
Efficiency of Several Cytochrome P450 Biomarkers in Highlighting the Exposure of Daphnia magna to an Organophosphate Pesticide


Abstract
:1. Introduction
2. Materials and Methods
2.1. D. magna Culture Conditions
2.2. Chemical Exposure
2.3. Study Design
2.3.1. D. magna Exposed to Several Diazinon Concentrations
2.3.2. D. magna Exposed to Diazinon during Its First 5 Days of Life
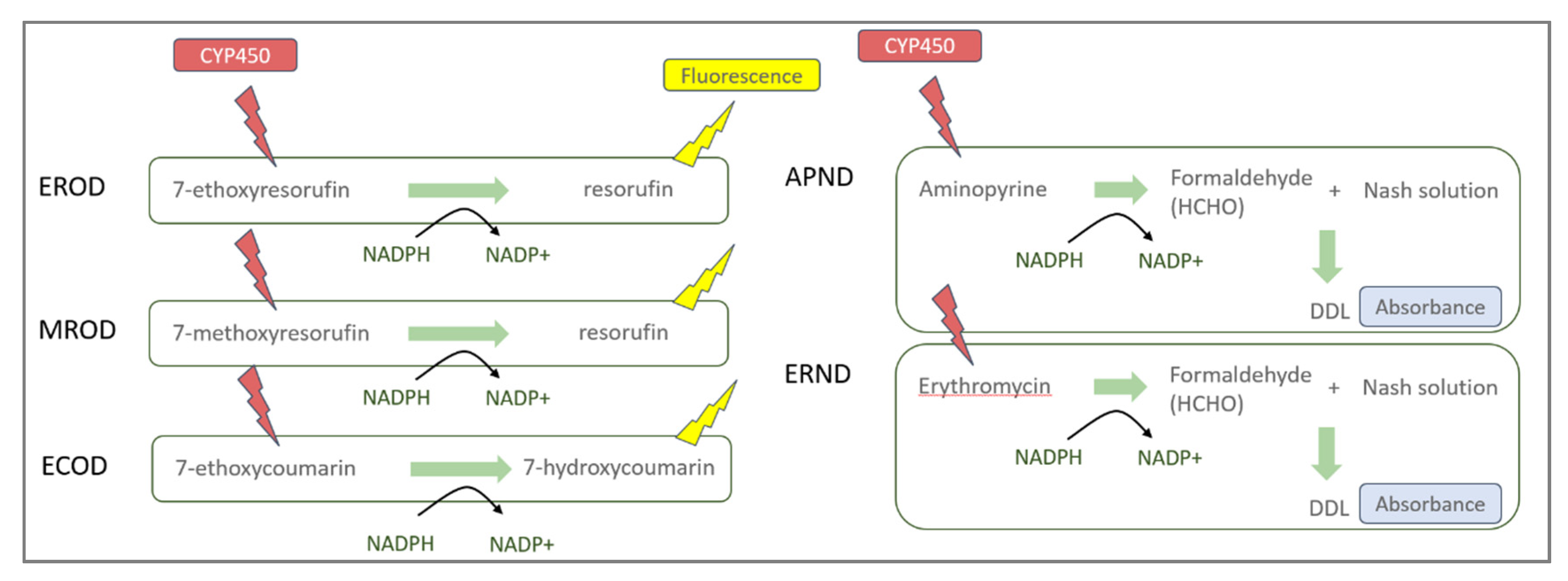
2.4. EROD, MROD, ECOD, APND, and ERND Activity Measurements
2.4.1. Protein Fraction Preparation
2.4.2. Protein Concentration
2.4.3. EROD, MROD, and ECOD Activities
2.4.4. APND and ERND Activities
2.5. Statistical Analysis
3. Results
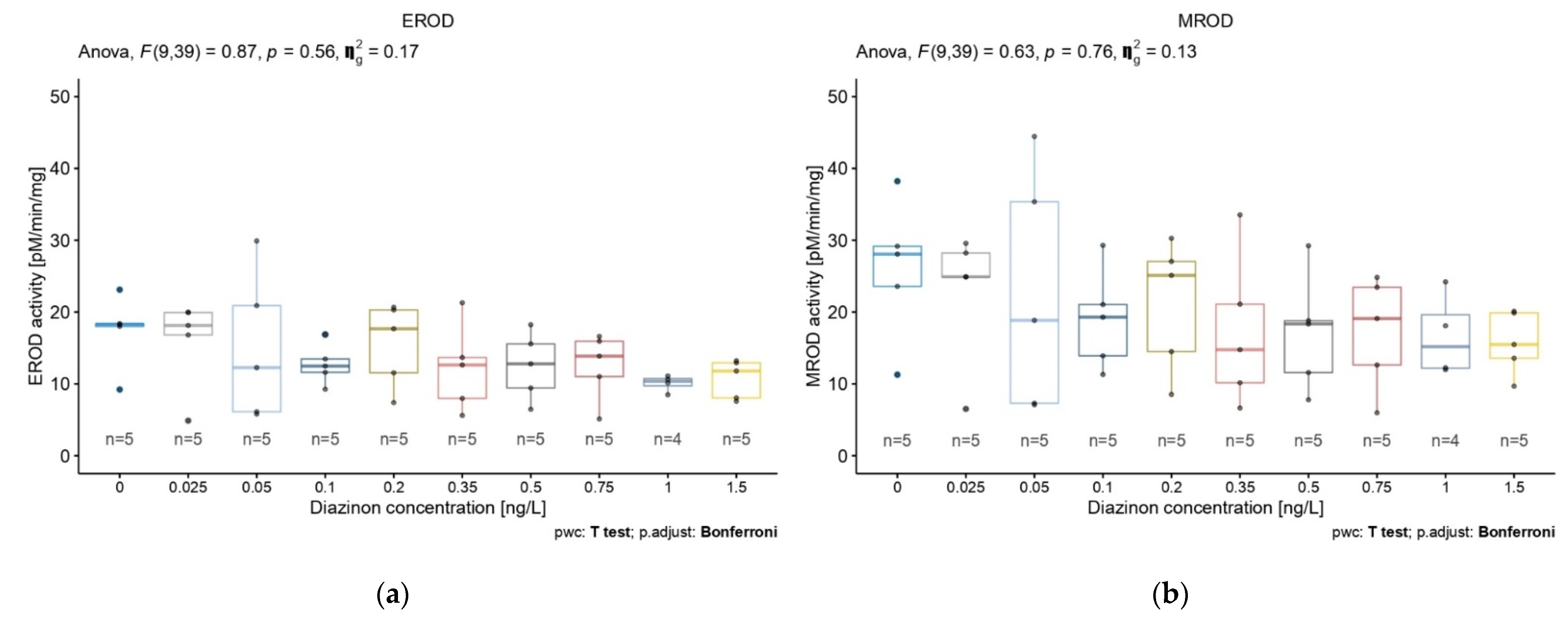
3.1. Exposure to Diazinon
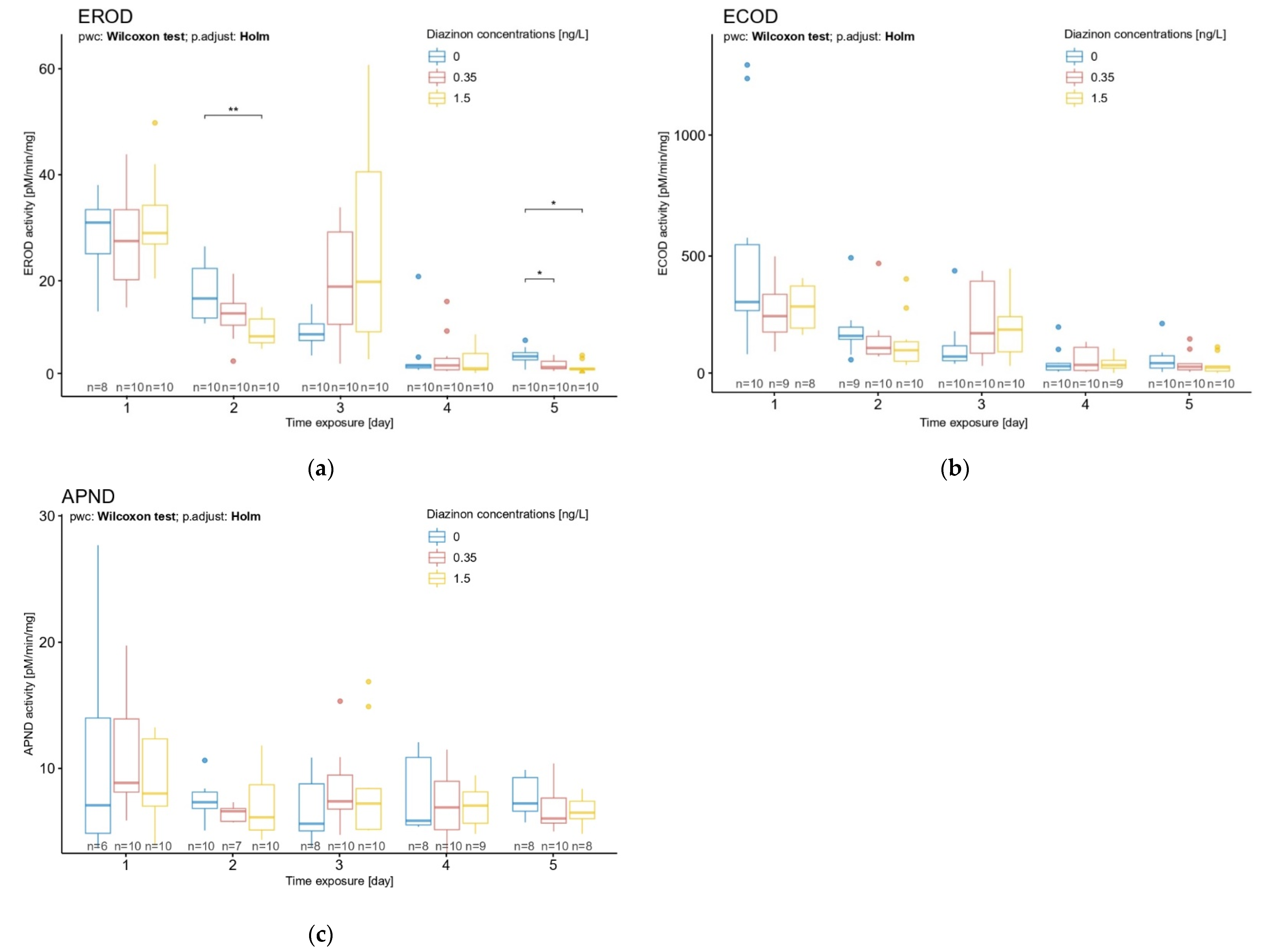
3.2. Diazinon Effect over Time on EROD, ECOD, and APND Biomarkers
3.2.1. Basal Activity of Biomarkers through the First 5 Days of D. magna Life
3.2.2. Time Exposure Effect on D. magna Exposed to Diazinon
4. Discussion
4.1. Detoxification Process Detection; CYP450 Biomarkers’ Optimization and Comparison
4.2. CYP450 Biomarkers’ Basal Activity
4.3. Effect of Diazinon through Time Exposure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Appendix A.1. List of Chemicals
Appendix A.2. EROD, MROD, and ECOD Stock Solution Details
Appendix A.3. ERND and APND Stock Solution Details
Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pairwise Wilcoxon Rank-Sum Test | ||
|---|---|---|
| D. magna Days Comparison | p | Statistic |
| 1 and 2 1 and 3 1 and 4 1 and 5 2 and 3 2 and 4 2 and 5 3 and 4 3 and 5 | 0.04 0.0009 0.0009 0.0005 0.007 0.005 0.0001 0.02 0.005 | 71 79 79 80 92 93 100 90 93 |
| Pairwise Wilcoxon Rank-Sum Test | ||
|---|---|---|
| D. magna Days Comparison | p | Statistic |
| 1 and 3 1 and 4 1 and 5 2 and 4 2 and 5 | 0.02 0.0008 0.0008 0.02 0.04 | 89 97 97 81 79 |
References
- Altshuler, I.; Demiri, B.; Xu, S.; Constantin, A.; Yan, N.D.; Cristescu, M.E. An Integrated Multi-Disciplinary Approach for Studying Multiple Stressors in Freshwater Ecosystems: Daphnia as a Model Organism. Integr. Comp. Biol. 2011, 51, 623–633. [Google Scholar] [CrossRef]
