


Effects of Glyphosate on Female Reproductive Output in the Marine Polychaete Worm Ophryotrocha diadema



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Model
2.2. Experimental Set Up
2.3. Statistical Analysis
3. Results
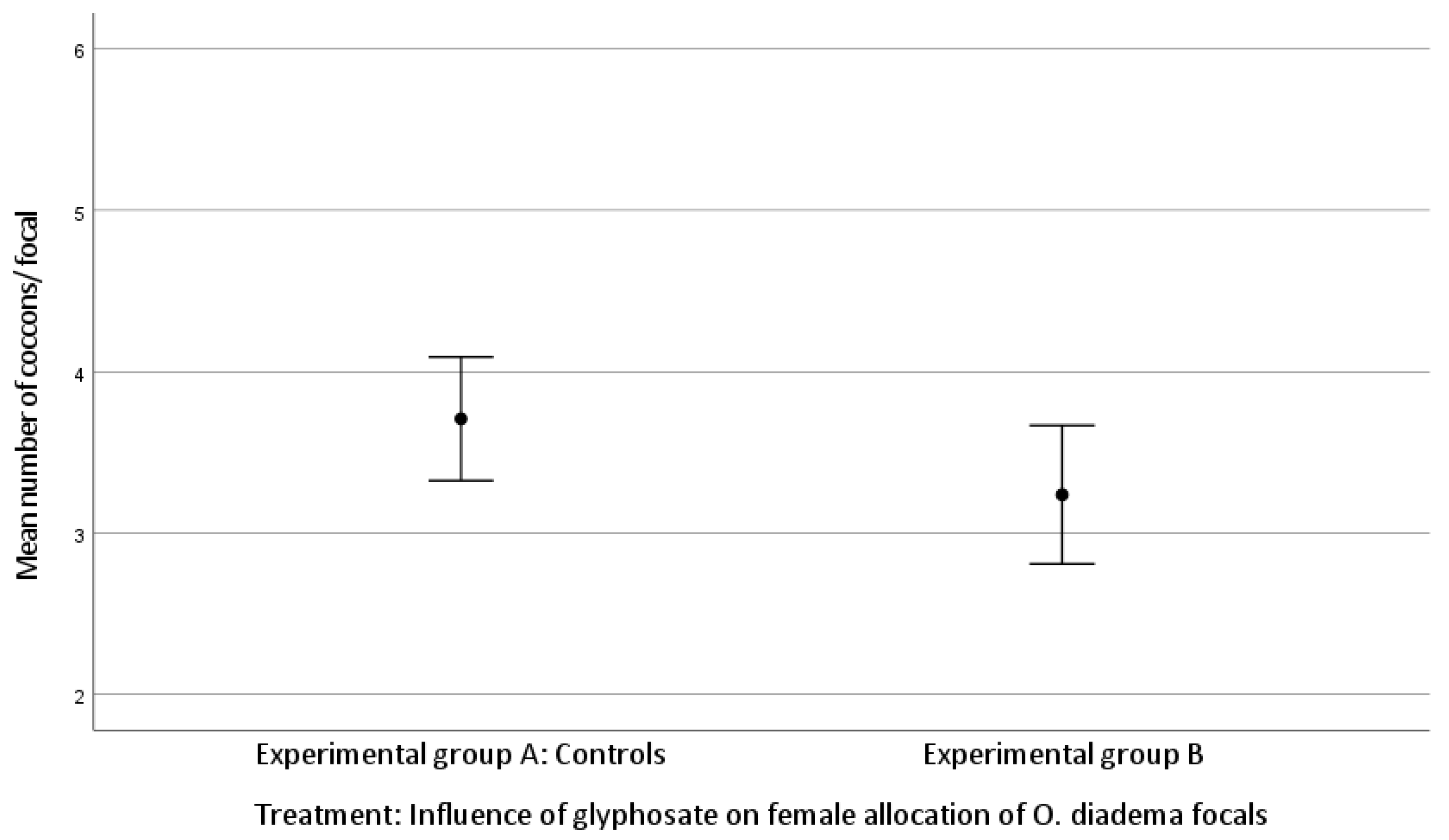
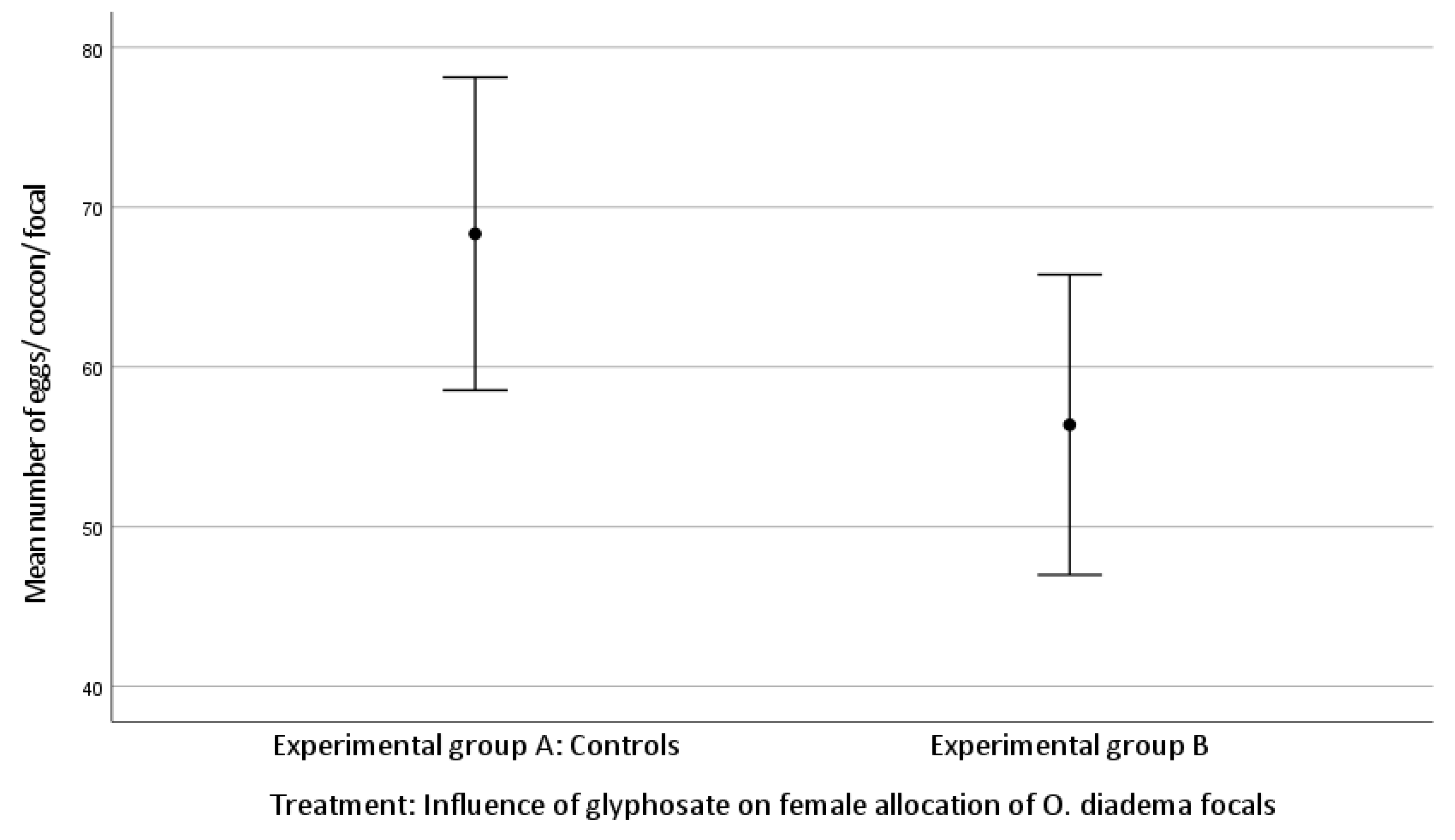
3.1. Effect of Glyphosate Exposure on Female Allocation in O. diadema Focals
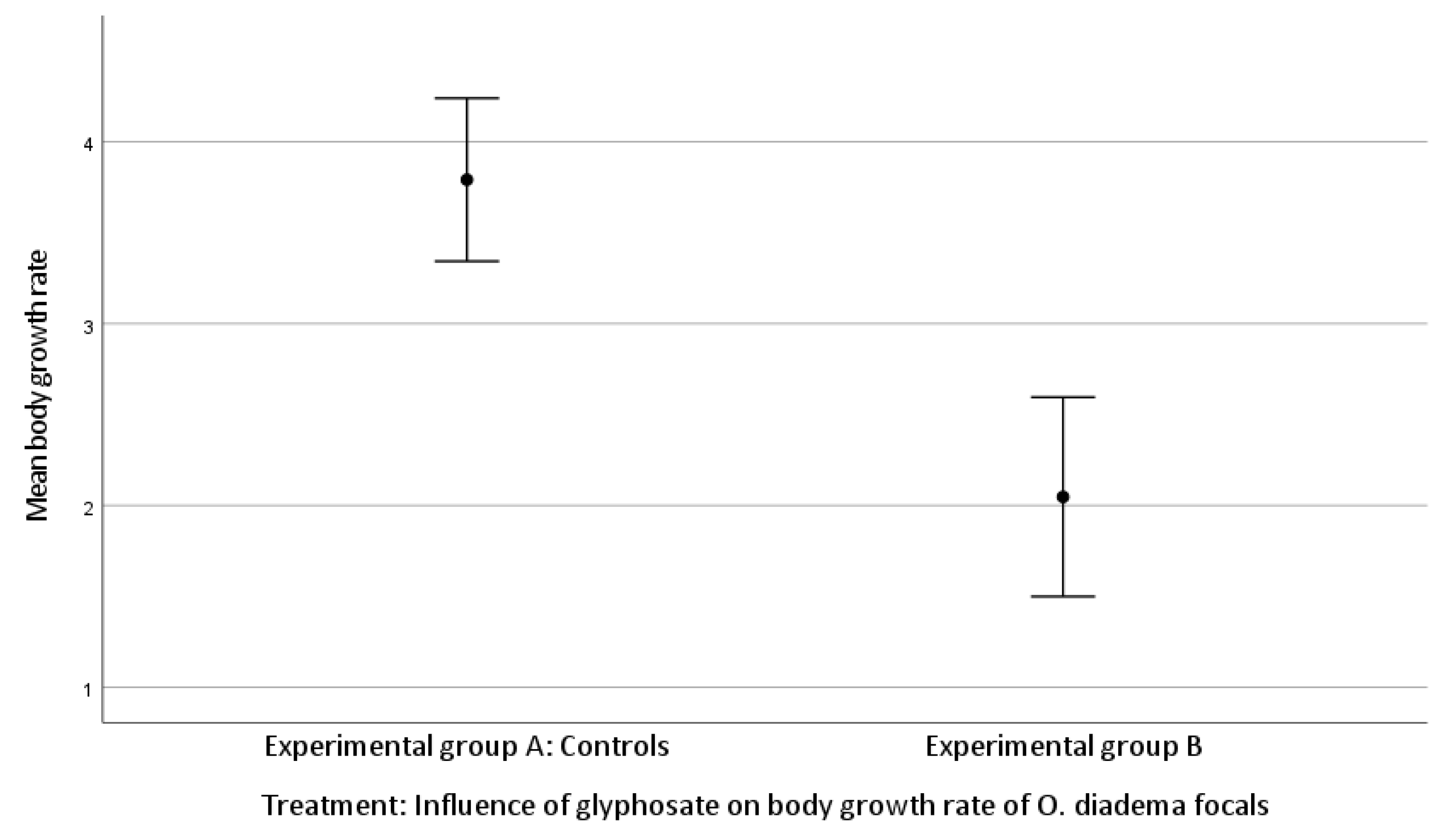
3.2. Effect of Glyphosate Exposure on Growth Rate in O. diadema Focals
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Blaustein, A.R.; Romansic, J.M.; Kiesecker, J.M.; Hatch, A.C. Ultraviolet radiation, toxic chemicals and amphibian population declines. Divers. Distrib. 2003, 9, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Stuart, N.S.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.L.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 3, 1783–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crain, D.A.; Eriksen, M.; Iguchi, T.; Jobling, S.; Laufer, H.; LeBlanc, G.A.; Guillette, L.J., Jr. An ecological assessment of bisphenol-A: Evidence from comparative biology. Reprod. Toxicol. 2007, 24, 225–239. [Google Scholar] [CrossRef]
- Duke, S.O. The history and current status of glyphosate. Pest Manag. Sci. 2017, 74, 1027–1034. [Google Scholar] [CrossRef]
- Bai, S.H.; Ogbourne, S.M. Glyphosate: Environmental contamination, toxicity and potential risks to human health via food contamination. Environ. Sci. Pollut. Res. 2016, 23, 18988–19001. [Google Scholar] [CrossRef]
- Baker, L.F.; Mudge, J.F.; Thompson, D.G.; Houlahan, J.E.; Kidd, K.A. The combined influence of two agricultural contaminants on natural communities of phytoplankton and zooplankton. Ecotoxicology 2016, 25, 1021–1032. [Google Scholar] [CrossRef]
- Baker, L.F.; Mudge, J.F.; Houlahan, J.E.; Thompson, D.G.; Kidd, K.A. The direct and indirect effects of a glyphosate-based herbicide and nutrients on Chrionomidae (Diptera) emerging from small wetlands. Environ. Toxicol. Chem. 2014, 33, 2076–2085. [Google Scholar] [CrossRef]
