



Long-Term Exposure to Cadmium Causes Hepatic Iron Deficiency through the Suppression of Iron-Transport-Related Gene Expression in the Proximal Duodenum


Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Animal Treatments
2.3. Hepatotoxicity
2.4. Real-Time RT-PCR Analysis
2.5. Cd and Fe Concentrations
2.6. Serum UIBC and TIBC Quantification
2.7. Statistical Analysis
3. Results
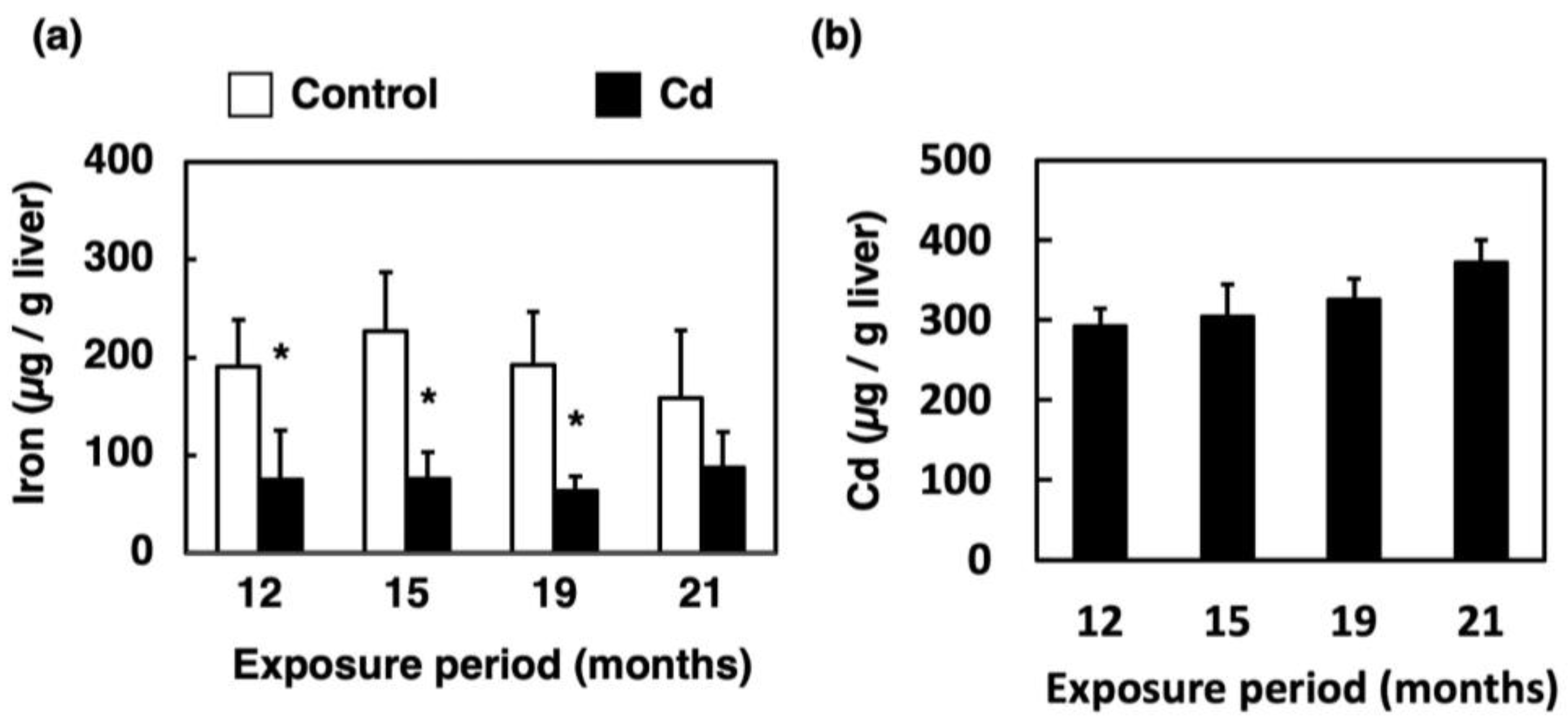
3.1. Effect of Long-Term Exposure to Cd on Accumulation of Iron and Cd in the Liver
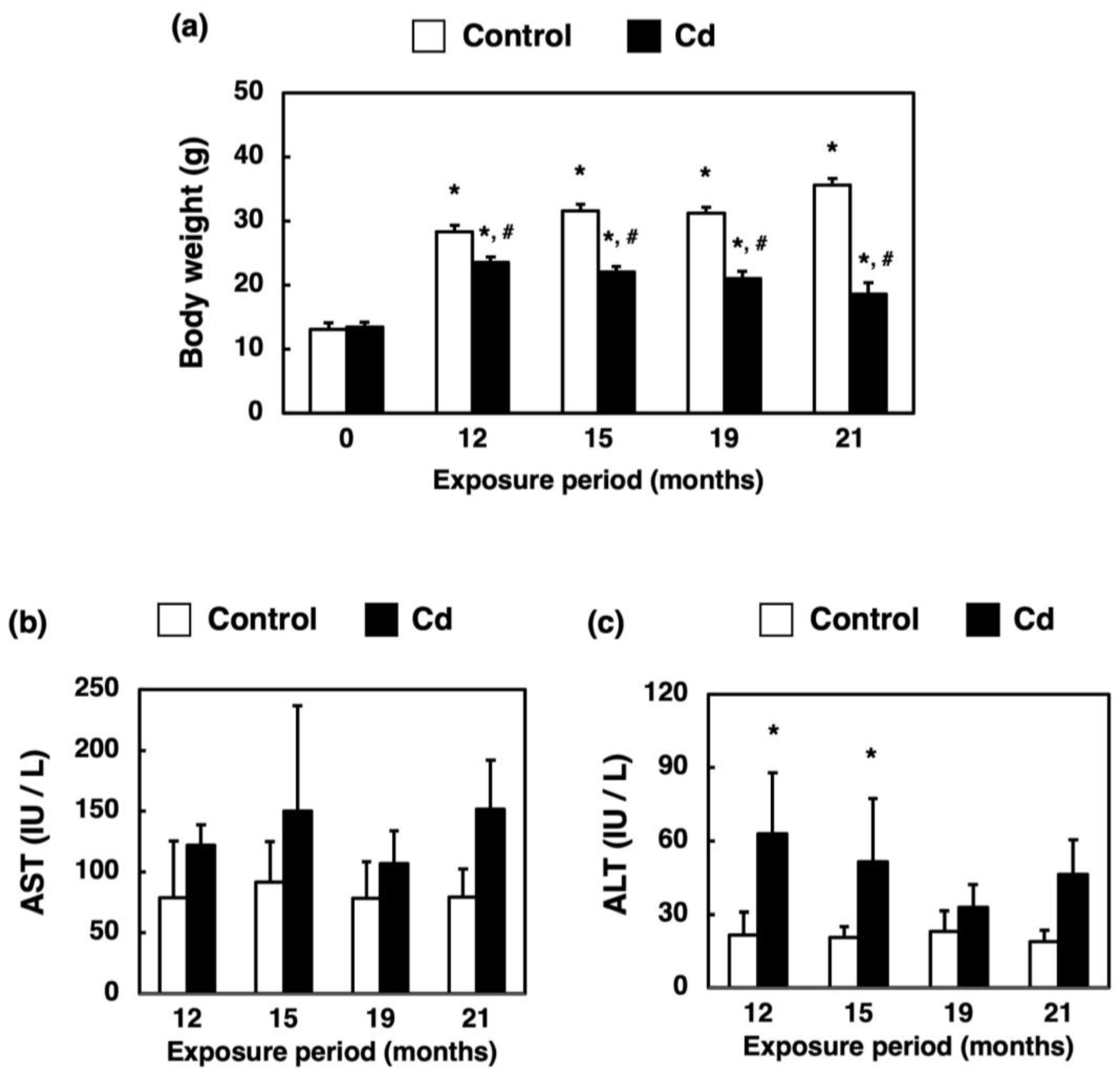
3.2. Changes in Body Weight and Hepatotoxicity in Mice Exposed to Cd for a Long Time
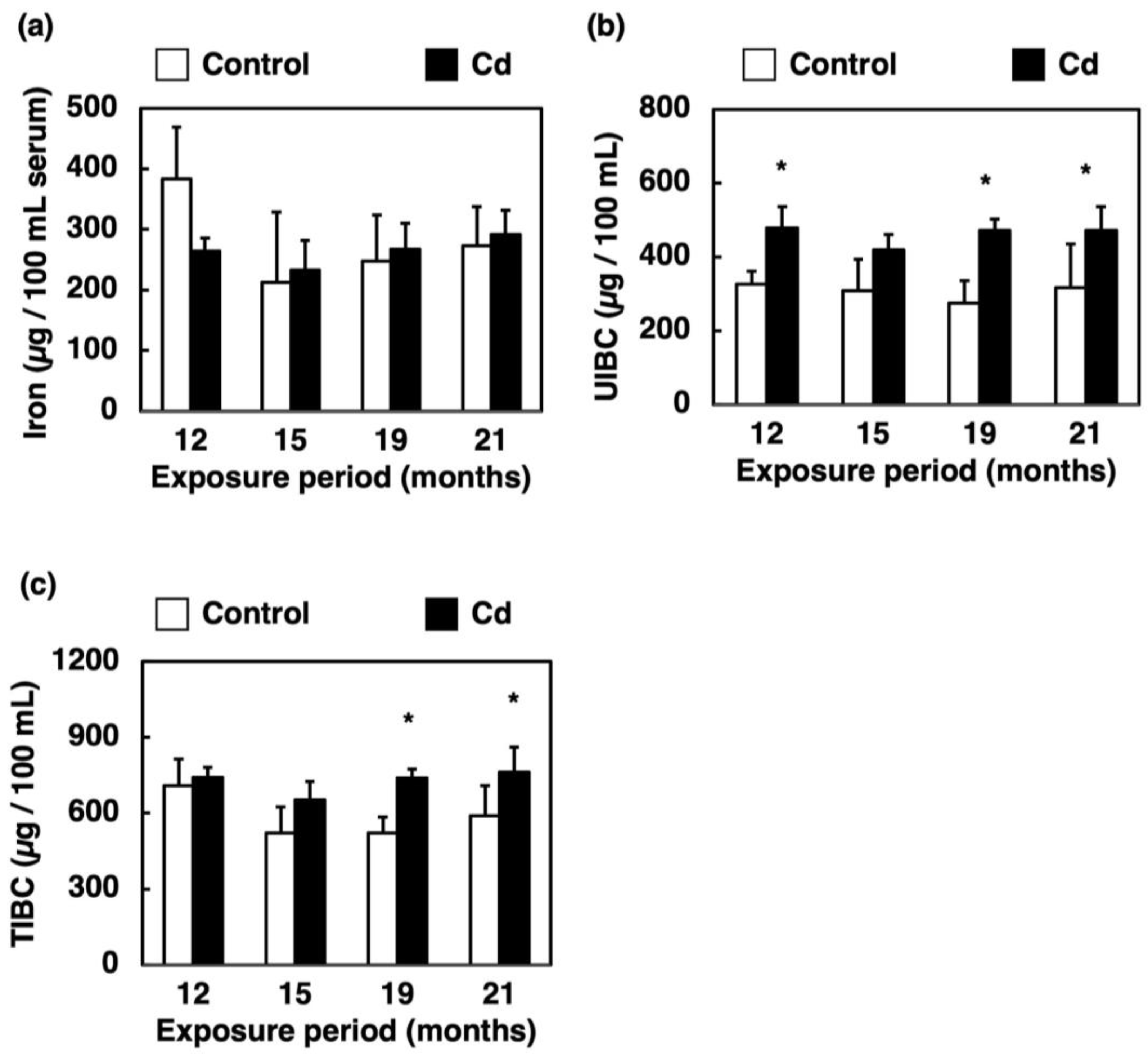
3.3. Effect of Long-Term Exposure to Cd on Iron Concentration, UIBC and TIBC in the Serum
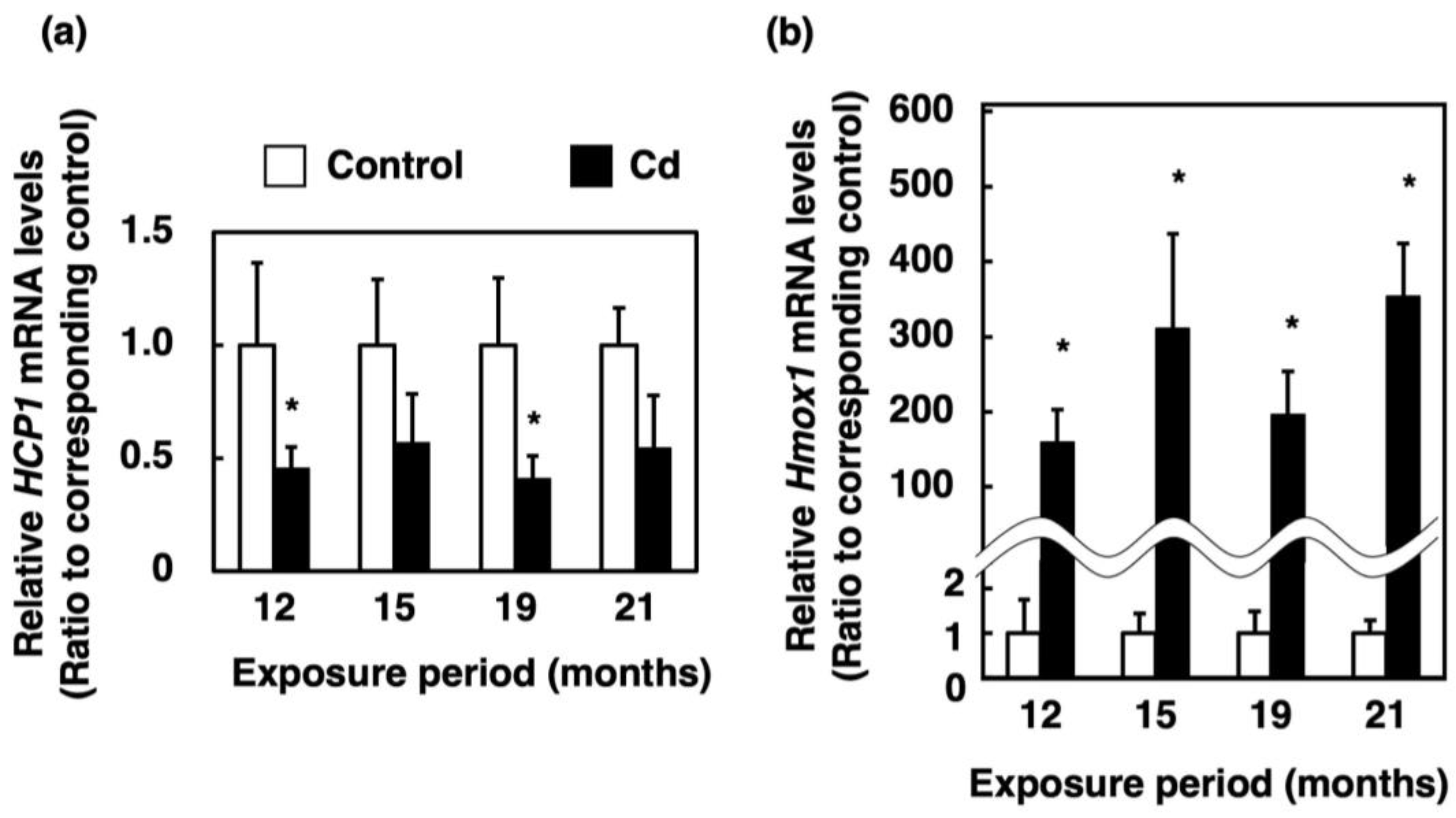
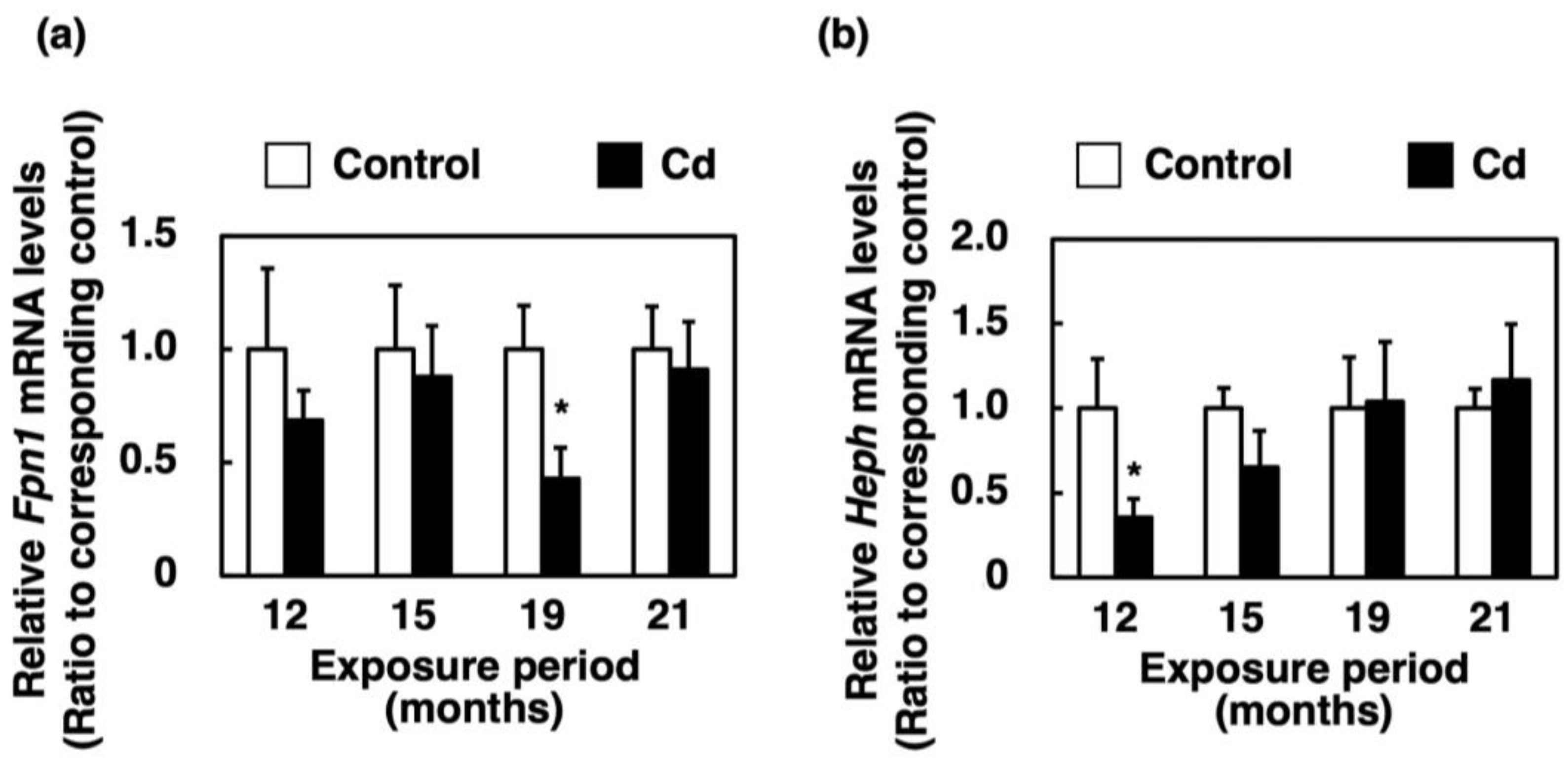
3.4. Effect of Long-Term Exposure to Cd on the Expression of Genes Involved in Absorption of Heme Iron in the Proximal Duodenum
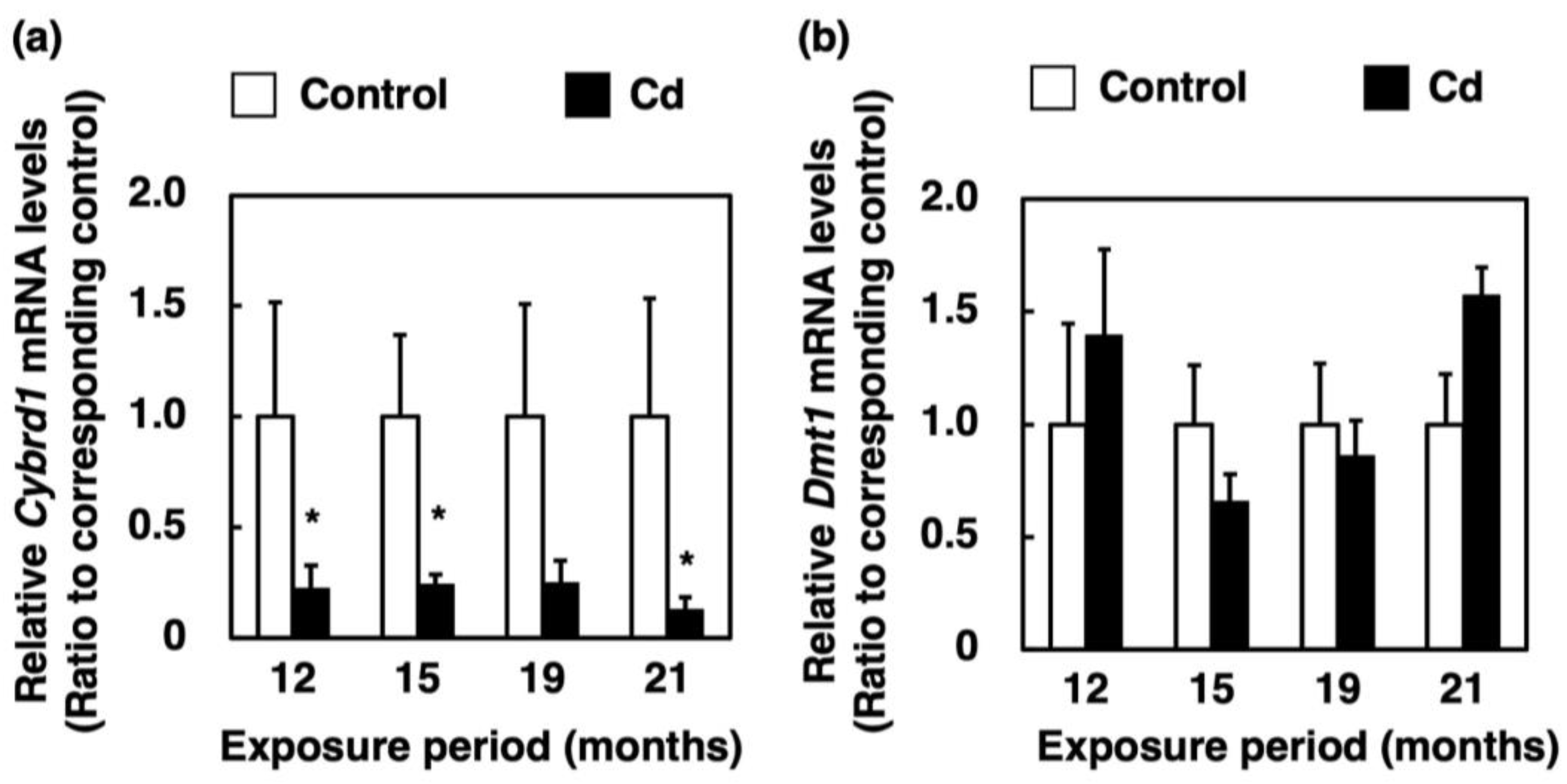
3.5. Effect of Long-Term Exposure to Cd on the Expression of Genes Involved in Absorption of Non-Heme Iron in the Proximal Duodenum
3.6. Effect of Long-Term Exposure to Cd on the Expression of Genes Involved in the Efflux of Iron from the Proximal Duodenum Enterocytes into Blood Vessels
