
Glyphosate in Runoff Waters and in the Root-Zone: A Review

Abstract
:1. Introduction
1.1. Background
1.2. Prevalence

{kind=link}
{kind=link}
{kind=link}
| Agricultural Sector | Amount Applied in Surveyed Year (kg) | % of Planted Hectares | Year Surveyed |
|---|---|---|---|
| Soybeans | 45,530,000 | 89 | 2012 |
| Corn | 2,610,000 | 66 | 2010 |
| Upland Cotton | 4,811,000 | 68 | 2010 |
| Sorghum | 1,354,000 | 47 | 2011 |
| Barley | 428,000 | 35 | 2011 |
| Nursery and Floriculture Crops | 89,000 | N/A | 2009 |
1.3. Chemical and Physical Properties
| Common Name | Glyphosate |
|---|---|
| Synonyms | N-(Phosphonomethyl)glycine (acid) |
| Glyphosate isopropylamine salt (IPA salt) | |
| Chemical formula | C3H8NO5P (acid) |
| C3H9N.C3H8NO5P (IPA salt) | |
| Chemical Abstracts Service (CAS) No. | 1071-83-6 (acid) |
| 38641-94-0 (IPA salt) | |
| Molecular weight (g·mol−1) | 169.09 (acid) |
| 227.2 (IPA salt) | |
| Physical description | White crystalline powder |
| Melting point | 200–230 °C |
| Boiling point | No data available |
| Water solubility | 10,000–15,700 mg·L−1 at 25 °C |
| Vapor pressure | 2.59 × 10−5 Pa at 25 °C |
| Octanol/water partition coefficient: log Kow | −4.59 to −1.70 |
| Sorption partition coefficient: Kd | 3–1188; geometric mean (n = 28), 64 |
| Sorption partition coefficient: Koc (L·kg−1) | 9–60,000; geometric mean (n = 28), 2.072 |
| Acid dissociation constants | |
| pKa1 (first phosphonic) | 0.8 |
| pKa2 (carboxylate) | 2.3 |
| pKa3 (second phosphonic) | 6.0 |
| pKa4 (amine) | 11.0 |
1.4. Plant Uptake, Transport and Metabolism
1.5. Mode of Action and Effects in Plants
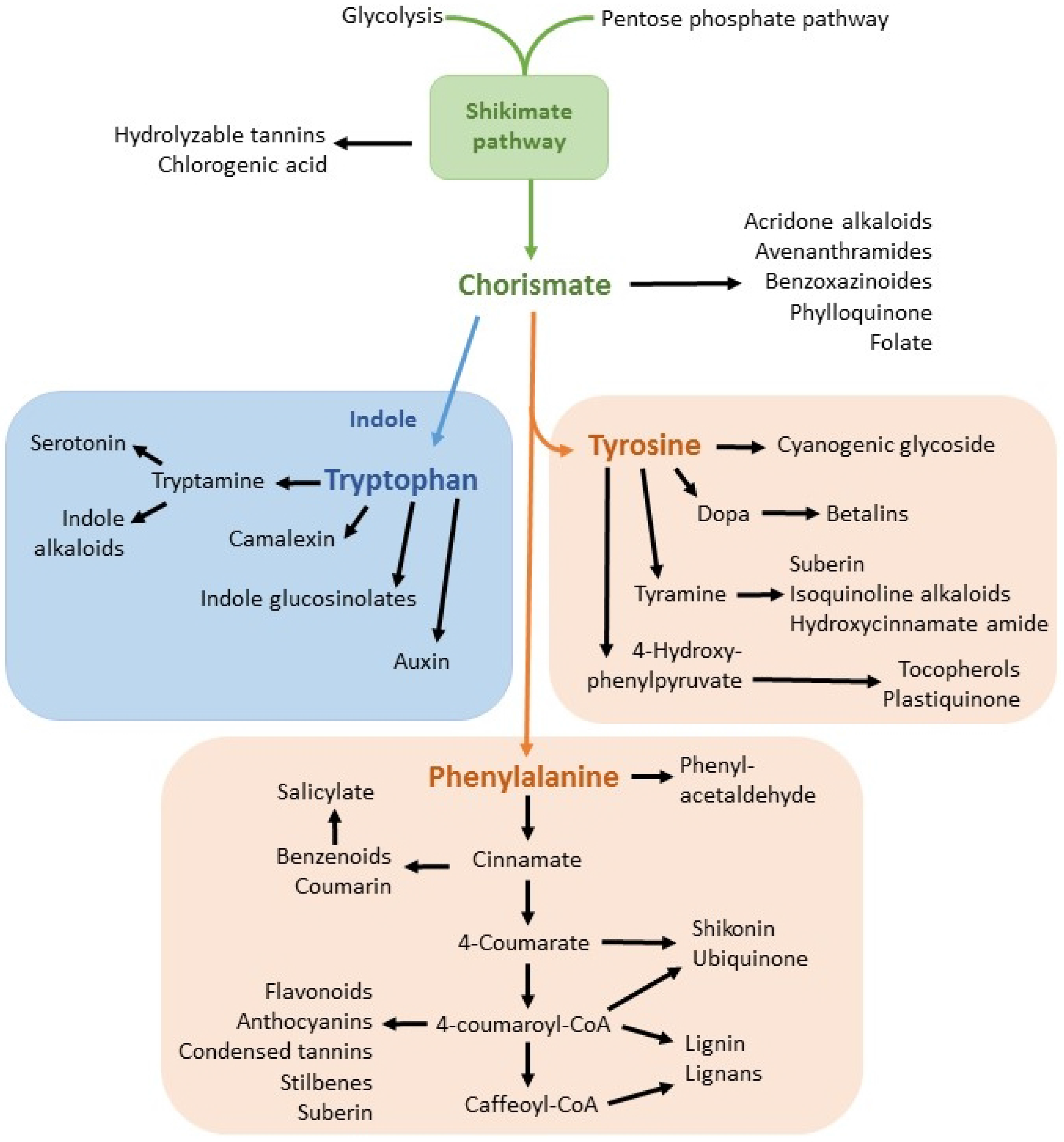
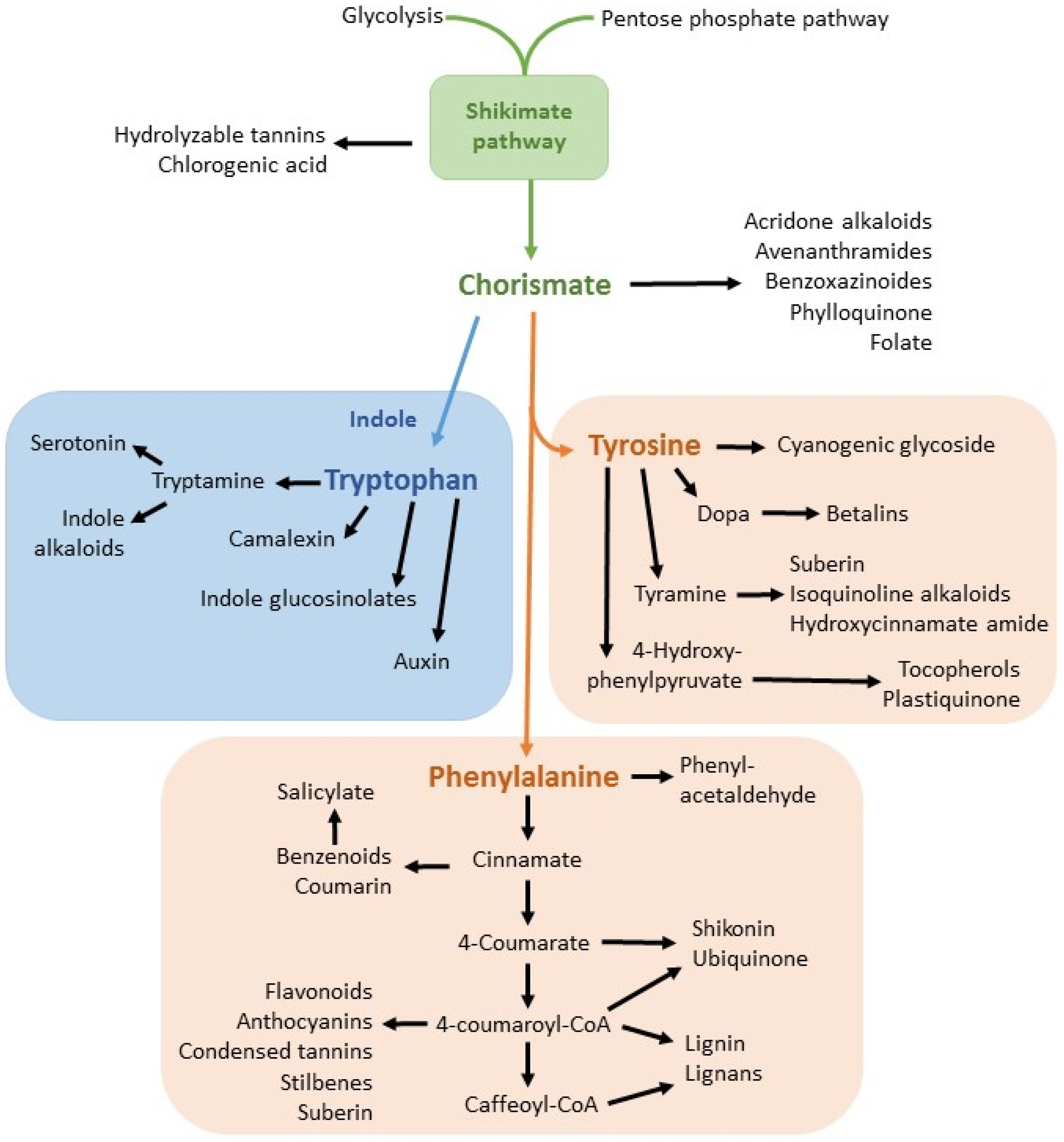
1.6. Resistance to Glyphosate
| Family | Species | Locations | Year(s) Reported |
|---|---|---|---|
| Amaranthaceae | Amaranthus palmeri | AL, AR, AZ, DE, FL, GA, IN, IL, KS, KY, LA, MD, MI, MS, MO, NC, NM, OH, PA, SC, TN, TX, VA, United States | 2005–2014 |
| Amaranthus quitensis | Argentina | 2013 | |
| Amaranthus spinosus | MS, United States | 2012 | |
| Amaranthus tuberculatus | IL, IN, IA, KS, KY, MN, MS, MO, NE, OH, OK, SD, TN, TX, United States | 2005–2012 | |
| Asteraceae/Compositae | Ambrosia artemisiifolia | AL, AR, IN, KS, KY, MN, MS, MO, NE, NJ, NC, ND, OH, PA, SD, United States; ON, Canada | 2004; 2006–2008; 2012–2014 |
