
Exposure–Response Relationship of Toxic Metal(loid)s in Mammals: Their Bioinorganic Chemistry in Blood Is an Intrinsic Component of the Selectivity Filters That Mediate Organ Availability



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Assessment of Human Exposure to TMs
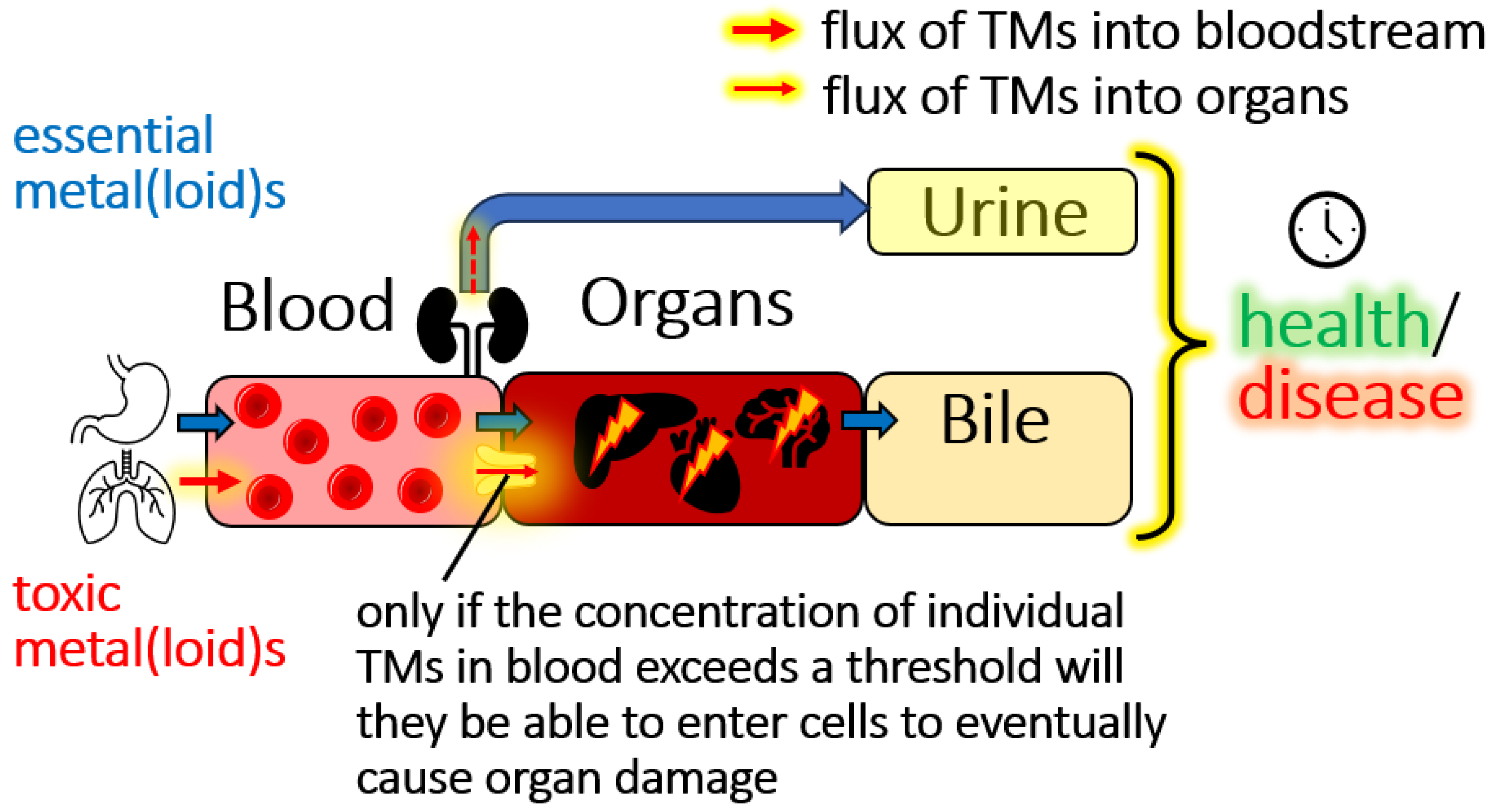
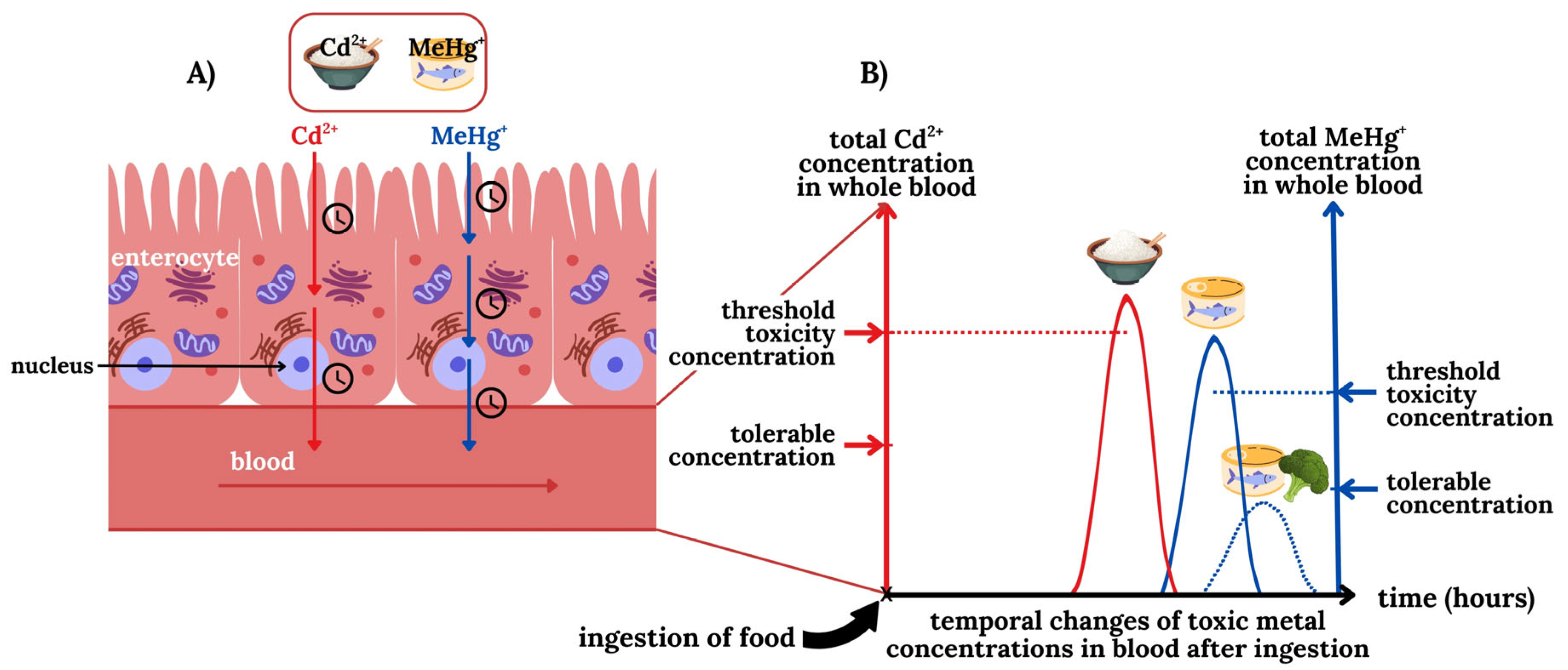
2.1. Can We Distinguish Between Tolerable and Threshold Toxicity Blood Concentrations of TMs?
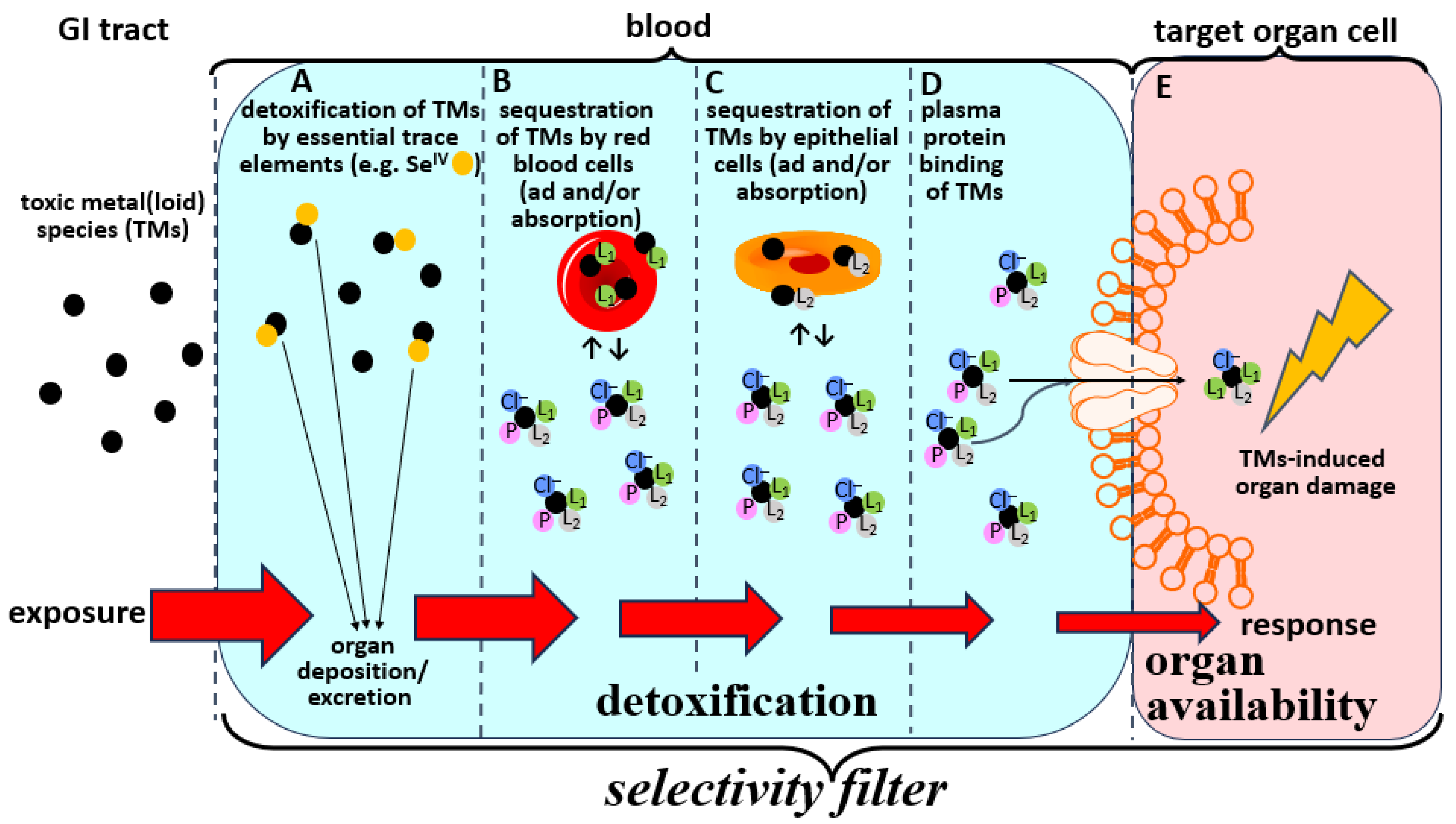
2.2. Bioinorganic Processes of TMs in the Blood-Organ Interface Represent Selectivity Filters
2.3. Structural Characterization of Organ Available TMs Species in Blood Plasma
2.4. Involvement of SMW Species in Plasma in Mediating the Organ Availability of TMs
3. Modulation of the Bioinorganic Chemistry of TMs in Blood to Mitigate Toxic Effects
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hou, D.; O’Connor, D.; Igalavithana, A.D.; Alessi, D.S.; Luo, J.; Tsang, D.C.W.; Sparks, D.L.; Yamauchi, Y.; Rinklebe, J.; Ok, Y.S. Metal contamination and bioremediation of agricultural soils for food safety and sustainability. Nat. Rev. Earth Environ. 2020, 1, 366–381. [Google Scholar] [CrossRef]
- Briffa, J.; Sinagra, E.; Blundell, R. Heavy metal pollution in the environment and their toxicological effects on humans. Heliyon 2020, 6, e044691. [Google Scholar] [CrossRef]
- Hou, D.; Jia, X.; Wang, L.; McGrath, S.P.; Zhu, Y.-G.; Hu, Q.; Zhao, F.J.; Bank, M.S.; O’Connor, D.; Nriagu, J. Global soil pollution by toxic metals threatens agriculture and human health. Science 2025, 388, 316–321. [Google Scholar] [CrossRef]
- Gailer, J. Toward a mechanim-driven integrated framework to link human exposure to multiple toxic metal(loid) species with environmental diseases. Int. J. Mol. Sci. 2024, 25, 3393. [Google Scholar] [CrossRef]
- Gustin, K.; Tofail, F.; Vahter, M.; Kippler, M. Cadmium exposure and cognitive abilities and behavior at 10 years of age: A prospective cohort study. Environ. Int. 2018, 113, 259–268. [Google Scholar] [CrossRef]