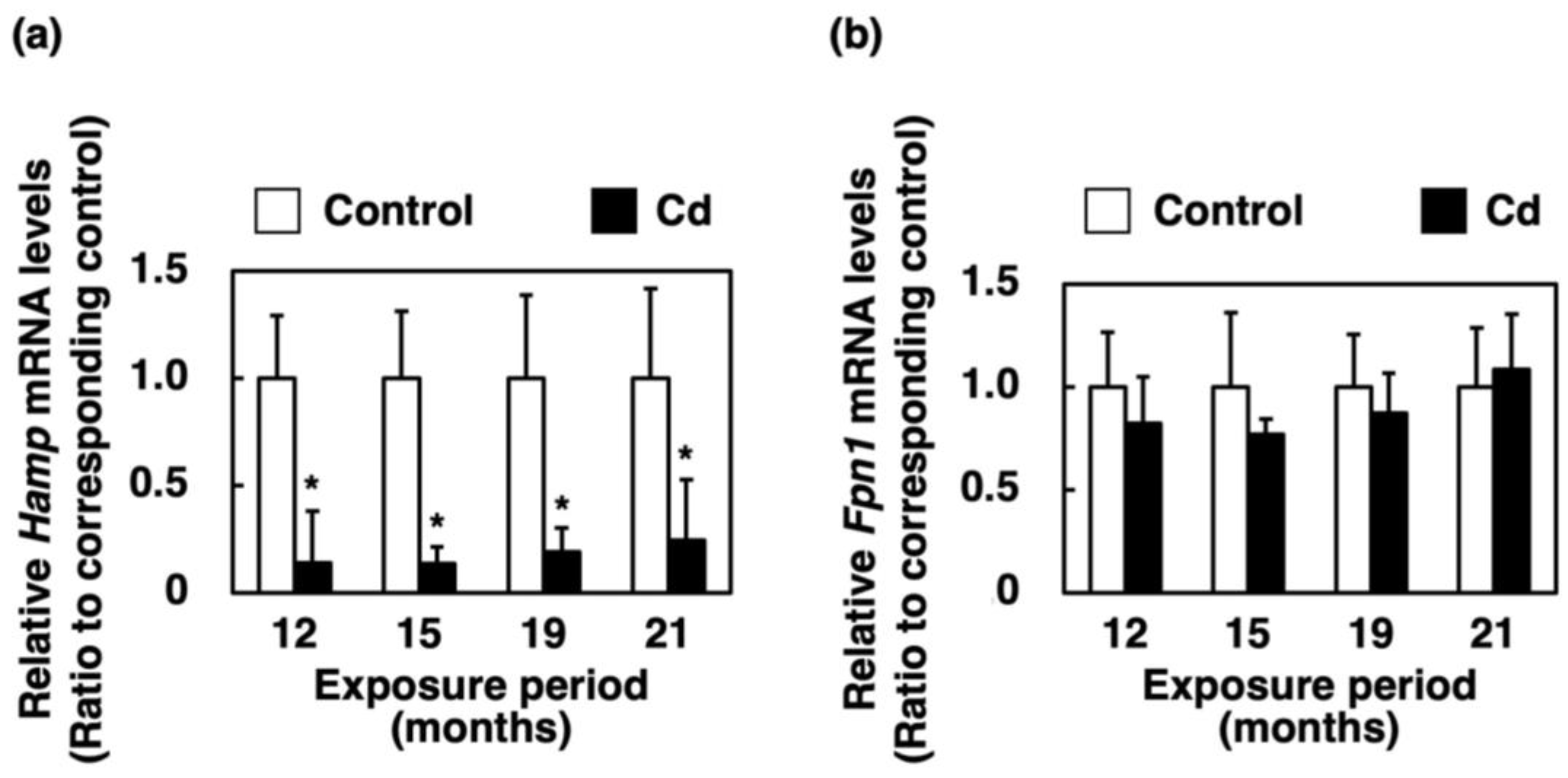
3.7. Effect of Long-Term Exposure to Cd on Gene Expression of Hamp and Fpn1 in the Liver
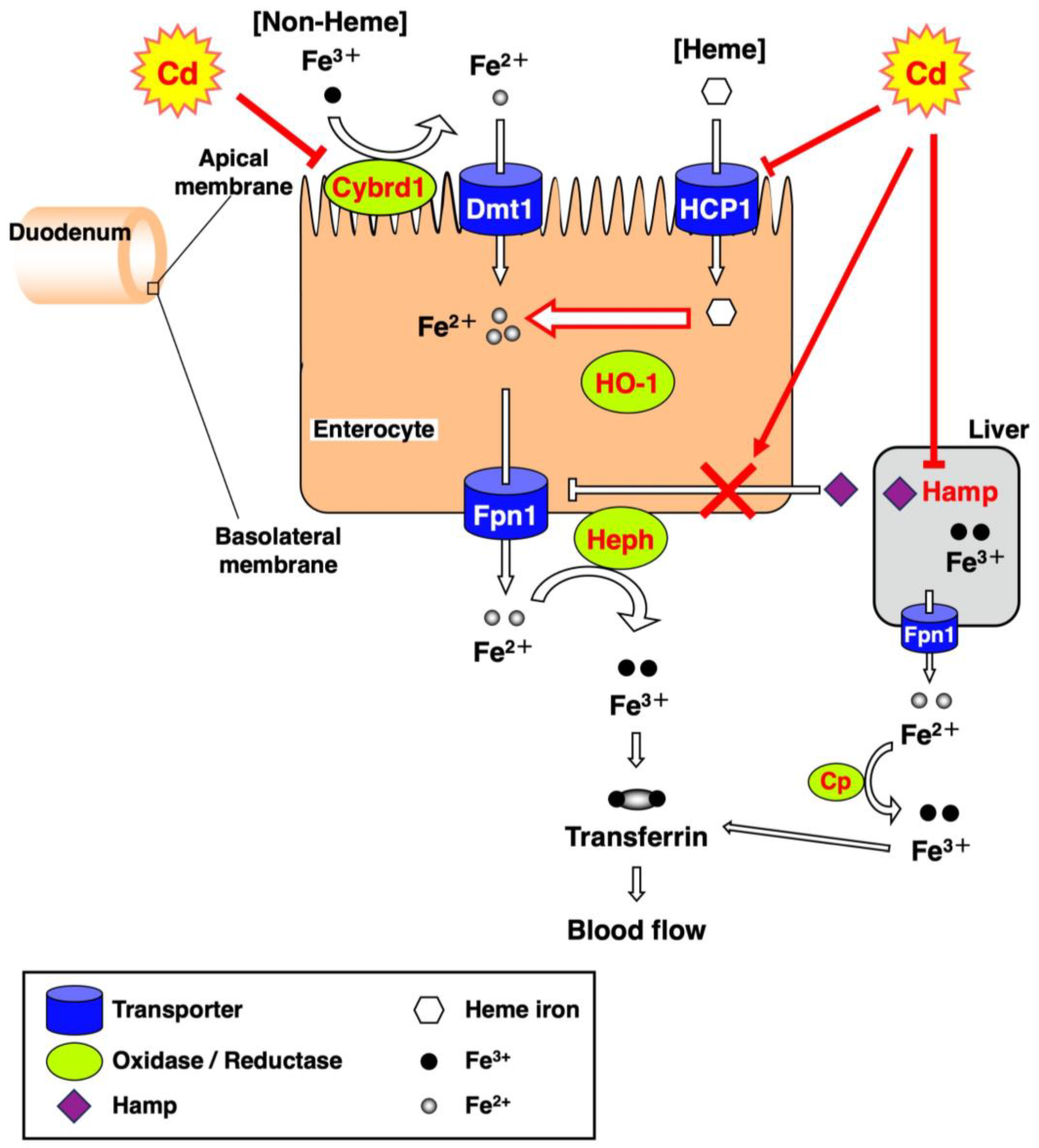
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jarup, L.; Berglund, M.; Elinder, C.G.; Nordberg, G.; Vahter, M. Health effects of cadmium exposure—A review of the literature and a risk estimate. Scand. J. Work Environ. Health 1998, 24 (Suppl. S1), 1–51. [Google Scholar] [PubMed]
- Satoh, M.; Koyama, H.; Kaji, T.; Kito, H.; Tohyama, C. Perspectives on cadmium toxicity research. Tohoku J. Exp. Med. 2002, 196, 23–32. [Google Scholar] [CrossRef]
- Zalups, R.K.; Ahmad, S. Molecular handling of cadmium in transporting epithelia. Toxicol. Appl. Pharmacol. 2003, 186, 163–188. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.K.; Kim, Y. Iron deficiency is associated with increased levels of blood cadmium in the Korean general population: Analysis of 2008–2009 Korean National Health and Nutrition Examination Survey data. Environ. Res. 2012, 112, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Silver, M.K.; Lozoff, B.; Meeker, J.D. Blood cadmium is elevated in iron deficient U.S. children: A cross-sectional study. Environ. Health 2013, 12, 117. [Google Scholar] [CrossRef]