| Ambrosia trifida | AR, IN, IA, KS, KY, MN, MS, MO, NE, OH, TN, WI, United States; ON, Canada | 2004–2011 | |
| Bidens pilosa | Mexico | 2014 | |
| Conyza bonariensis | NSW, QLD, SA, Australia; Brazil; Colombia; Greece; Israel; South Africa; Spain; Portugal; CA, United States | 2003–2007; 2009–2011 | |
| Conyza canadensis | AR, CA, DE, IN, IL, IA, KS, KY, MD, MI, MS, MO, NE, NJ, NC, OH, OK, PA, TN, VA, United States; Brazil; China; Czech Republic; Italy; Poland; Spain | 2000–2003; 2005–2007; 2009–2013 | |
| Conyza sumatrensis | Brazil; France; Greece; Spain | 2009–2012 | |
| Parthenium hysterophorus | Colombia | 2004 | |
| Brassicaceae/Cruciferae | Raphanus raphanistrum | WA, Australia | 2010 |
| Chenopodiaceae | Kochia scoparia | AB, SK, Canada; CO, KS, NE, ND, OK, MT, SD, United States | 2007; 2009; 2011–2013 |
| Plantaginaceae | Plantago lanceolata | South Africa | 2003 |
| Poaceae/Gramineae | Chloris elata | Brazil | 2014 |
| Chloris truncata | NSW, Australia | 2010 | |
| Cynodon hirsutus | Argentina | 2008 | |
| Digitaria insularis | Brazil; Paraguay | 2005; 2008 | |
| Echinochloa colona | Argentina; NSW, QLD, WA, Australia; CA, United States | 2007–2010 | |
| Eleusine indica | Argentina; Bolivia; China; Colombia; Costa Rica; Malaysia; MS, TN, United States | 1997; 2006–2007; 2010–2012; 2014 | |
| Leptochloa virgate | Mexico | 2010 | |
| Lolium perenne | Argentina; Brazil; Chile; Japan; Italy; New Zealand; Portugal; Spain; AR, CA, LA, MS, NC, OR, TN, United States | 2001–2012; 2014 | |
| Lolium rigidum | NSW, VIC, SA, WA, Australia; France; Israel; Italy; South Africa; Spain; CA, United States | 1996–1999; 2001; 2003; 2005–2008; 2010; 2013 | |
| Poa annua | CA, MO, TN, United States | 2010–2011; 2013 | |
| Sorghum halepense | Argentina; AR, LA, MS, United States | 2005; 2007–2008; 2010 | |
| Urochloa panicoides | NSW, Australia | 2008 | |
| Rubiaceae | Hedyotis verticillata | Malaysia | 2014 |
2. Environmental Fate
2.1. Soil Interactions
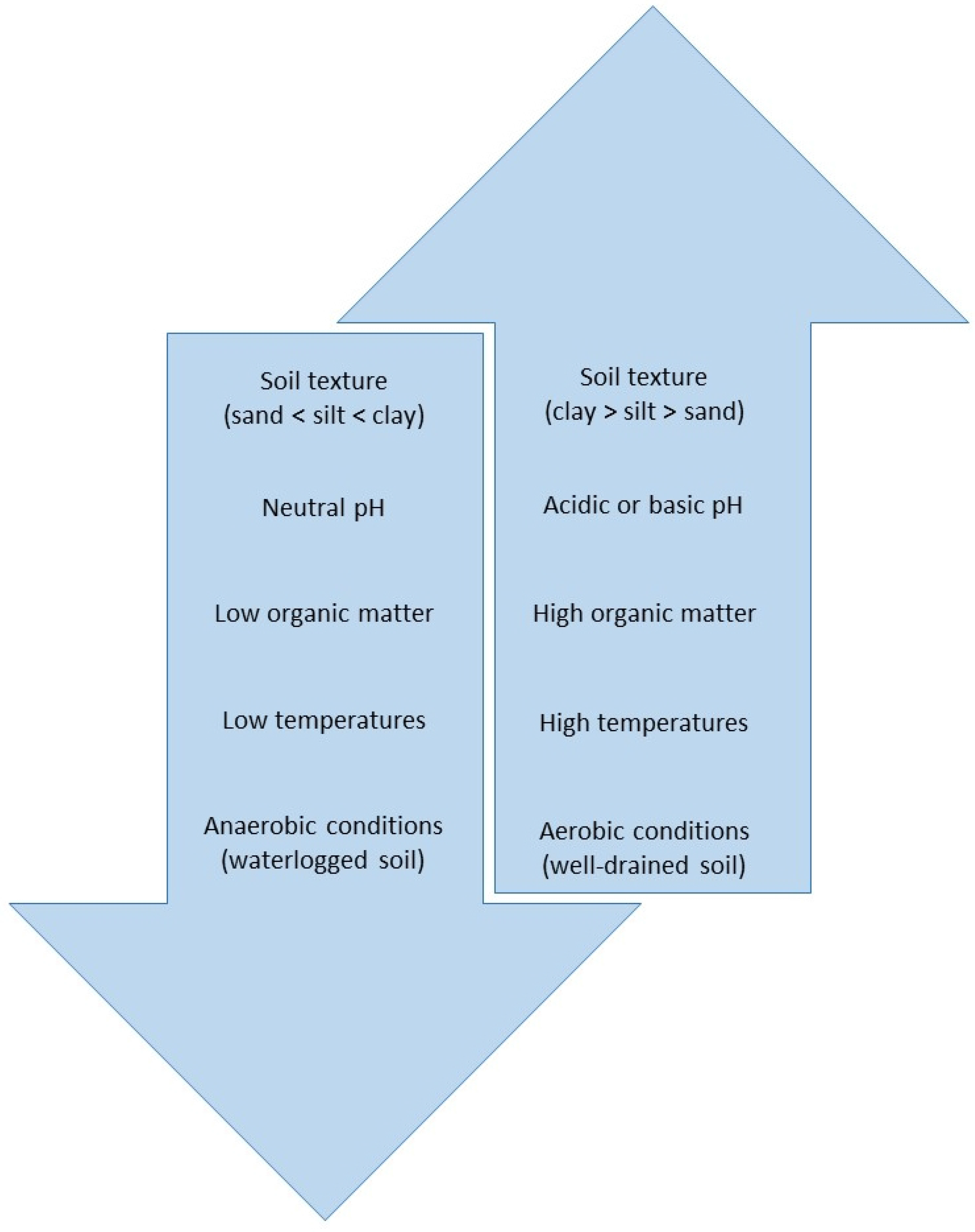
| Reference | Location | DT50 (Days) |
|---|---|---|
| Mestdagh, 1979 [70] | France | 5–197.3 |
| Mestdagh, 1979 [70] | Sweden | 1.2–24.3 |
| Danhaus, 1984 [71] | USA | 27.3–55.5 |
| Heinonen-Tanski et al., 1985 [72] | Finland | <58 |
| Ragab et al., 1985 [73] | Canada | <10 |