- Jeong, K.S.; Park, H.; Ha, E.H.; Hong, Y.-C.; Ha, M.; Park, H.; Kim, B.-N.; Lee, B.-E.; Lee, K.Y.; Kim, J.H.; et al. Performance IQ in children is associated with blood cadmium concentration in early pregnancy. J. Trace Elem. Med. Biol. 2015, 30, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Barranco, M.; Lacasana, M.; Gil, F.; Lorca, A.; Alguacil, J.; Rohlman, D.S.; Gonzalez-Alzaga, B.; Molina-Villalba, I.; Mendoza, R.; Agiuilar-Garduno, C. Cadmium exposure and neuropsychologic development in school children in southwestern Spain. Environ. Res. 2014, 134, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R. Regulating heavy metals in baby foods: The challenges of food manufacturers and the FDA being on the same page. Spectroscopy 2021, 36, 10–16. [Google Scholar]
- Karlsson, O.; Rockloev, J.; Lehoux, A.P.; Bergquist, J.; Rutgersson, A.; Blunt, M.J.; Birnbaum, L.S. The human exposome and health in the Anthropocene. Int. J. Epidemiol. 2021, 50, 378–389. [Google Scholar] [CrossRef] [PubMed]
- Tokumoto, M.; Lee, J.-Y.; Fujiwara, Y.; Satoh, M. Long-term expsoure to cadmium causes hepatic iron deficiency through the suppression of iron-transport-related gene expression in the proximal duodenum. Toxics 2023, 11, 641. [Google Scholar] [CrossRef]
- Fuller, R.; Landrigan, P.J.; Balakrishnan, K.; Bathan, G.; Bose-O’Rielly, S.; Brauer, M.; Caravanos, J.; Chiles, T.; Cohen, A.; Corra, L.; et al. Pollution and health: A progress update. Lancet Planet Health 2022, 6, e535–e547. [Google Scholar] [CrossRef]
- Molina, M.J.; Rowland, F.S. Stratospheric sink for chlorofluoromethanes: Chlorine atomc-atalysed destruction of ozone. Nature 1974, 249, 810–812. [Google Scholar] [CrossRef]
- Haney, J. Consideration of non-linear, non-threshold and threshold approaches for assessing the carcinogenicity of oral exposure to hexavalent chromium. Regul. Toxicol. Pharmacol. 2015, 73, 834–852. [Google Scholar] [CrossRef]
- Greenberg, M.I.; Vearrier, D. Metal fume fever and polymer fume fever. Clin. Toxicol. 2015, 53, 195–203. [Google Scholar] [CrossRef]
- Enwonwu, C.O. Potential health hazard of use of mercury in dentistry: Critical review of the literature. Environ. Res. 1987, 42, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Bates, M.N.; Fawcett, J.; Garrett, N.; Cutress, T.; Kjellstrom, T. Health effects of dental amalgam exposure: A retrospective cohort study. Int. J. Epidemiol. 2004, 33, 894–902. [Google Scholar] [CrossRef] [PubMed]