- King, J.J.; Wagner, R.S. Toxic effects of the herbicide roundup® regular on Pacific northwestern amphibians. Northwestern Nat. 2010, 91, 318–324. [Google Scholar] [CrossRef]
- Braz-Mota, S.; Sadauskas-Henrique, H.; Duarte, R.M.; Val, A.L.; Almeida-Val, V.M.F. Roundup® exposure promotes gills and liver impairments, DNA damage and inhibition of brain cholinergic activity in the Amazon teleost fish Colossoma Macropomum. Chemosphere 2015, 135, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Kelly, D.W.; Poulin, R.; Tompkins, D.M.; Townsend, C.R. Synergistic effects of glyphosate formulation and parasite infection on fish malformations and survival. J. Appl. Ecol. 2010, 47, 498–504. [Google Scholar] [CrossRef]
- Kreutz, L.C.; Barcellos, L.J.G.; Marteninghe, A.; dos Santos, E.D.; Zanatta, R. Exposure to sublethal concentration of glyphosate or atrazine-based herbicides alters the phagocytic function and increases the susceptibility of silver catfish fingerlings (Rhamdia quelen) to Aeromonas hydrophila challenge. Fish Shellfish Immunol. 2010, 29, 694–697. [Google Scholar] [CrossRef] [PubMed]
- Le Du-Carrée, J.; Cabon, J.; Louboutin, L.; Morin, T.; Danion, M. Changes in defense capacity to infectious hematopoietic necrosis virus (IHNv) in rainbow trout intergenerationally exposed to glyphosate. Fish Shellfish Immunol. 2022, 122, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Bu, Y.; Li, X. Immunological and histopathological responses of the kidney of common carp (Cyprinus carpio L.) sublethally exposed to glyphosate. Environ. Toxicol. Pharmacol. 2015, 29, 1–8. [Google Scholar] [CrossRef]