| Oppenhuizen 1993 [74] | USA | 1.7–141.9 |
| Oppenhuizen and Goure, 1993 [75] | Canada | 6–21 |
| Thompson et al., 2000 [76] | Canada | 10–12 |
| Veiga et al., 2001 [77] | Spain | <30 |
| Simonsen et al., 2008 [78] | Denmark | 9 |
2.2. Occurrence in Water
2.3. Implications for Non-Target Vegetation
| Species | Endpoint | Summary of Effects | Reference |
|---|---|---|---|
| Beet | Betacyanin efflux | Betacyanin efflux increased with increasing glyphosate concentration and time, demonstrating increased cell membrane permeability of root tissue | Fletcher et al., 1980 [15] |
| (Beta vulgaris) | |||
| Barley | Changes in dry weight | 23% reduction in shoot dry weight | Penn and Lynch, 1982 [16] |
| (Hordeum vulgare) | |||
| Cotton | Changes in fresh weight; lateral root development | 50% reduction in fresh weight of cotyledons, hypocotyls and roots; inhibition of lateral root development | Pline et al., 2002 [17] |
| (Gossypium hirsutum) | |||
| Maize | Changes in fresh weight; visual symptoms | Growth reduction of up to 44% of fresh weights following a logistic response curve; hormesis effect noted for exposures of less than 1 µg·L−1; wilting and chlorosis for exposures greater than 1 µg·L−1 | Wagner et al., 2003 [13] |
| (Zea mays) | |||
| Maize | Changes in fresh weight | Growth reduction of 50% of fresh weights for exposures of 30 mg·L−1 | Alister et al., 2005 [18] |
| (Zea mays) | |||
| Rapeseed | Changes in dry weight; visual symptoms | Growth reduction of 83% of dry weights for roots and 43% reduction for shoots; leaf chlorosis and necrosis for exposures of 20 µM·L−1 or greater | Petersen et al., 2007 [19] |
| (Brassica napus) | |||
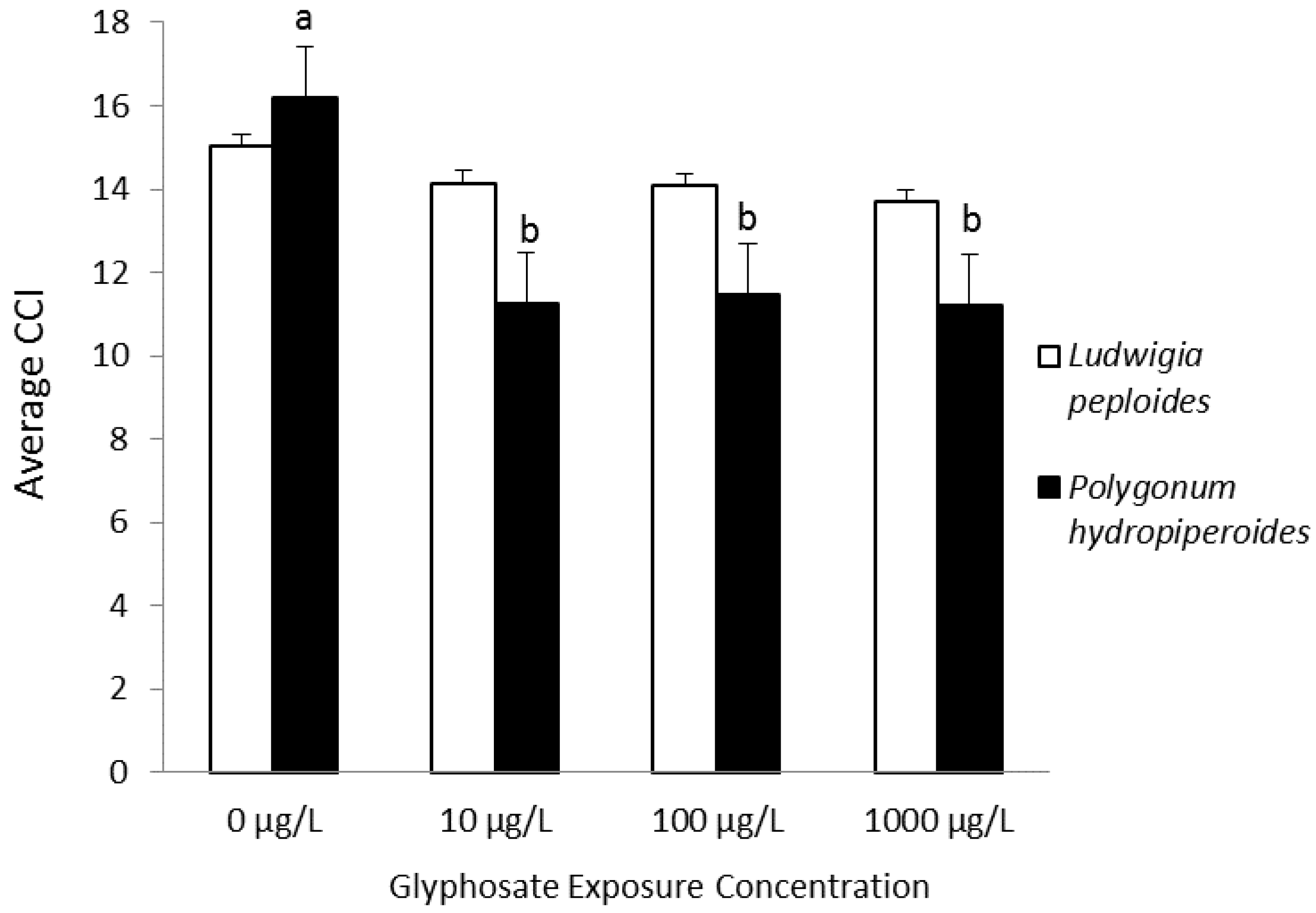
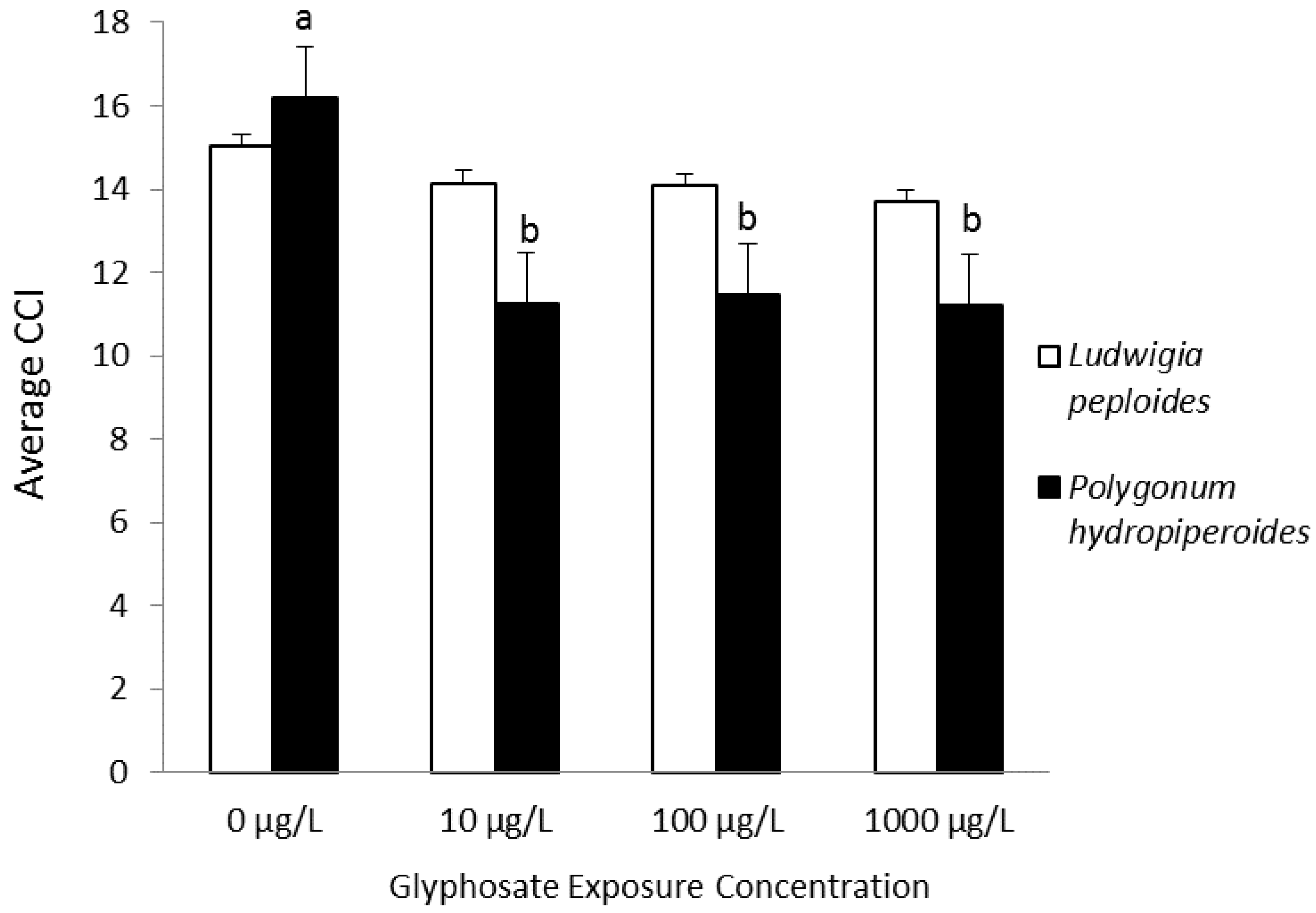
| Smartweed | Changes in leaf chlorophyll content and dry weight; survival | Dose-dependent reductions in leaf chlorophyll content in P. hydropiperoides and P. hemitomon; no differences in dry weight for either species; survival at 10 µg·L−1 for P. hydropiperoides and mortality at higher concentrations; survival by P. hemitomon except at 10,000 µg·L−1 | Saunders et al., 2013 [93] |
| (Polygonum hydropiperoides) | |||
| Maidencane | |||
| (Panicum hemitomon) | |||
