


Detection of Changes in Monoamine Neurotransmitters by the Neonicotinoid Pesticide Imidacloprid Using Mass Spectrometry

 , , ,
, , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Animals
2.3. IMI Administration, Euthanasia, Dissection, and Organ Collection
2.3.1. For the Behavioral Test
2.3.2. For the Monoamine Concentration Measurement
2.4. Behavioral Test
2.4.1. Open Field Test (OF)
2.4.2. Elevated Plus Maze Test (EPM)
2.5. Investigation of the Conditions for DPP Derivatization and Analysis
2.5.1. Target Substances
2.5.2. LC/MS Measurement Conditions
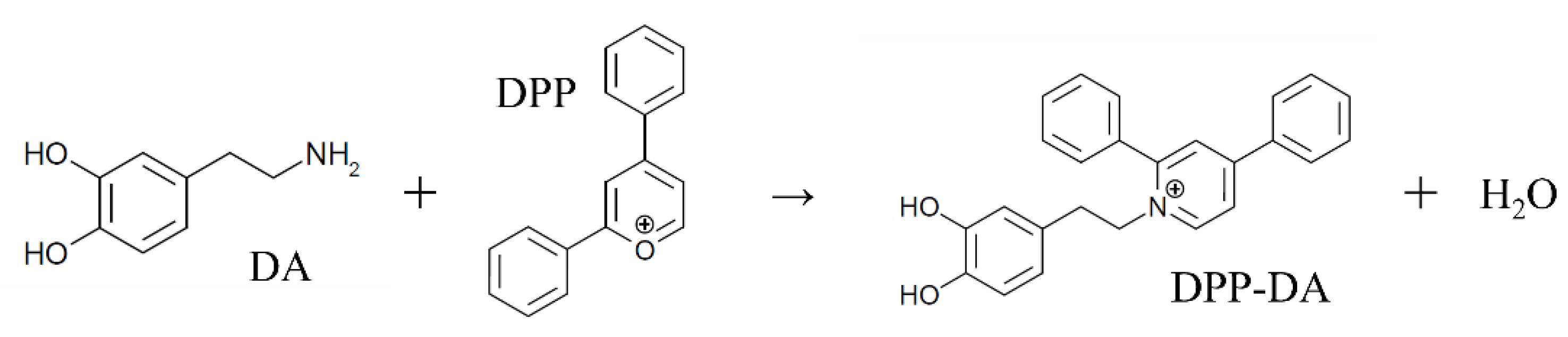
2.5.3. Investigation of Derivatization Reaction Methods
Preparation of Standard (STD) Mix and Internal Standard (IS) Mix
Preparation of the DPP Mix
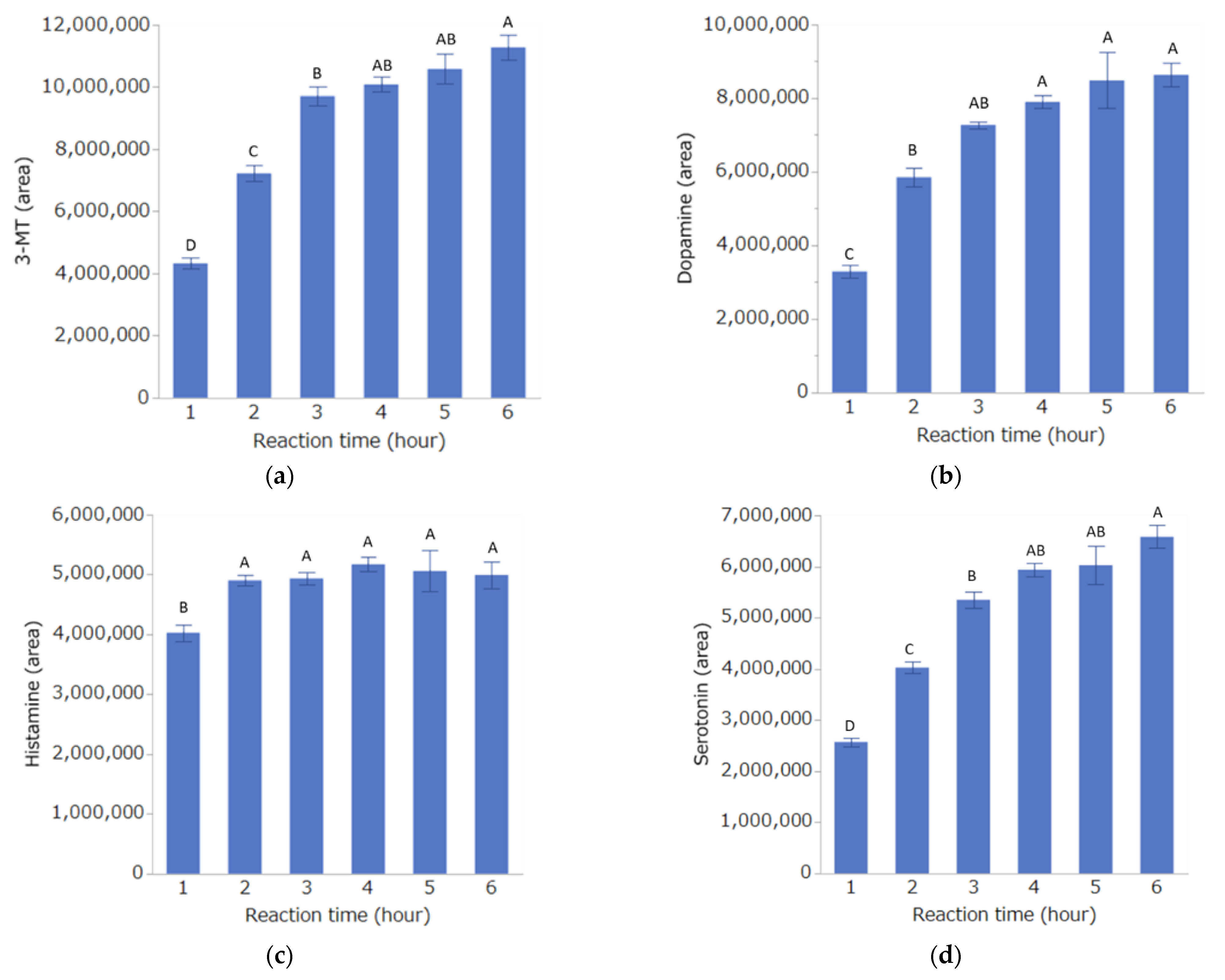
Investigation of Derivatization Reaction Time
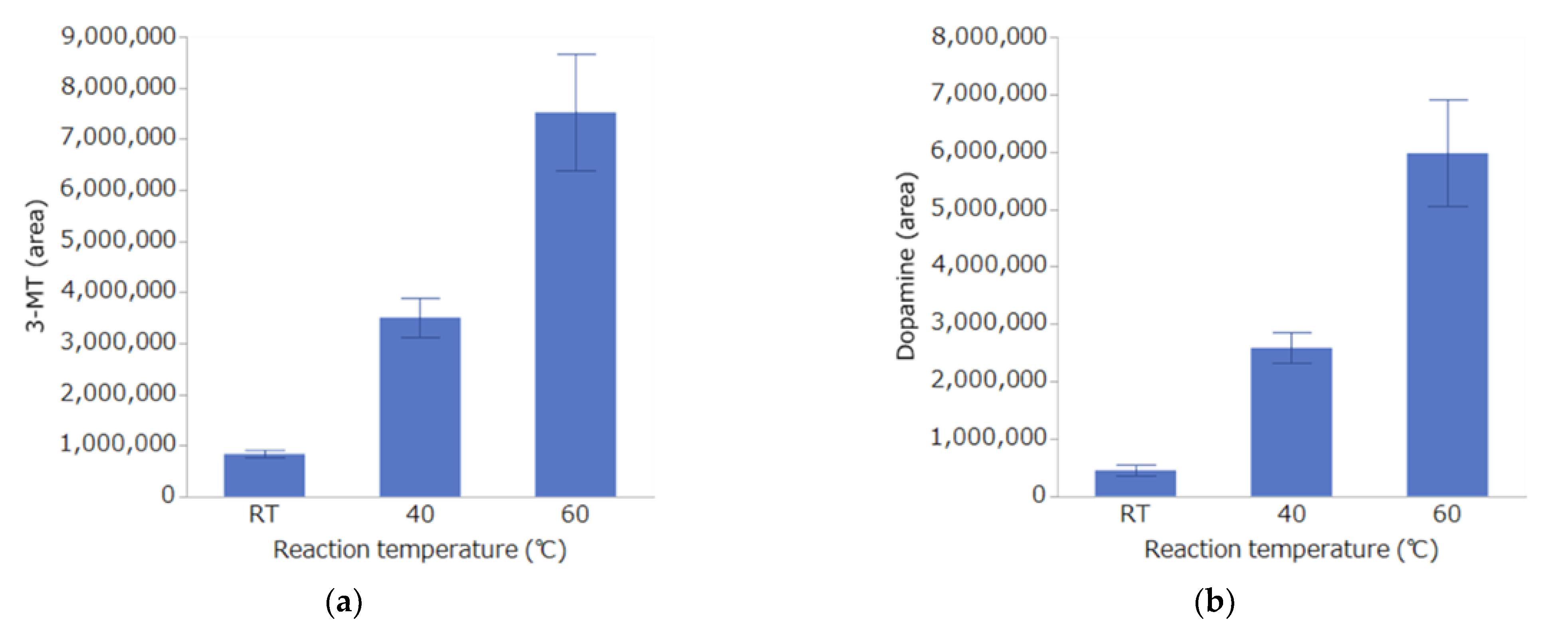
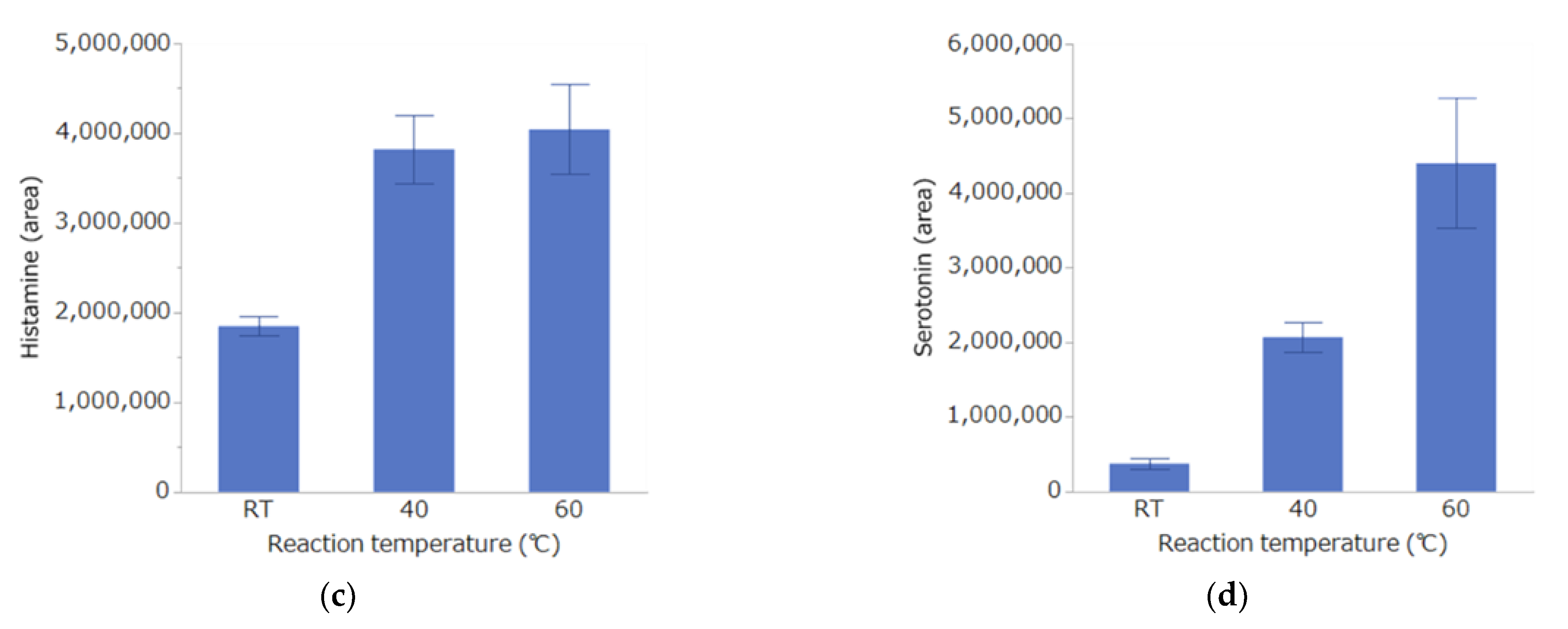
Investigation of Derivatization Reaction Temperature
Calculation of Instrument Detection Limit (IDL) and Method Detection Limit (MDL)
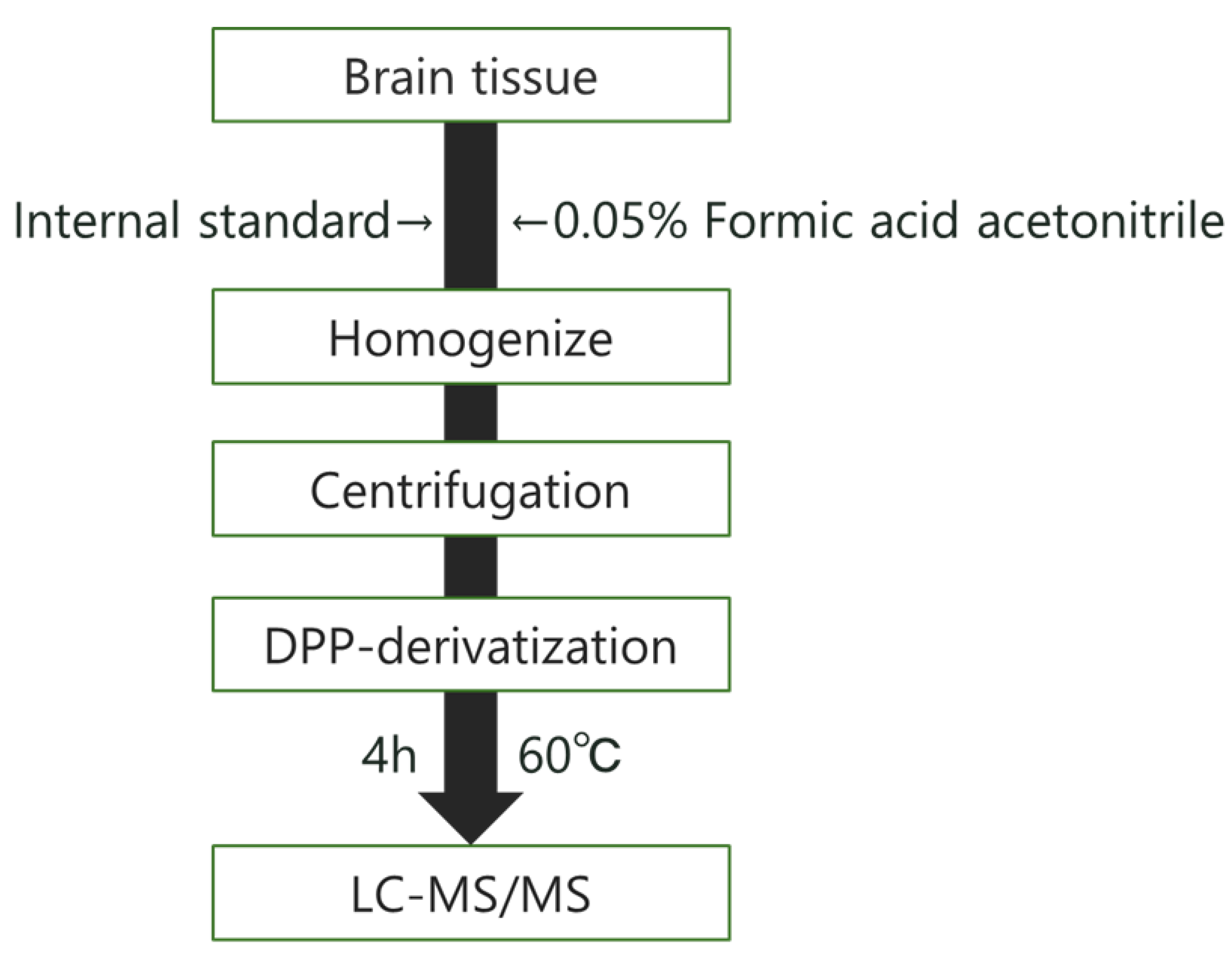
2.6. Measurement of Monoamine Concentration
2.6.1. Measurement of Monoamine Concentration in Brain Tissue