- Bali, Y.A.; Ba-Mhamed, S.; Bennis, M. Behavioral and Immunohistochemical study of the effects of subchronic and chronic exposure to glyphosate in mice. Front. Behav. Neurosci. 2017, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Romano, M.A.; Romano, R.M.; Santos, L.D.; Wisniewski, P.; Campos, D.A.; de Souza, P.B.; Viau, P.; Bernardi, M.M.; Nunes, M.T.; de Oliveira, C.A. Glyphosate impairs male offspring reproductive development by disrupting gonadotropin expression. Arch. Toxicol. 2012, 86, 663–673. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Jarosiewicz, P.; Bukowska, B. Glyphosate and its formulations—Toxicity, occupational and environmental exposure (Polish). Med. Pr. 2013, 64, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Thongprakaisang, S.; Thiantanawat, A.; Rangkadilok, N.; Suriyo, T.; Satayavivad, J. Glyphosate induces human breast cancer cells growth via estrogen receptors. Food Chem. Toxicol. 2013, 59, 129–136. [Google Scholar] [CrossRef]
- Kwiatkowska, M.; Jarosiewicz, P.; Michałowicz, J.; Koter-Michalak, M.; Huras, B.; Bukowska, B. The impact of glyphosate, its metabolites and impurities on viability, ATP level and morphological changes in human peripheral blood mononuclear cells. PLoS ONE 2016, 11, e0156946. [Google Scholar] [CrossRef] [Green Version]
- Kwiatkowska, M.; Reszka, E.; Katarzyna Wozniak, K.; Ewa Jabłonska, E.; Jaromir Michałowicz, J.; Bozena Bukowska, B. DNA damage and methylation induced by glyphosate in human peripheral blood mononuclear cells (in vitro study). Food Chem. Toxicol. 2017, 105, 93–98. [Google Scholar] [CrossRef]
- Madani, N.A.; Carpenter, D.O. Effects of glyphosate and glyphosate-based herbicides like Roundup™ on the mammalian nervous system: A review. Environ. Res. 2022, 214, 113933. [Google Scholar] [CrossRef]
- Santovito, A.; Ruberto, S.; Gendusa, C.; Cervella, P. In vitro evaluation of genomic damage induced by glyphosate on human lymphocytes. Environ. Sci. Pollut. Res. 2018, 25, 34693–34700. [Google Scholar] [CrossRef] [PubMed]