- Suh, Y.J.; Lee, J.E.; Lee, D.H.; Yi, H.G.; Lee, M.H.; Kim, C.S.; Nah, J.W.; Kim, S.K. Prevalence and Relationships of Iron Deficiency Anemia with Blood Cadmium and Vitamin D Levels in Korean Women. J Korean Med. Sci. 2016, 31, 25–32. [Google Scholar] [CrossRef]
- Turgut, S.; Polat, A.; Inan, M.; Turgut, G.; Emmungil, G.; Bican, M.; Karakus, T.Y.; Genc, O. Interaction between anemia and blood levels of iron, zinc, copper, cadmium and lead in children. Indian J. Pediatr. 2007, 74, 827–830. [Google Scholar] [CrossRef]
- Djukić-Cosić, D.; Curcić Jovanović, M.; Plamenac Bulat, Z.; Ninković, M.; Malicević, Z.; Matović, V. Relation between lipid peroxidation and iron concentration in mouse liver after acute and subacute cadmium intoxication. J. Trace Elem. Med. Biol. 2008, 22, 66–72. [Google Scholar] [CrossRef]
- Sugawara, N. Effect of dietary cadmium on serum and liver lipids in mice in relation to interaction of essential metals, zinc, copper and iron. J. Toxicol. Sci. 1984, 9, 29–36. [Google Scholar] [CrossRef]
- Timofeeva, S.N.; Kadikov, I.R.; Korchemkin, A.A.; Vafin, I.F. The effects of zinc and magnesium ingestion on essential trace-element accumulation in the organs of white rats upon cadmium exposure. BIO Web Conf. 2020, 27, 00110. [Google Scholar] [CrossRef]
- Whanger, P.D. Cadmium effects in rats on tissue iron, selenium, and blood pressure; blood and hair cadmium in some oregon residents. Environ. Health Perspect. 1979, 28, 115–121. [Google Scholar] [CrossRef][Green Version]