- Gunderson, M.P.; Pickett, M.A.; Martin, J.T.; Hulse, E.J.; Smith, S.S.; Smith, L.A.; Campbell, R.M.; Lowers, R.H.; Boggs, A.S.P.; Guillette, L.J. Variations in Hepatic Biomarkers in American Alligators (Alligator mississippiensis) from Three Sites in Florida, USA. Chemosphere 2016, 155, 180–187. [Google Scholar] [CrossRef]
- Guengerich, F.P. Cytochrome P450 and Chemical Toxicology. Chem. Res. Toxicol. 2008, 21, 70–83. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 Enzymes in Drug Metabolism: Regulation of Gene Expression, Enzyme Activities, and Impact of Genetic Variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Cedergreen, N. Quantifying Synergy: A Systematic Review of Mixture Toxicity Studies within Environmental Toxicology. PLoS ONE 2014, 9, e96580. [Google Scholar] [CrossRef]
- Komagata, O.; Kasai, S.; Tomita, T. Overexpression of Cytochrome P450 Genes in Pyrethroid-Resistant Culex Quinquefasciatus. Insect Biochem. Mol. Biol. 2010, 40, 146–152. [Google Scholar] [CrossRef]
- Yang, Y.; Chen, S.; Wu, S.; Yue, L.; Wu, Y. Constitutive Overexpression of Multiple Cytochrome P450 Genes Associated with Pyrethroid Resistance in Helicoverpa Armigera. J. Econ. Entomol. 2006, 99, 1784–1789. [Google Scholar] [CrossRef]
- Newman, M.C.; Unger, M.A. Fundamentals of Ecotoxicology; Ann Arbor Press: Chelsea, MI, USA, 1998; ISBN 978-1-57504-013-4. [Google Scholar]
- Timbrell, J.A. Factors Affecting Toxic Responses: Metabolism. In Principles of Biochemical Toxicology; CRC Press: Boca Raton, FL, USA, 2009; ISBN 978-0-429-12493-8. [Google Scholar]
- Kelly, S.L.; Kelly, D.E. Microbial Cytochromes P450: Biodiversity and Biotechnology. Where Do Cytochromes P450 Come from, What Do They Do and What Can They Do for Us? Philos. Trans. R. Soc. Lond. B Biol. Sci. 2013, 368, 20120476. [Google Scholar] [CrossRef]
- Xu, J.; Wang, X.; Guo, W. The Cytochrome P450 Superfamily: Key Players in Plant Development and Defense. J. Integr. Agric. 2015, 14, 1673–1686. [Google Scholar] [CrossRef]
- Hartl, M.G.J.; Kilemade, M.; Sheehan, D.; Mothersill, C.; O’Halloran, J.; O’Brien, N.M.; van Pelt, F.N.A.M. Hepatic Biomarkers of Sediment-Associated Pollution in Juvenile Turbot, Scophthalmus maximus L. Mar. Environ. Res. 2007, 64, 191–208. [Google Scholar] [CrossRef]