2.6.2. Measurement of Plasma Monoamine Concentration
2.7. Statistical Analysis
3. Results
3.1. Investigation of Conditions for DPP Derivatization Used in the Analysis Method
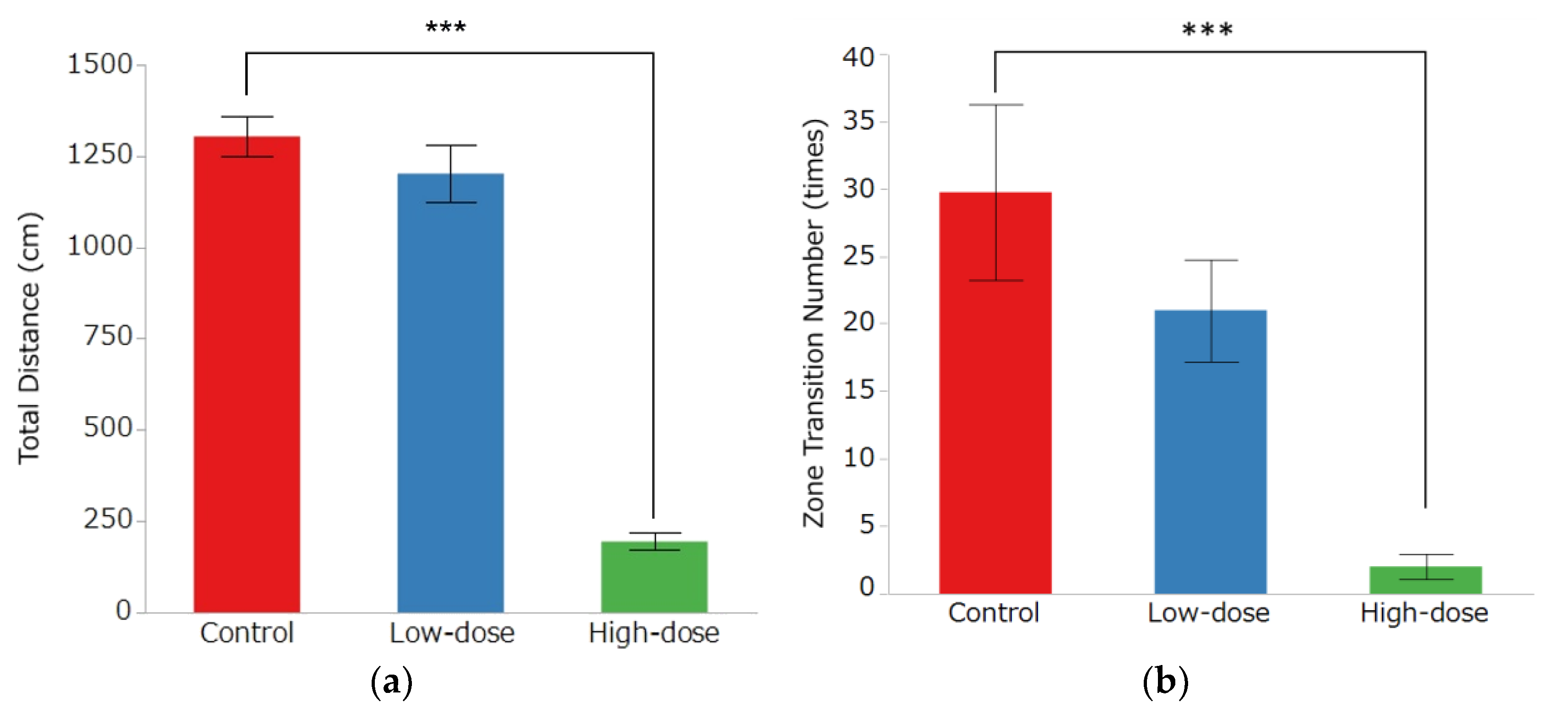
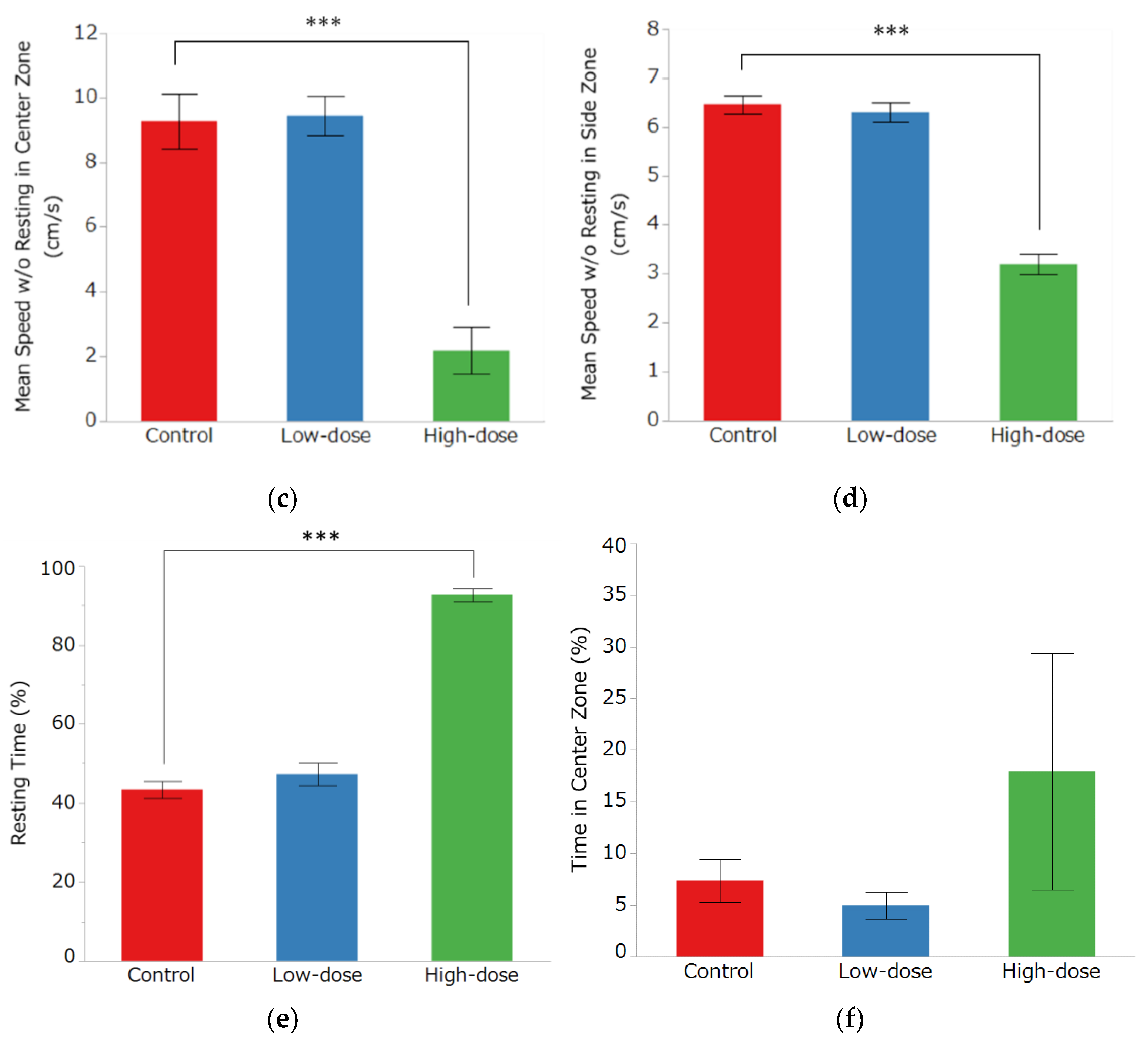
3.2. Open Field Test
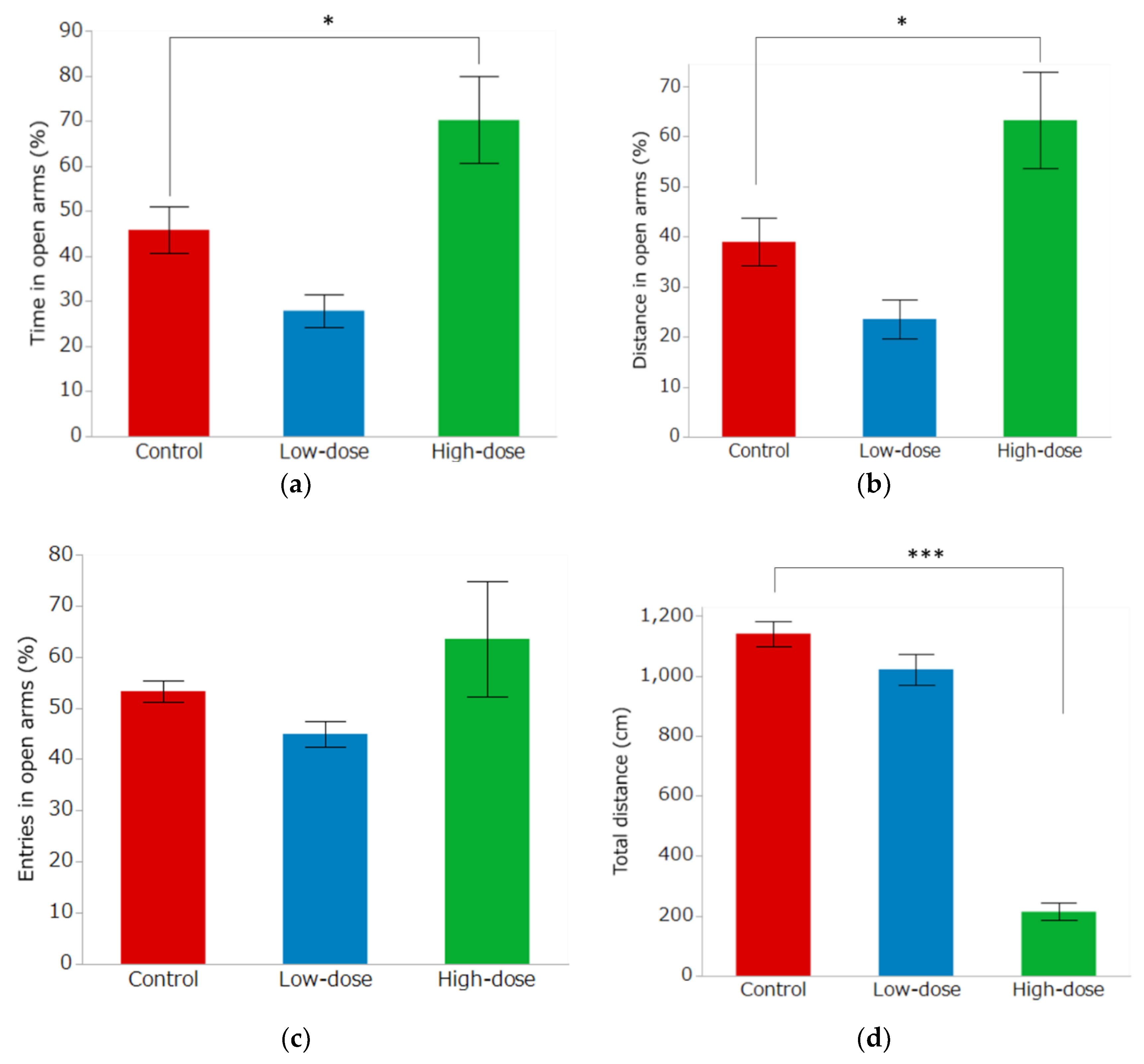
3.3. Elevated Plus Maze Test
3.4. Measurement of Monoamine Concentrations in Various Parts of the Brain
3.5. Measurement of Plasma Monoamine Concentration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bass, C.; Denholm, I.; Williamson, M.S.; Nauen, R. The global status of insect resistance to neonicotinoid insecticides. Pestic. Biochem. Physiol. 2015, 121, 78–87. [Google Scholar] [CrossRef]
- Jeschke, P.; Nauen, R.; Schindler, M.; Elbert, A. Overview of the status and global strategy for neonicotinoids. J. Agric. Food Chem. 2011, 59, 2897–2908. [Google Scholar] [CrossRef] [PubMed]
- Taillebois, E.; Cartereau, A.; Jones, A.K.; Thany, S.H. Neonicotinoid insecticides mode of action on insect nicotinic acetylcholine receptors using binding studies. Pestic. Biochem. Physiol. 2018, 151, 59–66. [Google Scholar] [CrossRef]