- Doyle, J.J.; Pfander, W.H. Interactions of cadmium with copper, iron, zinc, and manganese in ovine tissues. J. Nutr. 1975, 105, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Rama, R.; Planas, J. Dietary cadmium effect on iron metabolism in chickens. Biol. Trace Elem. Res. 1981, 3, 169–183. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Accelerating Anaemia Reduction: A Comprehensive Framework for Action. 2023. Available online: https://apps.who.int/iris/handle/10665/367661 (accessed on 23 June 2023).
- Shayeghi, M.; Latunde-Dada, G.O.; Oakhill, J.S.; Laftah, A.H.; Takeuchi, K.; Halliday, N.; Khan, Y.; Warley, A.; McCann, F.E.; Hider, R.C.; et al. Identification of an intestinal heme transporter. Cell 2005, 122, 789–801. [Google Scholar] [CrossRef]
- Gunshin, H.; Mackenzie, B.; Berger, U.V.; Gunshin, Y.; Romero, M.F.; Boron, W.F.; Nussberger, S.; Gollan, J.L.; Hediger, M.A. Cloning and characterization of a mammalian proton-coupled metal-ion transporter. Nature 1997, 388, 482–488. [Google Scholar] [CrossRef]
- McKie, A.T.; Barrow, D.; Latunde-Dada, G.O.; Rolfs, A.; Sager, G.; Mudaly, E.; Mudaly, M.; Richardson, C.; Barlow, D.; Bomford, A.; et al. An iron-regulated ferric reductase associated with the absorption of dietary iron. Science 2001, 291, 1755–1759. [Google Scholar] [CrossRef]