- Gunderson, M.P.; Oberdörster, E.; Guillette, L.J. Phase I and II Liver Enzyme Activities in Juvenile Alligators (Alligator mississippiensis) Collected from Three Sites in the Kissimmee-Everglades Drainage, Florida (USA). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2004, 139, 39–46. [Google Scholar] [CrossRef]
- Feyereisen, R. Insect P450 Enzymes. Annu. Rev. Entomol. 1999, 44, 507–533. [Google Scholar] [CrossRef]
- Gagnaire, B.; Geffard, O.; Noury, P.; Garric, J. In Vivo Indirect Measurement of Cytochrome P450-Associated Activities in Freshwater Gastropod Molluscs. Environ. Toxicol. 2010, 25, 545–553. [Google Scholar] [CrossRef]
- Gunderson, M.P.; Nguyen, B.T.; Cervantes Reyes, J.C.; Holden, L.L.; French, J.M.T.; Smith, B.D.; Lineberger, C. Response of Phase I and II Detoxification Enzymes, Glutathione, Metallothionein and Acetylcholine Esterase to Mercury and Dimethoate in Signal Crayfish (Pacifastacus leniusculus). Chemosphere 2018, 208, 749–756. [Google Scholar] [CrossRef]
- Dejong, C.A.; Wilson, J.Y. The Cytochrome P450 Superfamily Complement (CYPome) in the Annelid Capitella Teleta. PLoS ONE 2014, 9, e107728. [Google Scholar] [CrossRef]
- Koenig, S.; Fernández, P.; Solé, M. Differences in Cytochrome P450 Enzyme Activities between Fish and Crustacea: Relationship with the Bioaccumulation Patterns of Polychlorobiphenyls (PCBs). Aquat. Toxicol. 2012, 108, 11–17. [Google Scholar] [CrossRef]
- Lionetto, M.G.; Caricato, R.; Giordano, M.E. Pollution Biomarkers in the Framework of Marine Biodiversity Conservation: State of Art and Perspectives. Water 2021, 13, 1847. [Google Scholar] [CrossRef]
- Li, H.; Boon, J.P.; Lewis, W.E.; van den Berg, M.; Nyman, M.; Letcher, R.J. Hepatic Microsomal Cytochrome P450 Enzyme Activity in Relation to in Vitro Metabolism/Inhibition of Polychlorinated Biphenyls and Testosterone in Baltic Grey Seal (Halichoerus grypus). Environ. Toxicol. Chem. 2003, 22, 636–644. [Google Scholar] [CrossRef]
- Gottardi, M.; Kretschmann, A.; Cedergreen, N. Measuring Cytochrome P450 Activity in Aquatic Invertebrates: A Critical Evaluation of in Vitro and in Vivo Methods. Ecotoxicology 2016, 25, 419–430. [Google Scholar] [CrossRef]