- Tomizawa, M.; Lee, D.L.; Casida, J.E. Neonicotinoid insecticides: Molecular features conferring selectivity for insect versus mammalian nicotinic receptors. J. Agric. Food Chem. 2000, 48, 6016–6024. [Google Scholar] [CrossRef] [PubMed]
- Lansdell, S.J.; Millar, N.S. The influence of nicotinic receptor subunit composition upon agonist, α-bungarotoxin and insecticide (imidacloprid) binding affinity. Neuropharmacology 2000, 39, 671–679. [Google Scholar] [CrossRef]
- Matsuda, K.; Buckingham, S.D.; Kleier, D.; Rauh, J.J.; Grauso, M.; Sattelle, D.B.; Buckingham, S.D. Neonicotinoids: Insecticides actingon insect nicotinic acetylcholinereceptors. J. Agric. Food Chem. 2001, 22, 573–580. [Google Scholar] [CrossRef]
- Tomizawa, M.; Casida, J.E. Selective Toxicity of Neonicotinoids Attributable to Specificity of Insect and Mammalian Nicotinic Receptors. Annu. Rev. Entomol. 2003, 48, 339–364. [Google Scholar] [CrossRef] [PubMed]
- Tomizawa, M.; Casida, J.E. Neonicotinoid insecticide toxicology: Mechanisms of selective action. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 247–268. [Google Scholar] [CrossRef]