- Donovan, A.; Brownlie, A.; Zhou, Y.; Shepard, J.; Pratt, S.J.; Moynihan, J.; Paw, B.H.; Drejer, A.; Barut, B.; Zapata, A.; et al. Positional cloning of zebrafish ferroportin1 identifies a conserved vertebrate iron exporter. Nature 2000, 403, 776–781. [Google Scholar] [CrossRef]
- Vulpe, C.D.; Kuo, Y.M.; Murphy, T.L.; Cowley, L.; Askwith, C.; Libina, N.; Gitschier, J.; Anderson, G.J. Hephaestin, a ceruloplasmin homologue implicated in intestinal iron transport, is defective in the sla mouse. Nat. Genet. 1999, 21, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, E.; Ganz, T. Regulation of iron metabolism by hepcidin. Annu. Rev. Nutr. 2006, 26, 323–342. [Google Scholar] [CrossRef]
- Nemeth, E.; Tuttle, M.S.; Powelson, J.; Vaughn, M.B.; Donovan, A.; Ward, D.M.; Ganz, T.; Kaplan, J. Hepcidin regulates cellular iron efflux by binding to ferroportin and inducing its internalization. Science 2004, 306, 2090–2093. [Google Scholar] [CrossRef]
- Fujiwara, Y.; Lee, J.Y.; Banno, H.; Imai, S.; Tokumoto, M.; Hasegawa, T.; Seko, Y.; Nagase, H.; Satoh, M. Cadmium induces iron deficiency anemia through the suppression of iron transport in the duodenum. Toxicol. Lett. 2020, 332, 130–139. [Google Scholar] [CrossRef]