- Kutlu, M.; Susuz, F. Effects of Lead as an Environmental Pollutant on EROD Enzyme in Gammarus pulex (L.) (Crustacea: Amphipoda). Bull. Environ. Contam. Toxicol. 2004, 72, 750–755. [Google Scholar] [CrossRef]
- Peng, Y.; Luo, Y.; Nie, X.-P.; Liao, W.; Yang, Y.-F.; Ying, G.-G. Toxic Effects of Triclosan on the Detoxification System and Breeding of Daphnia Magna. Ecotoxicology 2013, 22, 1384–1394. [Google Scholar] [CrossRef]
- Wang, L.; Peng, Y.; Nie, X.; Pan, B.; Ku, P.; Bao, S. Gene Response of CYP360A, CYP314, and GST and Whole-Organism Changes in Daphnia Magna Exposed to Ibuprofen. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2016, 179, 49–56. [Google Scholar] [CrossRef]
- Waxman, D.J.; Chang, T.K. Use of 7-Ethoxycoumarin to Monitor Multiple Enzymes in the Human CYP1, CYP2, and CYP3 Families. Methods Mol. Biol. 1998, 107, 175–179. [Google Scholar] [CrossRef]
- Dong, X.; Zhu, L.; Wang, J.; Wang, J.; Xie, H.; Hou, X.; Jia, W. Effects of Atrazine on Cytochrome P450 Enzymes of Zebrafish (Danio rerio). Chemosphere 2009, 77, 404–412. [Google Scholar] [CrossRef]
- Kristofco, L.A.; Cruz, L.C.; Haddad, S.P.; Behra, M.L.; Chambliss, C.K.; Brooks, B.W. Age Matters: Developmental Stage of Danio rerio Larvae Influences Photomotor Response Thresholds to Diazinion or Diphenhydramine. Aquat. Toxicol. 2016, 170, 344–354. [Google Scholar] [CrossRef]
- Sleiderink, H.M.; Oostingh, I.; Goksøyr, A.; Boon, J.P. Sensitivity of Cytochrome P450 1A Induction in Dab (Limanda limanda) of Different Age and Sex as a Biomarker for Environmental Contaminants in the Southern North Sea. Arch. Environ. Contam. Toxicol. 1995, 28, 423–430. [Google Scholar] [CrossRef]
- Bownik, A. Daphnia Swimming Behaviour as a Biomarker in Toxicity Assessment: A Review. Sci Total Environ. 2017, 601–602, 194–205. [Google Scholar] [CrossRef]
- Hebert, P.D.N. The Population Bilogy of Daphnia (Crustacea, Daphnidae). Biol. Rev. 1978, 53, 387–426. [Google Scholar] [CrossRef]
- Lampert, W. Daphnia: Model Herbivore, Predator and Prey. Pol. J. Ecol. 2006, 54, 607–620. [Google Scholar]
