M. Anti-Aging and Antioxidant Potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans Indicate a New Utilization for Roasted Seeds of Guarana. Medicines. 2017; 4(3):61. https://doi.org/10.3390/medicines4030061
Chicago/Turabian StylePeixoto, Herbenya, Mariana Roxo, Teresa Röhrig, Elke Richling, Xiaojuan Wang, and Michael Wink. 2017. "Anti-Aging and Antioxidant Potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans Indicate a New Utilization for Roasted Seeds of Guarana" Medicines 4, no. 3: 61. https://doi.org/10.3390/medicines4030061
APA StylePeixoto, H., Roxo, M., Röhrig, T., Richling, E., Wang, X., & Wink, M. (2017). Anti-Aging and Antioxidant Potential of Paullinia cupana var. sorbilis: Findings in Caenorhabditis elegans Indicate a New Utilization for Roasted Seeds of Guarana. Medicines, 4(3), 61. https://doi.org/10.3390/medicines4030061