| Smartweed | Changes in leaf chlorophyll content, chlorophyll fluorescence parameters, and dry weight | Reduction in chlorophyll content for treated P. hydropiperoides; species-specific reductions in chlorophyll fluorescence parameters; no differences in dry weight | Saunders and Pezeshki, 2014 [94] |
| (Polygonum hydropiperoides) | |||
| Creeping water primrose | |||
| (Ludwigia peploides) | |||
| Creeping water primrose | Changes in morphology and dry weight | Hormesis effect depending on root density of connected ramets | Saunders and Pezeshki, 2015 [95] |
| (Ludwigia peploides) |
3. Conclusions
Acknowledgments
Conflicts of Interest
References
- Franz, J.E.; Mao, M.K.; Sikorski, J.A. Glyphosate: A Unique Global Herbicide; American Chemical Society: Washington, DC, USA, 1997. [Google Scholar]
- Duke, S.O.; Powles, S.B. Glyphosate: A once-in-a-century herbicide. Pest Manag. Sci. 2008, 64, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Dill, G.M.; CaJacob, C.A.; Padgette, S.R. Glyphosate-resistant crops: Adoption, use and future considerations. Pest Manag. Sci. 2008, 64, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Giesy, J.P.; Dobson, S.; Solomon, K.R. Ecotoxicological risk assessment for Roundup® herbicide. Rev. Environ. Contam. Toxicol. 2000, 167, 35–120. [Google Scholar]
- 2012 Agricultural Chemical Use Survey: Soybeans. NASS Highlights. Available online: http://www.nass.usda.gov/Surveys/Guide_to_NASS_Surveys/Chemical_Use/2012_Soybeans_Highlights/ChemUseHighlights-Soybeans-2012.pdf (accessed on 20 November 2014).
- Agricultural Chemical Use Field Crops 2011: Barley and Sorghum. Available online: http://www.nass.usda.gov/Surveys/Guide_to_NASS_Surveys/Chemical_Use/BarleySorghumChemicalUseFactSheet.pdf (accessed on 20 November 2014).
- Agricultural Chemical Use: Corn, Upland Cotton and Fall Potatoes 2010. Available online: http://www.nass.usda.gov/Surveys/Guide_to_NASS_Surveys/Chemical_Use/FieldCropChemicalUseFactSheet06.09.11.pdf (accessed on 20 November 2014).
- Agricultural Chemical Use: Nursery and Floriculture Crops 2009. Available online: http://www.nass.usda.gov/Surveys/Guide_to_NASS_Surveys/Chemical_Use/NurseryFloricultureChemicalUseFactSheet.pdf (accessed on 20 November 2014).
- Dikshith, T.S.; Diwan, P.V. Industrial Guide to Chemical and Drug Safety; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2003. [Google Scholar]