- Auerbach, M.; Adamson, J.W. How we diagnose and treat iron deficiency anemia. Am. J. Hematol. 2016, 91, 31–38. [Google Scholar] [CrossRef]
- Thijssen, S.; Maringwa, J.; Faes, C.; Lambrichts, I.; Van Kerkhove, E. Chronic exposure of mice to environmentally relevant, low doses of cadmium leads to early renal damage, not predicted by blood or urine cadmium levels. Toxicology 2007, 229, 145–156. [Google Scholar] [CrossRef]
- Drakesmith, H.; Nemeth, E.; Ganz, T. Ironing out Ferroportin. Cell Metab. 2015, 22, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Taketani, S.; Kohno, H.; Yoshinaga, T.; Tokunaga, R. The human 32-kDa stress protein induced by exposure to arsenite and cadmium ions is heme oxygenase. FEBS Lett. 1989, 245, 173–176. [Google Scholar] [CrossRef]
- Helman, S.L.; Zhou, J.; Fuqua, B.K.; Lu, Y.; Collins, J.F.; Chen, H.; Vulpe, C.D.; Anderson, G.J.; Frazer, D.M. The biology of mammalian multi-copper ferroxidases. Biometals 2023, 36, 263–281. [Google Scholar] [CrossRef] [PubMed]
- Hellman, N.E.; Gitlin, J.D. Ceruloplasmin metabolism and function. Annu. Rev. Nutr. 2002, 22, 439–458. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.Y.; David, S. Glycosylphosphatidylinositol-anchored ceruloplasmin is required for iron efflux from cells in the central nervous system. J. Biol. Chem. 2003, 278, 27144–27148. [Google Scholar] [CrossRef] [PubMed]