- Anderson, B.G. The Toxicity Thresholds of Various Substances Found in Industrial Wastes as Determined by the Use of Daphnia Magna. Sew. Work. J. 1944, 16, 1156–1165. [Google Scholar]
- Snyder, M.J. Cytochrome P450 Enzymes in Aquatic Invertebrates: Recent Advances and Future Directions. Aquat. Toxicol. 2000, 48, 529–547. [Google Scholar] [CrossRef]
- Livingstone, D.R. The Fate of Organic Xenobiotics in Aquatic Ecosystems: Quantitative and Qualitative Differences in Biotransformation by Invertebrates and Fish. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 1998, 120, 43–49. [Google Scholar] [CrossRef]
- Kretschmann, A.; Ashauer, R.; Preuss, T.G.; Spaak, P.; Escher, B.I.; Hollender, J. Toxicokinetic Model Describing Bioconcentration and Biotransformation of Diazinon in Daphnia Magna. Environ. Sci. Technol. 2011, 45, 4995–5002. [Google Scholar] [CrossRef]
- Mutch, E.; Williams, F.M. Diazinon, Chlorpyrifos and Parathion Are Metabolised by Multiple Cytochromes P450 in Human Liver. Toxicology 2006, 224, 22–32. [Google Scholar] [CrossRef]
- Sánchez, M.; Ferrando, M.D.; Sancho, E.; Andreu, E. Assessment of the Toxicity of a Pesticide with a Two-Generation Reproduction Test Using Daphnia Magna. Comp. Biochem. Physiol. C Pharmacol. Toxicol. Endocrinol. 1999, 124, 247–252. [Google Scholar] [CrossRef]
- OECD. Test No. 211: Daphnia Magna Reproduction Test; Organisation for Economic Co-Operation and Development: Paris, France, 2012. [Google Scholar]
- Jemec, A.; Drobne, D.; Tisler, T.; Trebse, P.; Ros, M.; Sepcić, K. The Applicability of Acetylcholinesterase and Glutathione S-Transferase in Daphnia Magna Toxicity Test. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 144, 303–309. [Google Scholar] [CrossRef]
- Afnor Mesurages Biochimiques et Physiologiques Sur Poisson—Partie 2: Dosage de l’éthoxyrésorufine-O-dééthylase (EROD). XP ISO/TS 23893-2. Normes Nationales et Documents Normatifs Nationaux. 2007; ICS code 13.060.70.
- Noury, P. Irstea Dosage de l’activité Ethoxyrésorufine-O-Dééthylase (EROD) Sur Microplate; HAL INRAE: Paris, France, 2016. [Google Scholar]
- Nash, T. The Colorimetric Estimation of Formaldehyde by Means of the Hantzsch Reaction. Biochem J. 1953, 55, 416–421. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Daphnia Genomics. Available online: http://wfleabase.org/ (accessed on 14 July 2022).