- Wild, C.P. Complementing the genome with an “exposome”: The outstanding challenge of environmental exposure measurement in molecular epidemiology. Cancer Epidemiol. Biomark. Prev. 2005, 14, 1847–1850. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.W.; Banbury Exposomics Consortium. Integrating exposomics into biomedicine. Assessing a full range of environmental exposures will improve human health. Science 2025, 338, 356–358. [Google Scholar] [CrossRef]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; SErrano, M.; Kroemer, G. The hallmarks of ageing. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef]
- Pourzadi, N.; Gailer, J. The emerging application of LC-based metallomics techniques to unravel the bioinorganic chemistry of toxic metal(loid)s. J. Chromatogr. A 2024, 1736, 465409. [Google Scholar] [CrossRef]
- Sarpong-Kumankomah, S.; Gibson, M.A.; Gailer, J. Organ damage by toxic metals is critically determined by the bloodstream. Coord. Chem. Rev. 2018, 374, 376–386. [Google Scholar] [CrossRef]
- Hafey, M.J.; Aleksunes, L.M.; Bridges, C.C.; Brouwer, K.R.; Chieh, H.-C.; Leslie, E.M.; Hu, S.; Li, Y.; Shen, J.; Sparreboom, A.; et al. Transporters and toxicity: Insights from the international transporter consortium workshop 4. Clin. Pharmacol. Ther. 2022, 112, 527–539. [Google Scholar] [CrossRef] [PubMed]
- Rayman, M.P.; Infante, H.G.; Sargent, M. Food-chain selenium and human health: Spotlight on speciation. Br. J. Nutr. 2008, 100, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Alarcon, M.; Cabrera-Vique, C. Selenium in food and the human body: A review. Sci. Total Environ. 2008, 400, 115–141. [Google Scholar] [CrossRef] [PubMed]
- Ponomarenko, O.; LaPorte, P.F.; Singh, S.P.; Langan, G.; Fleming, D.E.B.; Spallholz, J.; Alauddin, M.; Ahsan, H.; Ahmed, S.; Gailer, J.; et al. Selenium-mediated arsenic excretion in mammals: A synchrotron-based study of whole-body distribution and tissue-specific chemistry. Metallomics 2017, 9, 1585–1595. [Google Scholar] [CrossRef]
- Gailer, J.; Ruprecht, L.; Reitmeir, P.; Benker, B.; Schramel, P. Mobilization of exogenous and endogenous selenium to bile after the intravenous administration of environmentally relevant doses of arsenite to rabbits. Appl. Organometal. Chem. 2004, 18, 670–675. [Google Scholar] [CrossRef]