- Linder, M.C. Ceruloplasmin and other copper binding components of blood plasma and their functions: An update. Metallomics 2016, 8, 887–905. [Google Scholar] [CrossRef]
- Fuqua, B.K.; Lu, Y.; Darshan, D.; Frazer, D.M.; Wilkins, S.J.; Wolkow, N.; Bell, A.G.; Hsu, J.; Yu, C.C.; Chen, H.; et al. The multicopper ferroxidase hephaestin enhances intestinal iron absorption in mice. PLoS ONE 2014, 9, e98792. [Google Scholar] [CrossRef]
- Ohta, H.; Ohba, K. Involvement of metal transporters in the intestinal uptake of cadmium. J. Toxicol. Sci. 2020, 45, 539–548. [Google Scholar] [CrossRef] [PubMed]
- Leazer, T.M.; Liu, Y.; Klaassen, C.D. Cadmium absorption and its relationship to divalent metal transporter-1 in the pregnant rat. Toxicol. Appl. Pharmacol. 2002, 185, 18–24. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Names | Forward | Reverse | Product Size (bp) |
|---|---|---|---|
| Slc46a1 (HCP1) | ATCTACCCGGCCACTCTGAA | GGAACTCCGGGTGTGGATTA | 121 |
| Hmox1 | ACCTTCCCGAACATCGACAG | GGAAGGCGGTCTTAGCCTCT | 121 |
| Slc11a2 (Dmt1) | GGCTTTCTTATGAGCATTGCCTA | GGAGCACCCAGAGCAGCTTA | 97 |
| Cybrd1 | GCAGCGGGCTCGAGTTTA | TTCCAGGTCCATGGCAGTCT | 103 |
| Slc40a1 (Fpn1) | CTGTCGGCCAGATTATGACA | GAGCAGGGGTCTTCTGGTAA | 126 |
| Heph | TTGTCTCATGAAGAACATTTACAGCAC | CATATGGCAATCAAAGCAGAAGA | 161 |