- de Melo Tarouco, F.; de Godoi, F.G.A.; Velasques, R.R.; da Silveira Guerreiro, A.; Geihs, M.A.; da Rosa, C.E. Effects of the herbicide Roundup on the polychaeta Laeonereis acuta: Cholinesterases and oxidative stress. Ecotoxicol. Environ. Saf. 2017, 135, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Amid, C.; Olstedt, M.; Gunnarsson, J.S.; Le Lan, H.; Tran Thi Minh, H.; Van den Brink, P.J.; Hellström, M.; Tedengren, M. Additive effects of the herbicide glyphosate and elevated temperature on the branched coral Acropora formosa in Nha Trang, Vietnam. Environ. Sci. Pollut. Res. 2018, 25, 13360–13372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Åkesson, B. Morphology and life cycle of Ophryotrocha diadema, a new polychaete species from California. Ophelia 1976, 15, 25–35. [Google Scholar] [CrossRef]
- Åkesson, B. A life table study on three genetic strains of Ophryotrocha diadema (Polychaeta, Dorvilleidae). Int. J. Invert. Reprod. 1982, 5, 59–69. [Google Scholar] [CrossRef]
- Sella, G. Reciprocal egg trading and brood care in a hermaphroditic polychaete worm. Anim. Behav. 1985, 33, 938–944. [Google Scholar] [CrossRef]
- Sella, G. Reciprocation, reproductive success, and safeguards against cheating in the mating system of a hermaphroditic polychaete worm, Ophryotrocha diadema. Biol. Bull. 1988, 175, 212–217. [Google Scholar] [CrossRef]
- Sella, G. Sex allocation in the simultaneous hermaphroditic polychaete worm Ophryotrocha diadema. Ecology 1990, 71, 27–32. [Google Scholar] [CrossRef]
- Sella, G. Evolution of biparental care in the hermaphroditic polychaete worm Ophryotrocha diadema. Evolution 1991, 45, 63–68. [Google Scholar] [CrossRef]
- Premoli, M.C.; Sella, G. Sex economy in benthic polychaetes. Ethol. Ecol. Evol. 1995, 7, 27–48. [Google Scholar] [CrossRef]
- Sella, G.; Lorenzi, M.C. Partner fidelity and egg reciprocation in the simultaneously hermaphroditic polychaete worm Ophryotrocha diadema. Behav. Ecol. 2000, 11, 260–264. [Google Scholar] [CrossRef]
- Westheide, W. The concept of reproduction in polychaetes with small body size. Fortsch. Zool. 1984, 29, 265–287. [Google Scholar]