- Lonare, M.; Kumar, M.; Raut, S.; Badgujar, P.; Doltade, S.; Telang, A. Evaluation of imidacloprid-induced neurotoxicity in male rats: A protective effect of curcumin. Neurochem. Int. 2014, 78, 122–129. [Google Scholar] [CrossRef]
- Zheng, M.; Qin, Q.; Zhou, W.; Liu, Q.; Zeng, S.; Xiao, H.; Bai, Q.; Gao, J. Metabolic disturbance in hippocampus and liver of mice: A primary response to imidacloprid exposure. Sci. Rep. 2020, 10, 5713. [Google Scholar] [CrossRef]
- Gu, Y.h.; Li, Y.; Huang, X.f.; Zheng, J.f.; Yang, J.; Diao, H.; Yuan, Y.; Xu, Y.; Liu, M.; Shi, H.j.; et al. Reproductive Effects of Two Neonicotinoid Insecticides on Mouse Sperm Function and Early Embryonic Development In Vitro. PLoS ONE 2013, 8, e70112. [Google Scholar] [CrossRef] [PubMed]
- Mohany, M.; Badr, G.; Refaat, I.; El-Feki, M. Immunological and histological effects of exposure to imidacloprid insecticide in male albino rats. African J. Pharm. Pharmacol. 2011, 5, 2106–2114. [Google Scholar] [CrossRef]
- Toor, H.K.; Sangha, G.K.; Khera, K.S. Imidacloprid induced histological and biochemical alterations in liver of female albino rats. Pestic. Biochem. Physiol. 2013, 105, 1–4. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, I.M.; Nunes, B.V.F.; Barbosa, D.R.; Pallares, A.M.; Faro, L.R.F. Effects of the neonicotinoids thiametoxam and clothianidin on in vivo dopamine release in rat striatum. Toxicol. Lett. 2010, 192, 294–297. [Google Scholar] [CrossRef]
- Nieoullon, A.; Coquerel, A. Dopamine: A key regulator to adapt action, emotion, motivation and cognition. Curr. Opin. Neurol. 2003, 16, 3–9. [Google Scholar] [CrossRef]
- Bhardwaj, S.; Srivastava, M.K.; Kapoor, U.; Srivastava, L.P. A 90 days oral toxicity of imidacloprid in female rats: Morphological, biochemical and histopathological evaluations. Food Chem. Toxicol. 2010, 48, 1185–1190. [Google Scholar] [CrossRef]
- Rodrigues, K.J.A.; Santana, M.B.; Nascimento, J.L.M.D.; Picanço-Diniz, D.L.W.; Maués, L.A.L.; Santos, S.N.; Ferreira, V.M.M.; Alfonso, M.; Durán, R.; Faro, L.R.F. Behavioral and biochemical effects of neonicotinoid thiamethoxam on the cholinergic system in rats. Ecotoxicol. Environ. Saf. 2010, 73, 101–107. [Google Scholar] [CrossRef]
- Hirano, T.; Yanai, S.; Takada, T.; Yoneda, N.; Omotehara, T.; Kubota, N.; Minami, K.; Yamamoto, A.; Mantani, Y.; Yokoyama, T.; et al. NOAEL-dose of a neonicotinoid pesticide, clothianidin, acutely induce anxiety-related behavior with human-audible vocalizations in male mice in a novel environment. Toxicol. Lett. 2018, 282, 57–63. [Google Scholar] [CrossRef]