- Bromilow, R.H.; Chamberlain, K. The herbicide glyphosate and related molecules: Physiochemical and structural factors determining their mobility in phloem. Pest Manag. Sci. 2000, 56, 368–373. [Google Scholar] [CrossRef]
- Dewey, S.A. Manipulation of Assimilate Transport Patterns as a Method of Studying Glyphosate Translocation in Tall Morningglory [Ipomoea purpurea (L.) Roth]. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, June 1981. [Google Scholar]
- Duke, S.O. Glyphosate. In Herbicides: Chemistry, Degradation, and Mode of Action; Kearney, P.C., Kaufman, D.D., Eds.; Marcel Dekker: New York, NY, USA, 1988; pp. 1–70. [Google Scholar]
- Wagner, R.; Kogan, M.; Parada, A.M. Phytotoxic activity of root absorbed glyphosate in corn seedlings (Zea mays L.). Weed Biol. Manag. 2003, 3, 228–232. [Google Scholar] [CrossRef]
- Clua, A.; Conti, M.; Beltrano, J. The effects of glyphosate on the growth of Birdsfoot Trefoil (Lotus corniculatus) and its interaction with different phosphorus contents in soil. J. Agric. Sci. 2012, 4, 208–218. [Google Scholar]
- Fletcher, R.A.; Hildebrand, P.; Akey, W. Effect of glyphosate on membrane permeability in red beet (Beta vulgaris) root tissue. Weed Sci. 1980, 28, 671–673. [Google Scholar]
- Penn, D.J.; Lynch, J.M. Toxicity of glyphosate applied to roots of barley seedlings. New Phytol. 1982, 90, 51–55. [Google Scholar] [CrossRef]
- Pline, W.A.; Wilcut, J.W.; Edmisten, K.L.; Wells, R. Physiological and morphological response of glyphosate-resistant and non-glyphosate-resistant cotton seedlings to root-absorbed glyphosate. Pestic. Biochem. Phys. 2002, 73, 48–58. [Google Scholar] [CrossRef]
- Alister, C.; Kogan, M.; Pino, I. Differential phytotoxicity of glyphosate in maize seedlings following applications to roots or shoot. Weed Res. 2005, 45, 27–32. [Google Scholar] [CrossRef]
- Petersen, I.L.; Hansen, H.C.; Ravn, H.W.; Sørensen, J.C.; Sørensen, H. Metabolic effects in rapeseed (Brassica napus L.) seedlings after root exposure to glyphosate. Pestic. Biochem. Phys. 2007, 89, 220–229. [Google Scholar] [CrossRef]
- Causal Analysis/Diagnosis Decision Information System (CADDIS) Volume 2: Sources, Stressors & Responses: Herbicides: Introduction. Available online: http://www.epa.gov/caddis/ssr_herb_int.html (accessed on 22 November 2014).