- Berruti, G.; Ferraguti, M.; Lamia Donin, C.L. The aflagellate spermatozoon of Ophryotrocha: A line of evolution of fertilization among polychaetes. Gamete Res. 1978, 1, 278–292. [Google Scholar] [CrossRef]
- Ferreira, C.; Durán, R.; Faro, L.R.F. Toxic effects of glyphosate on the nervous system: A systematic review. Int. J. Mol. Sci. 2022, 23, 4605. [Google Scholar] [CrossRef]
- Bali, Y.A.; Kaikai, N.-E.; Ba-M’Hamed, S.; Bennis, M. Learning and memory impairments associated to acetylcholinesterase inhibition and oxidative stress following glyphosate based-herbicide exposure in mice. Toxicology 2019, 415, 18–25. [Google Scholar] [CrossRef]
- Bali, Y.A.; Ba-M’Hamed, S.; Gambarotta, G.; Sassoè-Pognetto, M.; Giustetto, M.; Bennis, M. Pre- and postnatal exposure to glyphosate-based herbicide causes behavioral and cognitive impairments in adult mice: Evidence of cortical ad hippocampal dysfunction. Arch. Toxicol. 2020, 94, 1703–1723. [Google Scholar] [CrossRef]
- McVey, K.A.; Snapp, I.B.; Johnson, M.B.; Negga, R.; Pressley, A.S.; Fitsanakis, V.A. Exposure of C. elegans eggs to a glyphosate containing herbicide leads to abnormal neuronal morphology. Neurotoxicol. Teratol. 2016, 55, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Burchfield, S.L.; Bailey, D.C.; Todt, C.E.; Denney, R.D.; Negga, R.; Fitsanakis, V.A. Acute exposure to a glyphosate-containing herbicide formulation inhibits Complex II and increases hydrogen peroxide in the model organism Caenorhabditis elegans. Environ. Toxicol. Pharmacol. 2018, 66, 36–42. [Google Scholar] [CrossRef]
- Myers, J.P.; Antoniou, M.N.A.; Blumberg, B.; Carroll, L.; Colborn, T.; Everett, L.G.; Hansen, M.; Landrigan. P., L.; Lanphear, B.P.; Mesnage, R.; et al. Concerns over use of glyphosate-based herbicides and risks associated with exposures: A consensus statement. Environ. Health 2016, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Uren Webster, T.M.; Laing, L.V.; Florance, H.; Santos, E.M. Effects of glyphosate and its formulation, Roundup, on reproduction in zebrafish (Danio rerio). Environ. Sci. Technol. 2014, 48, 1271–1279. [Google Scholar] [CrossRef]