- Yoneda, N.; Takada, T.; Hirano, T.; Yanai, S.; Yamamoto, A.; Mantani, Y.; Yokoyama, T.; Kitagawa, H.; Tabuchi, Y.; Hoshi, N. Peripubertal exposure to the neonicotinoid pesticide dinotefuran affects dopaminergic neurons and causes hyperactivity in male mice. J. Vet. Med. Sci. 2018, 80, 634–637. [Google Scholar] [CrossRef]
- Hirano, T.; Yanai, S.; Omotehara, T.; Hashimoto, R.; Umemura, Y.; Kubota, N.; Minami, K.; Nagahara, D.; Matsuo, E.; Aihara, Y.; et al. The combined effect of clothianidin and environmental stress on the behavioral and reproductive function in male mice. J. Vet. Med. Sci. 2015, 77, 1207–1215. [Google Scholar] [CrossRef]
- Abd-Elhakim, Y.M.; Mohammed, H.H.; Mohamed, W.A.M. Imidacloprid Impacts on Neurobehavioral Performance, Oxidative Stress, and Apoptotic Events in the Brain of Adolescent and Adult Rats. J. Agric. Food Chem. 2018, 66, 13513–13524. [Google Scholar] [CrossRef] [PubMed]
- Kubo, S.; Hirano, T.; Miyata, Y.; Ohno, S.; Onaru, K.; Ikenaka, Y.; Nakayama, S.M.M.; Ishizuka, M.; Mantani, Y.; Yokoyama, T.; et al. Sex-specific behavioral effects of acute exposure to the neonicotinoid clothianidin in mice. Toxicol. Appl. Pharmacol. 2022, 456, 116283. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.; Cañadas, F.; Cardona, D.; Suñol, C.; Campa, L.; Sánchez-Amate, M.C.; Flores, P.; Sanchez-Santed, F. Long-term monoamine changes in the striatum and nucleus accumbens after acute chlorpyrifos exposure. Toxicol. Lett. 2008, 176, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Kanno, J. Introduction to the concept of signal toxicity. J. Toxicol. Sci. 2016, 41, SP105–SP109. [Google Scholar] [CrossRef]
- Yoshimura, S. Neurotoxicity test. Pharm. Soc. Japan. 1993, 29, 35–37. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Fu, Q.; Leung, K.W.; Wong, Z.C.F.; Choi, R.C.Y.; Tsim, K.W.K. The establishment of a sensitive method in determining different neurotransmitters simultaneously in rat brains by using liquid chromatography-electrospray tandem mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2011, 879, 737–742. [Google Scholar] [CrossRef] [PubMed]