- Steinrücken, H.C.; Amrhein, N. The herbicide glyphosate is a potent inhibitor of 5-enolpyruvylshikimic acid-3-phosphate synthase. Biochem. Biophys. Res. Commun. 1980, 94, 1207–1212. [Google Scholar] [CrossRef]
- Maeda, H.; Dudareva, N. The shikimate pathway and aromatic amino acid biosynthesis in plants. Annu. Rev. Plant Biol. 2012, 63, 73–105. [Google Scholar] [CrossRef] [PubMed]
- Schmid, J.; Amrhein, N. The shikimate pathway. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B.H., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 147–169. [Google Scholar]
- Mesnage, R.; Defarge, N.; Spiroux de Vendômois, J.; Séralini, G.E. Potential toxic effects of glyphosate and its commercial formulations below regulatory limits. Food Chem. Toxicol. 2015, 84, 133–153. [Google Scholar] [CrossRef] [PubMed]
- Weaver, L.M.; Herrmann, K.M. Dynamics of the shikimic pathway in plants. Trends Plant Sci. 1997, 9, 346–351. [Google Scholar] [CrossRef]
- Gruys, K.J.; Sikorski, J.A. Inhibitors of tryptophan, phenylalanine, and tyrosine biosynthesis as herbicides. In Plant Amino Acids: Biochemistry and Biotechnology; Singh, B.H., Ed.; Marcel Dekker: New York, NY, USA, 1999; pp. 357–384. [Google Scholar]
- Sprankle, P.; Meggit, W.F.; Penner, D. Absorption, action, and translocation of glyphosate. Weed Sci. 1975, 23, 235–240. [Google Scholar]
- Haderlie, L.C.; Slife, F.W.; Butler, H.S. 14C-Glyphosate absorption and translocation in germinating maize (Zea mays) and soybean (Glycine max) seeds and in soybean plants. Weed Res. 1978, 18, 269–273. [Google Scholar] [CrossRef]
- Gougler, J.A.; Geiger, D.R. Uptake and distribution of N-phosphonomethylglycine in sugar beet plants. Plant Physiol. 1981, 68, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Sprankle, P.; Meggit, W.F.; Penner, D. Absorption, mobility, and microbial degradation of glyphosate in the soil. Weed Sci. 1975, 23, 229–234. [Google Scholar]
- Geiger, D.R.; Kapitan, S.W.; Tucci, M.A. Glyphosate inhibits photosynthesis and allocation of carbon to starch in sugar beet leaves. Plant Physiol. 1986, 82, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Madsen, K.H.; Heitholt, J.J.; Duke, S.O.; Smeda, R.J.; Streibig, J.C. Photosynthetic parameters in glyphosate-treated sugarbeet (Beta vulgaris L.). Weed Res. 1995, 35, 81–88. [Google Scholar] [CrossRef]
- Olesen, C.F.; Cedergreen, N. Glyphosate uncouples gas exchange and chlorophyll fluorescence. Pest Manag. Sci. 2010, 66, 536–542. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Silva, E.N.; Shen, Z.; Jiang, B.; Lu, H. Effects of glyphosate on photosynthesis, chlorophyll fluorescence and physiochemical properties of congograss (Imperata cylindrical L.). Plant Omics 2012, 5, 177–183. [Google Scholar]
- Silva, F.; Costa, A.; Pereira Alves, R.; Megguer, C. Chlorophyll fluorescence as an indicator of cellular damage by glyphosate herbicide in Raphanus sativus L. plants. Am. J. Plant Sci. 2014, 5, 2509–2519. [Google Scholar] [CrossRef]
- Kitchen, L.M.; Witt, W.W.; Rieck, C.E. Inhibition of chlorophyll accumulation by glyphosate. Weed Sci. 1981, 29, 513–516. [Google Scholar]
- Reddy, K.N.; Hoagland, R.E.; Zablotowicz, R.M. Effect of glyphosate on growth, chlorophyll, and nodulation in glyphosate-resistant and susceptible soybean (Glycine max) varieties. J. New Seeds 2000, 2, 37–52. [Google Scholar] [CrossRef]
- Velini, E.D.; Alves, E.; Godoy, M.C.; Meschede, D.K.; Souza, R.T.; Duke, S.O. Glyphosate applied at low doses can stimulate plant growth. Pest Manag. Sci. 2008, 64, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Cedergreen, N. Is the growth stimulation by low doses of glyphosate sustained over time? Environ. Pollut. 2008, 156, 1099–1104. [Google Scholar] [CrossRef] [PubMed]
- Belz, R.G.; Cedergreen, N.; Duke, S.O. Herbicide hormesis—Can it be useful in crop production? Weed Res. 2011, 51, 321–332. [Google Scholar] [CrossRef]
- Belz, R.G.; Duke, S.O. Herbicides and plant hormesis. Pest Manag. Sci. 2014, 70, 698–707. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, L.B.; Alves, P.L.C.A.; Duke, S.O. Hormesis with glyphosate depends on coffee growth stage. Ann. Braz. Acad. Sci. 2013, 85, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Shaner, D.L.; Lindenmeyer, R.B.; Ostlie, M.H. What have the mechanisms of resistance to glyphosate taught us? Pest Manag. Sci. 2012, 68, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Pollegioni, L.; Schonbrunn, E.; Siehl, D. Molecular basis of glyphosate resistance: Different approaches through protein engineering. FEBS J. 2011, 278, 2753–2766. [Google Scholar] [CrossRef] [PubMed]
- Heap, I. The International Survey of Herbicide Resistant Weeds. Available online: http://www.weedscience.org (accessed on 5 December 2014).