- Payliss, B.J.; Hassanin, M.; Prenner, E.J. The structural and functional effects of Hg(II) and Cd(II) on lipid model systems and human erythrocytes: A review. Chem. Phys. Lipids 2015, 193, 36–51. [Google Scholar] [CrossRef]
- Kerek, E.; Hassanin, M.; Prenner, E.J. Inorganic mercury and cadmium induce rigidity in eukaryotic lipid extracts while mercury also ruptures red blood cells. Biochim. Biophys. Acta 2018, 1860, 710–717. [Google Scholar] [CrossRef] [PubMed]
- Rother, R.P.; Bell, L.; Hillmen, P.; Gladwin, M.T. The clinical sequelae of intravascular hemolysis and extracellular plasma hemoglobin. J. Am. Med. Assoc. 2005, 293, 1653–1662. [Google Scholar] [CrossRef]
- Zalups, R.K.; Ahmad, S. Molecular handling of cadmium in transporting epithelia. Toxicol. Appl. Pharmacol. 2003, 186, 163–188. [Google Scholar] [CrossRef]
- Prozialeck, W.C.; Edwards, J.R.; Woods, J.M. The vascular endothelium as a target of cadmium toxicity. Life Sci. 2006, 79, 1493–1506. [Google Scholar] [CrossRef]
- Hill, A.; Gailer, J. Linking molecular targets of Cd in the bloodstream to organ-based adverse health effects. J. Inorg. Biochem. 2021, 216, 111279. [Google Scholar] [CrossRef]
- Gautam, A.; Gailer, J. More effective mobilization of Hg2+ from human serum albumin compared to Cd2+ by L-cysteine at near physiological conditions. Toxics 2023, 11, 599. [Google Scholar] [CrossRef] [PubMed]
- Farina, M.; Rocha, J.B.T.; Aschner, M. Mechanisms of methylmercury-induced neurotoxicity: Evidence from experimental studies. Life Sci. 2011, 89, 555–563. [Google Scholar] [CrossRef]
- Maret, W. Metallomics. A Primer of Integrated Biometal Sciences; Imperial College Press: London, UK, 2016. [Google Scholar]
- Shin, S.-Y.; Fauman, E.B.; Petersen, A.-K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.-P.; et al. An atlas of genetic influences on human blood metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Katrusiak, A.E.; Paterson, P.G.; Kamencic, H.; Shoker, A.; Lyon, A.W. Pre-column derivatization high-performance liquid chromatographic method for determination of cysteine, cysteinyl-glycine, homocysteine and glutathione in plasma and cell extracts. J. Chromatogr. B 2001, 758, 207–212. [Google Scholar] [CrossRef]