| Hamp | GGCAGACATTGCGATACCAA | TGGCTCTAGGCTATGTTTTGCA | 128 |
| Actin, beta (Actb) | CCTAAGGCCAACCGTGAAAA | AGGCATACAGGGACAGCACA | 100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tokumoto, M.; Lee, J.-Y.; Fujiwara, Y.; Satoh, M. Long-Term Exposure to Cadmium Causes Hepatic Iron Deficiency through the Suppression of Iron-Transport-Related Gene Expression in the Proximal Duodenum. Toxics 2023, 11, 641. https://doi.org/10.3390/toxics11070641
Tokumoto M, Lee J-Y, Fujiwara Y, Satoh M. Long-Term Exposure to Cadmium Causes Hepatic Iron Deficiency through the Suppression of Iron-Transport-Related Gene Expression in the Proximal Duodenum. Toxics. 2023; 11(7):641. https://doi.org/10.3390/toxics11070641
Chicago/Turabian StyleTokumoto, Maki, Jin-Yong Lee, Yasuyuki Fujiwara, and Masahiko Satoh. 2023. "Long-Term Exposure to Cadmium Causes Hepatic Iron Deficiency through the Suppression of Iron-Transport-Related Gene Expression in the Proximal Duodenum" Toxics 11, no. 7: 641. https://doi.org/10.3390/toxics11070641
APA StyleTokumoto, M., Lee, J.-Y., Fujiwara, Y., & Satoh, M. (2023). Long-Term Exposure to Cadmium Causes Hepatic Iron Deficiency through the Suppression of Iron-Transport-Related Gene Expression in the Proximal Duodenum. Toxics, 11(7), 641. https://doi.org/10.3390/toxics11070641