- Hance, R.J. Adsorption of glyphosate by soils. Pestic. Sci. 1976, 7, 363–366. [Google Scholar] [CrossRef]
- Roy, D.N.; Konar, S.K.; Banerjee, S.; Charles, D.A.; Thompson, D.G.; Prasad, R. Persistence, movement and degradation of glyphosate in selected Canadian boreal forest soils. J. Agric. Food Chem. 1989, 37, 437–440. [Google Scholar] [CrossRef]
- Piccolo, A.; Celano, G. Hydrogen bonding interactions between the herbicide glyphosate and water-soluble humic substances. Environ. Toxicol. Chem. 1994, 13, 1737–1741. [Google Scholar] [CrossRef]
- Sheals, J.; Sjӧberg, S.; Persson, P. Adsorption of glyphosate on goethite: Molecular characteristics of surface complexes. Environ. Sci. Technol. 2002, 36, 3090–3095. [Google Scholar] [CrossRef] [PubMed]
- Gimsing, A.L.; Borggaard, O.K. Competitive adsorption and desorption of glyphosate and phosphate on clay silicates and oxides. Clay Miner. 2002, 37, 509–515. [Google Scholar] [CrossRef]
- Gimsing, A.L.; Borggaard, O.K.; Bang, M. Influence of soil composition on adsorption of glyphosate and phosphate by contrasting Danish surface soils. Eur. J. Soil Sci. 2004, 55, 183–191. [Google Scholar] [CrossRef]
- Vereecken, H. Mobility and leaching of glyphosate: A review. Pest Manag. Sci. 2005, 61, 1139–1151. [Google Scholar] [CrossRef] [PubMed]
- Gimsing, A.L.; Szilas, C.; Borggaard, O.K. Sorption of glyphosate and phosphate by variable-charge tropical soils from Tanzania. Geoderma 2007, 138, 127–132. [Google Scholar] [CrossRef]
- Borggaard, O.K.; Gimsing, A.L. Fate of glyphosate in soil and the possibility of leaching to ground and surface waters: A review. Pest Manag. Sci. 2008, 64, 441–456. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.J.; Moye, H.A. Extraction of glyphosate herbicide from soil and clay minerals and determination of residues in soil. J. Agric. Food Chem. 1988, 36, 486–491. [Google Scholar] [CrossRef]
- Dion, H.M.; Harsh, J.B.; Hill, H.H., Jr. Competitive sorption between glyphosate and inorganic phosphate on clay minerals. J. Radioanal. Nucl. Chem. 2001, 249, 385–390. [Google Scholar]
- Madhun, Y.A.; Young, J.L.; Freed, V.H. Binding of herbicides by water-soluble organic materials from soil. J. Environ. Qual. 1986, 15, 64–68. [Google Scholar] [CrossRef]
- Piccolo, A.; Gatta, L.; Campanella, L. Interactions of glyphosate herbicide with a humic acid and its iron complex. Ann. Chim. 1995, 85, 31–40. [Google Scholar]
- Noruma, N.S.; Hilton, H.W. The adsorption and degradation of glyphosate in five Hawaiian sugarcane soils. Weed Res. 1977, 17, 113–121. [Google Scholar]
- Piccolo, A.; Celano, G.; Conte, P. Adsorption of glyphosate by humic substances. J. Agric. Food Chem. 1996, 44, 2442–2446. [Google Scholar] [CrossRef]
- Piccolo, A.; Celano, G.; Arienzo, M.; Mirabella, A. Adsorption and desorption of glyphosate in some European soils. J. Environ. Sci. Health B 1996, 29, 1105–1115. [Google Scholar] [CrossRef]
- Gerritse, R.G.; Beltran, J.; Hernandez, F. Adsorption of atrazine, simazine, and glyphosate in soil of Gnangara Mound, Western Australia. Austr. J. Soil Res. 1996, 34, 599–607. [Google Scholar] [CrossRef]
- Morillo, E.; Undabeytia, T.; Maqueda, C.; Ramos, A. Glyphosate adsorption on soils of different characteristics. Influence of copper addition. Chemosphere 1999, 40, 103–107. [Google Scholar] [CrossRef]
- De Jonge, H.; de Jonge, L.W.; Jacobson, O.H.; Yamaguchi, T.; Moldrup, P. Glyphosate sorption in soils of different pH and phosphorus content. Soil Sci. 2001, 166, 230–238. [Google Scholar] [CrossRef]
- Reddy, K.N.; Rimando, A.M.; Duke, S.O. Aminomethylphosphonic acid, a metabolite of glyphosate, causes injury in glyphosate-treated, glyphosate-resistant soybean. J. Agric. Food Chem. 2004, 52, 5139–5143. [Google Scholar] [CrossRef] [PubMed]
- Gomes, M.P.; Smedbol, E.; Chalifour, A.; Hénault-Ethier, L.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid: An overview. J. Exp. Bot. 2014, 65, 4691–4703. [Google Scholar] [CrossRef] [PubMed]
- Moshier, L.J.; Penner, D. Factors influencing microbial degradation of 14C-glyphosate to 14CO2 in soil. Weed Sci. 1978, 26, 686–691. [Google Scholar]
- Carlisle, S.M.; Trevors, J.T. Glyphosate in the environment. Water Air Soil Pollut. 1988, 39, 409–420. [Google Scholar]
- Heinonen-Tanski, H. The effect of temperature and liming on the degradation of glyphosate in two artic forest soils. Soil Biol. Biochem. 1989, 21, 313–317. [Google Scholar] [CrossRef]
- Mestdagh, P. Field Soil Dissipation Studies of Roundup Conducted in Sweden and France; Report MLL-30.033; Monsanto Company: St. Louis, MO, USA, Unpublished work; 1979. [Google Scholar]
- Danhaus, R.G. Dissipation of Glyphosate in USA Field Soils Following Multiple Applications of Roundup Herbicide; Report MSL-3352; Monsanto Company: St. Louis, MO, USA, Unpublished work; 1984. [Google Scholar]
- Heinonen-Tanski, H.; Rosenburg, C.; Siltanen, H.; Kilpi, S.; Simojoki, P. The effect of annual use of pesticides on soil microorganisms, pesticide residues in the soil and barley yields. Pestic. Sci. 1985, 16, 341–348. [Google Scholar] [CrossRef]
- Ragab, M.T.H.; Abdel-Kader, M.K.H.; Stiles, D.A. Fate of glyphosate in sandy loam soil and analysis for residues in field-grown crops. Proc. N. S. Inst. Sci. 1985, 35, 67–70. [Google Scholar]
- Oppenhuizen, M.E. The Terrestrial Field Dissipation of Glyphosate: Final Report; Report MSL-9238; Monsanto Company: St. Louis, MO, USA, Unpublished work; 1993. [Google Scholar]
- Oppenhuizen, M.E.; Goure, W.F. The Terrestrial Field Dissipation of Glyphosate in Canadian Soils; Report MSL-12605; Monsanto Company: St. Louis, MO, USA, Unpublished work; 1993. [Google Scholar]
- Thompson, D.G.; Pitt, D.G.; Buscarini, T.M.; Staznik, B.; Thomas, D.R. Comparative fate of glyphosate and triclopyr herbicides in the forest floor and mineral soul of an Acadian forest regeneration site. Can. J. For. Res. 2000, 30, 1808–1816. [Google Scholar] [CrossRef]
- Veiga, F.; Zapata, J.M.; Fernandez Marcos, M.L.; Alvarez, E. Dynamics of glyphosate and aminomethylphosphonic acid in forest soil in Galicia, north-west Spain. Sci. Total Environ. 2001, 271, 135–144. [Google Scholar] [CrossRef]
- Simonsen, L.; Fomsgaard, I.S.; Svensmark, B.; Spliid, N.H. Fate and availability of glyphosate and AMPA in agricultural soil. J. Environ. Sci. Health B 2008, 43, 365–375. [Google Scholar] [CrossRef] [PubMed]
- Technical Factsheet on Glyphosate. Available online: http://www.epa.gov/safewater/pdfs/factsheets/soc/tech/glyphosa.pdf (accessed on 4 December 2014).
- US Environmental Protection Agency. Occurrence estimation methodology and occurrence findings report for the six year review of existing national primary drinking water regulations. Office of Water EPA-815-R-03-006; US Environmental Protection Agency: Washington, DC, USA, Unpublished work; 2003. [Google Scholar]
- Edwards, W.M.; Triplett, G.B.; Kramer, R.M. A watershed study of glyphosate transport in runoff. J. Environ. Qual. 1980, 9, 661–665. [Google Scholar] [CrossRef]
- Kjaer, J.; Ullum, M.; Olsen, P.; Helweg, A.; Mogensen, B.B.; Plauborg, F.; Grant, R.; Fomsgaard, I.S.; Brüsch, W. The Danish Pesticide Leaching Assessment Programme: Monitoring Results May 1999–June 2002. Available online: http://pesticidvarsling.dk/xpdf/vap-results-99-10.pdf (accessed on 4 December 2014).