- Huang, J.; Williams, M. Total plasma homocysteine determination using ion-exchange chromatography. J. Liq. Chromatogr. 2000, 23, 3143–3153. [Google Scholar] [CrossRef]
- Leslie, E.M.; Liu, J.; Klaassen, C.D.; Waalkes, M.P. Acquired cadmium resistance in metallothionein-I/II(−/−) knockout cells: Role of the T-type calcium channel Cacna1G in cadmium uptake. Mol. Pharmacol. 2002, 69, 629–639. [Google Scholar] [CrossRef]
- Thomas, D.J.; Smith, J.C. Effects of coadministered low-molecular weight thiol compounds on short-term distribution of methylmercury in the rat. Toxicol. Appl. Pharmacol. 1982, 62, 104–110. [Google Scholar] [CrossRef] [PubMed]
- House, D.A.; Robinson, W.T.; McKee, V. Chloromercury (II) anions. Coord. Chem. Rev. 1994, 135/136, 533–586. [Google Scholar] [CrossRef]
- Wiberg, H. Das Cadmium. In Lehrbuch der Anorganischen Chemie; Wiberg, E., Ed.; Walter de Gruyter: Berlin, Germany, 1976; p. 829. [Google Scholar]
- Gautam, A.; Gomez, A.; Mendoza Rengifo, E.; George, G.N.; Pickering, I.J.; Gailer, J. Structural characterization of toxicologically relevant Cd2+-L-Cysteine complexes. Toxics 2023, 11, 294. [Google Scholar] [CrossRef]
- Craig, W.Y.; Ledue, T.B.; Ritchie, R.F. Plasma Proteins. Clinical Utility and Interpretation; Dade Behring Inc.: Newark, CA, USA, 2000. [Google Scholar]
- Azzini, E.; Ruggeri, S.; Polito, A. Homocysteine: Its possible emerging role in at-risk population groups. Int. J. Mol. Sci. 2020, 21, 1421. [Google Scholar] [CrossRef]
- Bridle, T.G.; Doroudian, M.; White, W.; Gailer, J. Physiologically relevant hCys concentrations mobilize MeHg from rabbit serum albumin to form MeHg-hCys complexes. Metallomics 2022, 14, mfac010. [Google Scholar] [CrossRef]
- Medieu, A.; Point, D.; Sonke, J.E.; Angot, H.; Allain, V.; Bodin, N.; Adams, D.H.; Bignert, A.; Streets, D.G.; Buchanan, P.B.; et al. Stable tuna mercury concentrations since 1971 illustrate marine inertia and the need for strong emission reductions under the Minamata Convention. Environ. Sci. Technol. Lett. 2024, 11, 250–258. [Google Scholar] [CrossRef]