- Reddy, S.B.; Nolan, C.J.; Zygar Plautz, C. Disturbances in reproduction and expression of steroidogenic enzymes in aquatic invertebrates exposed to components of the herbicide Roundup. Toxicol. Res. Appl. 2018, 2, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Asnicar, D.; Cappelli, C.; Sallehuddin, A.S.; Maznan, N.A.; Marin, M.G. Effects of glyphosate-based and derived products on sea urchin larval development. J. Mar. Sci. Eng. 2020, 8, 661. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Group | Treatment: [Glyphosate] | No. of ind./bowl | No. of Replicates |
|---|---|---|---|
| Control group A | 0.0 μg/mL Pure artificial sea water | 2 (1Yy − focal + 1yy) | 24 |
| Group B | 0.125 μg/mL | 2 (1Yy − focal + 1yy) | 24 |
| Group C | 0.250 μg/mL | 2 (1Yy − focal + 1yy) | 24 |
| Group D | 0.500 μg/mL | 2 (1Yy − focal + 1yy) | 24 |
| Group E | 1.000 μg/mL | 2 (1Yy − focal + 1yy) | 24 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schleicherová, D.; Prearo, M.; Di Nunno, C.; Santovito, A. Effects of Glyphosate on Female Reproductive Output in the Marine Polychaete Worm Ophryotrocha diadema. Toxics 2023, 11, 501. https://doi.org/10.3390/toxics11060501
Schleicherová D, Prearo M, Di Nunno C, Santovito A. Effects of Glyphosate on Female Reproductive Output in the Marine Polychaete Worm Ophryotrocha diadema. Toxics. 2023; 11(6):501. https://doi.org/10.3390/toxics11060501
Chicago/Turabian StyleSchleicherová, Dáša, Marino Prearo, Crystal Di Nunno, and Alfredo Santovito. 2023. "Effects of Glyphosate on Female Reproductive Output in the Marine Polychaete Worm Ophryotrocha diadema" Toxics 11, no. 6: 501. https://doi.org/10.3390/toxics11060501
APA StyleSchleicherová, D., Prearo, M., Di Nunno, C., & Santovito, A. (2023). Effects of Glyphosate on Female Reproductive Output in the Marine Polychaete Worm Ophryotrocha diadema. Toxics, 11(6), 501. https://doi.org/10.3390/toxics11060501