- Shipitalo, M.J.; Owens, L.B. Comparative losses of glyphosate and selected residual herbicides in surface runoff from conservation-tilled watersheds planted with corn or soybean. J. Environ. Qual. 2011, 40, 1281–1289. [Google Scholar] [CrossRef] [PubMed]
- Battaglin, W.A.; Kolpin, D.W.; Scribner, E.A.; Kuivila, K.M.; Sandstrom, M.W. Glyphosate, other herbicides, and transformation products in Midwestern streams. J. Am. Water Resour. Assoc. 2005, 41, 323–332. [Google Scholar] [CrossRef]
- Coupe, R.H.; Kalkhoff, S.J.; Capel, P.D.; Gregoire, C. Fate and transport of glyphosate and aminomethylphosphonic acid in surface waters of agricultural basins. Pest Manag. Sci. 2011, 68, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Breeze, V.; Thomas, G.; Butler, R. Use of a model and toxicity data to predict the risks to some wild plant species from drift of four herbicides. Ann. Appl. Bot. 1992, 121, 669–677. [Google Scholar] [CrossRef]
- Marrs, R.H.; Frost, A.J.; Plant, R.A.; Lunnis, P. Determination of buffer zones to protect seedlings of non-target plants from the effects of glyphosate spray drift. Agric. Ecosyst. Environ. 1993, 45, 283–293. [Google Scholar] [CrossRef]
- Kleijn, D.; Snoeijing, I.J. Field boundary vegetation and the effects of agrochemical drift: Botanical change caused by low levels of herbicide and fertilizer. J. Appl. Ecol. 1997, 34, 1413–1425. [Google Scholar] [CrossRef]
- De Snoo, G.R.; van der Poll, R.J. Effect of herbicide drift on adjacent boundary vegetation. Agric. Ecosyst. Environ. 1999, 73, 1–6. [Google Scholar] [CrossRef]
- Dixon, F.L.; Clay, D.V.; Willoughby, I. Susceptibility of woodland plants to herbicide drift. Q. J. For. 2002, 96, 32–36. [Google Scholar]
- Hewitt, A.J.; Solomon, K.R.; Marshall, E.J.P. Spray droplet size, drift potential, and risks to nontarget organisms from aerially applied glyphosate for coca control in Colombia. J. Toxicol. Environ. Health A 2009, 72, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Thomson, S.J.; Ortiz, B.V.; Reddy, K.N.; Ding, W.; Zablotowicz, R.M.; Bright, J.R. Airborne remote sensing assessment of the damage to cotton caused by spray drift from aerially applied glyphosate through spray deposition measurements. Biosyst. Eng. 2010, 107, 212–220. [Google Scholar] [CrossRef]
- Saunders, L.E.; Koontz, M.B.; Pezeshki, R. Root-zone glyphosate exposure adversely affects two ditch species. Biology 2013, 2, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Saunders, L.E.; Pezeshki, R. Sublethal effects of environmentally relevant run-off concentrations of glyphosate in the root zone of Ludwigia peploides (creeping water primrose) and Polygonum hydropiperoides (smartweed). Weed Biol. Manag. 2014, 14, 242–250. [Google Scholar] [CrossRef]
- Saunders, L.E.; Pezeshki, S. Morphological differences in response to physiological integration and spatial heterogeneity of root zone glyphosate exposure in connected ramets of Lugwigia peploides (creeping water primrose). Water Air Soil Pollut. 2015, 226, 171. [Google Scholar] [CrossRef]
- Moore, M.T.; Bennett, E.R.; Cooper, C.M.; Smith, S., Jr.; Shields, F.D., Jr.; Milam, C.D.; Farris, J.L. Transport and fate of atrazine and lambda-cyhalothrin in an agricultural drainage ditch in the Mississippi Delta, USA. Agric. Ecosyst. Environ. 2001, 87, 309–314. [Google Scholar] [CrossRef]
- Cooper, C.M.; Moore, M.T.; Bennett, E.R.; Smith, S., Jr.; Farris, J.L. Alternative environmental benefits of agricultural drainage ditches. Verh. Int. Ver. Limnol. 2002, 28, 1678–1682. [Google Scholar]
- Cooper, C.M.; Moore, M.T.; Bennett, E.R.; Smith, S., Jr.; Farris, J.L.; Milam, C.D.; Shields, F.D., Jr. Innovative uses of vegetated drainage ditches for reducing agricultural runoff. Water Sci. Technol. 2004, 49, 117–123. [Google Scholar] [PubMed]
- Bouldin, J.L.; Farris, J.L.; Moore, M.T.; Smith, S., Jr.; Stephens, W.W.; Cooper, C.M. Evaluated fate and effects of atrazine and lambda-cyhalothrin in vegetated and unvegetated microcosms. Environ. Toxicol. 2005, 20, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Bouldin, J.L.; Farris, J.L.; Moore, M.T.; Smith, S., Jr.; Cooper, C.M. Hydroponic uptake of atrazine and lambda-cyhalothrin in Juncus effusus and Ludwigia peploides. Chemosphere 2006, 65, 1049–1057. [Google Scholar] [CrossRef] [PubMed]
- Krӧger, R.; Moore, M.T.; Farris, J.L.; Gopalan, M. Evidence for the use of low-grade weirs in drainage ditches to improve nutrient reductions from agriculture. Water Air Soil Pollut. 2011, 221, 223–234. [Google Scholar] [CrossRef]
- Stehle, S.; Elsaesser, D.; Gregoire, C.; Imfeld, G.; Niehaus, E.; Passeport, E.; Payraudea, S.; Schӓfer, R.B.; Tournebize, J.; Schulz, R. Pesticide risk mitigation by vegetated treatment systems: A meta-analysis. J. Environ. Qual. 2011, 40, 1068–1080. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saunders, L.E.; Pezeshki, R. Glyphosate in Runoff Waters and in the Root-Zone: A Review. Toxics 2015, 3, 462-480. https://doi.org/10.3390/toxics3040462
Saunders LE, Pezeshki R. Glyphosate in Runoff Waters and in the Root-Zone: A Review. Toxics. 2015; 3(4):462-480. https://doi.org/10.3390/toxics3040462
Chicago/Turabian StyleSaunders, Lyndsay E., and Reza Pezeshki. 2015. "Glyphosate in Runoff Waters and in the Root-Zone: A Review" Toxics 3, no. 4: 462-480. https://doi.org/10.3390/toxics3040462
APA StyleSaunders, L. E., & Pezeshki, R. (2015). Glyphosate in Runoff Waters and in the Root-Zone: A Review. Toxics, 3(4), 462-480. https://doi.org/10.3390/toxics3040462