- Ledda, C.; Cannizzaro, E.; Lovreglio, P.; Vitale, E.; Stufano, A.; Montana, A.; Li Volti, G.; Rapisarda, V. EXposure to toxic heavy metals can influence homocysteine metabolism? Antioxidants 2020, 9, 30. [Google Scholar] [CrossRef]
- Wang, W.; Rusin, O.; Xu, X.; Kim, K.K.; Escobedo, J.O.; Fakayode, S.O.; Fletcher, K.A.; Lowry, M.; Schowalter, C.M.; Lawrence, C.M.; et al. Detection of homocysteine and cysteine. J. Am. Chem. Soc. 2005, 127, 15949–15958. [Google Scholar] [CrossRef]
- Grzych, G.; Douillard, C.; Lannoy, J.; Curt, M.J.C. Very high plasma homocysteine without malnutrition or inherited disorder. Clin. Chem. 2020, 66, 1468–1469. [Google Scholar] [CrossRef]
- Shibeeb, S.; Abdallah, A.; Shi, Z. Blood homocysteine levels mediate the association between blood lead levels and cardiovascular disease. Cardiovasc. Toxicol. 2024, 24, 62–70. [Google Scholar] [CrossRef]
- Doroudian, M.; Pourzadi, N.; Gautam, A.; Gailer, J. Translational toxicology of metal(loid) species: Linking their bioinorganic chemistry in the bloodstream to organ damage onset. Biometals 2024, 37, 739–753. [Google Scholar] [CrossRef] [PubMed]
- Chapman, L.; Chan, H.M. The influence of nutrition on methyl mercury intoxication. Environ. Health Perspect. 2000, 108, 29–56. [Google Scholar] [PubMed]
- Buenzli, J.-C.G. Grand challenges in inorganic chemistry: Toward better life quality and a more sustainable world. Front. Chem. 2013, 1, 1–3. [Google Scholar] [CrossRef]
- Lahner, B.; Gong, J.; Mahmoudian, M.; Smith, E.L.; Abid, K.B.; Rogers, E.E.; Guerinot, M.L.; Harper, J.F.; Ward, J.M.; McIntyre, L.; et al. Genomic scale profiling of nutrient and trace elements in Arabidopsis thaliana. Nat. Biotechnol. 2003, 21, 1215–1221. [Google Scholar] [CrossRef]
- Issah, I.; Duah, M.S.; Arko-Mensah, J.; Bawua, S.A.; Agyekum, T.P.; Fobil, J.N. Exposure to metal mixture and adverse pregnancy and birth outcomes: A systematic review. Sci. Total Environ. 2024, 908, 168380. [Google Scholar] [CrossRef]
- Rider, C.V.; McHale, C.M.; Webster, T.F.; Lowe, L.; Goodson, W.H.; La Merrill, M.A.; Rice, G.; Zeise, L.; Zhang, L.; Smith, M.T. Using the key characteristics of carcinogens to develop research on chemical mixtures and cancer. Environ. Health Perspect. 2021, 129, 035003-1–035003-11. [Google Scholar] [CrossRef] [PubMed]