- Lee, B.-Y.; Choi, B.-S.; Kim, M.-S.; Park, J.C.; Jeong, C.-B.; Han, J.; Lee, J.-S. The Genome of the Freshwater Water Flea Daphnia Magna: A Potential Use for Freshwater Molecular Ecotoxicology. Aquat. Toxicol. 2019, 210, 69–84. [Google Scholar] [CrossRef]
- Religia, P.; Nguyen, N.D.; Nong, Q.D.; Matsuura, T.; Kato, Y.; Watanabe, H. Mutation of the Cytochrome P450 CYP360A8 Gene Increases Sensitivity to Paraquat in Daphnia Magna. Environ. Toxicol. Chem. 2021, 40, 1279–1288. [Google Scholar] [CrossRef]
- Kim, R.-O.; Jo, M.-A.; Song, J.; Kim, I.-C.; Yoon, S.; Kim, W.-K. Novel Approach for Evaluating Pharmaceuticals Toxicity Using Daphnia Model: Analysis of the Mode of Cytochrome P450-Generated Metabolite Action after Acetaminophen Exposure. Aquat. Toxicol. 2018, 196, 35–42. [Google Scholar] [CrossRef]
- Waxman, D.J.; Chang, T.K.H. Spectrofluorometric Analysis of CYP2A6-Catalyzed Coumarin 7-Hydroxylation. Methods Mol. Biol. 2006, 320, 91–96. [Google Scholar] [CrossRef]
- Poulsen, R.; De Fine Licht, H.H.; Hansen, M.; Cedergreen, N. Grandmother’s Pesticide Exposure Revealed Bi-Generational Effects in Daphnia Magna. Aquat. Toxicol. 2021, 236, 105861. [Google Scholar] [CrossRef]
- Roberts, B.J.; Shoaf, S.E.; Song, B.J. Rapid Changes in Cytochrome P4502E1 (CYP2E1) Activity and Other P450 Isozymes Following Ethanol Withdrawal in Rats. Biochem. Pharmacol. 1995, 49, 1665–1673. [Google Scholar] [CrossRef]
- Vaccaro, E.; Giorgi, M.; Longo, V.; Mengozzi, G.; Gervasi, P.G. Inhibition of Cytochrome P450 Enzymes by Enrofloxacin in the Sea Bass (Dicentrarchus Labrax). Aquat. Toxicol. 2003, 62, 27–33. [Google Scholar] [CrossRef]
- Borbás, T.; Benko, B.; Dalmadi, B.; Szabó, I.; Tihanyi, K. Insulin in Flavin-Containing Monooxygenase Regulation. Flavin-Containing Monooxygenase and Cytochrome P450 Activities in Experimental Diabetes. Eur. J. Pharm. Sci. 2006, 28, 51–58. [Google Scholar] [CrossRef]
- Sved, D.W.; Van Veld, P.A.; Roberts, M.H. Hepatic EROD Activity in Spot, Leiostomus Xanthurus, Exposed to Creosote-Contaminated Sediments. Mar. Environ. Res. 1992, 34, 189–193. [Google Scholar] [CrossRef]