- Rizivi, S.A.A. Health systems strategies to reduce global impacts of toxic substance exposure. Acad. Med. 2025, 2, 1–7. [Google Scholar] [CrossRef]
- Sebastian, A.; Prasad, M.N.V. Cadmium minimization in rice: A review. Agron. Sustain. Dev. 2014, 34, 155–173. [Google Scholar] [CrossRef]
- Lodeiro, C.; Capelo-Martinez, J.L.; Santos, H.M.; Oliveira, E. Impacts of environmental issues on health and well-being: A global pollution challenge. Environ. Sci. Pollut. Res. 2021, 28, 18309–18313. [Google Scholar] [CrossRef] [PubMed]
- Radfard, M.; Hashemi, H.; Baghapour, M.A.; Samaei, M.R.; Yunesian, M.; Soleimani, H.; Azhdarpoor, A. Prediction of human health rsik and disability-adjusted life years induced by heavy metals exposure through drinking water in Fars Province, Iran. Sci. Rep. 2023, 13, 19080. [Google Scholar] [CrossRef]
- Martins, A.C.; Ferrer, B.; Tinkov, A.A.; Caito, S.; Deza-Ponzio, R.; Skalny, A.V.; Bowman, A.B.; Aschner, M. Association between heavy metals, metalloids and metabolic syndrome: New insights and approaches. Toxics 2023, 11, 670. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Degorge, M.F.; Gailer, J. Exposure–Response Relationship of Toxic Metal(loid)s in Mammals: Their Bioinorganic Chemistry in Blood Is an Intrinsic Component of the Selectivity Filters That Mediate Organ Availability. Toxics 2025, 13, 636. https://doi.org/10.3390/toxics13080636
Degorge MF, Gailer J. Exposure–Response Relationship of Toxic Metal(loid)s in Mammals: Their Bioinorganic Chemistry in Blood Is an Intrinsic Component of the Selectivity Filters That Mediate Organ Availability. Toxics. 2025; 13(8):636. https://doi.org/10.3390/toxics13080636
Chicago/Turabian StyleDegorge, Manon Fanny, and Jürgen Gailer. 2025. "Exposure–Response Relationship of Toxic Metal(loid)s in Mammals: Their Bioinorganic Chemistry in Blood Is an Intrinsic Component of the Selectivity Filters That Mediate Organ Availability" Toxics 13, no. 8: 636. https://doi.org/10.3390/toxics13080636
APA StyleDegorge, M. F., & Gailer, J. (2025). Exposure–Response Relationship of Toxic Metal(loid)s in Mammals: Their Bioinorganic Chemistry in Blood Is an Intrinsic Component of the Selectivity Filters That Mediate Organ Availability. Toxics, 13(8), 636. https://doi.org/10.3390/toxics13080636