- Martínez-Jerónimo, F. Description of the Individual Growth of Daphnia Magna (Crustacea: Cladocera) through the von Bertalanffy Growth Equation. Effect of Photoperiod and Temperature. Limnology 2012, 13, 65–71. [Google Scholar] [CrossRef]
- Rossiter MaryCarol. Incidence and Consequences of Inherited Environmental Effects. Annu. Rev. Ecol. Syst. 1996, 27, 451–476. [Google Scholar] [CrossRef]
- Frost, P.C.; Ebert, D.; Larson, J.H.; Marcus, M.A.; Wagner, N.D.; Zalewski, A. Transgenerational Effects of Poor Elemental Food Quality on Daphnia Magna. Oecologia 2010, 162, 865–872. [Google Scholar] [CrossRef]
- Boersma, M. Offspring Size in Daphnia: Does It Pay to Be Overweight? In Cladocera: The Biology of Model Organisms; Springer: Dordrecht, The Netherlands, 1997; Volume 360, pp. 79–88. [Google Scholar] [CrossRef]
- dos Santos, K.C.; Martinez, C.B.R. Genotoxic and Biochemical Effects of Atrazine and Roundup(®), Alone and in Combination, on the Asian Clam Corbicula Fluminea. Ecotoxicol. Environ. Saf. 2014, 100, 7–14. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo de Almeida, E.; Tisserand, F.; Faria, M.; Chèvre, N. Efficiency of Several Cytochrome P450 Biomarkers in Highlighting the Exposure of Daphnia magna to an Organophosphate Pesticide. Toxics 2022, 10, 482. https://doi.org/10.3390/toxics10080482
Melo de Almeida E, Tisserand F, Faria M, Chèvre N. Efficiency of Several Cytochrome P450 Biomarkers in Highlighting the Exposure of Daphnia magna to an Organophosphate Pesticide. Toxics. 2022; 10(8):482. https://doi.org/10.3390/toxics10080482
Chicago/Turabian StyleMelo de Almeida, Elodie, Floriane Tisserand, Micaela Faria, and Nathalie Chèvre. 2022. "Efficiency of Several Cytochrome P450 Biomarkers in Highlighting the Exposure of Daphnia magna to an Organophosphate Pesticide" Toxics 10, no. 8: 482. https://doi.org/10.3390/toxics10080482
APA StyleMelo de Almeida, E., Tisserand, F., Faria, M., & Chèvre, N. (2022). Efficiency of Several Cytochrome P450 Biomarkers in Highlighting the Exposure of Daphnia magna to an Organophosphate Pesticide. Toxics, 10(8), 482. https://doi.org/10.3390/toxics10080482