- Mabrouk, O.S.; Li, Q.; Song, P.; Kennedy, R.T. Microdialysis and mass spectrometric monitoring of dopamine and enkephalins in the globus pallidus reveal reciprocal interactions that regulate movement. J. Neurochem. 2011, 118, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.X.; Xian, Y.Y.; Liu, S.; Yu, F.; Mu, H.J.; Sun, K.X.; Liu, W.H. Development, validation and comparison of surrogate matrix and surrogate analyte approaches with UHPLC–MS/MS to simultaneously quantify dopamine, serotonin and γ-aminobutyric acid in four rat brain regions. Biomed. Chromatogr. 2018, 32, e4276. [Google Scholar] [CrossRef]
- Sørensen, L.K.; Johannsen, M. Sensitive determination of monoamine neurotransmitters, their main metabolites and precursor amino acids in different mouse brain components by liquid chromatography–electrospray tandem mass spectrometry after selective sample clean-up. Biomed. Chromatogr. 2019, 33, e4479. [Google Scholar] [CrossRef]
- Shariatgorji, M.; Nilsson, A.; Goodwin, R.J.A.; Källback, P.; Schintu, N.; Zhang, X.; Crossman, A.R.; Bezard, E.; Svenningsson, P.; Andren, P.E. Direct targeted quantitative molecular imaging of neurotransmitters in brain tissue sections. Neuron 2014, 84, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Shariatgorji, M.; Nilsson, A.; Källback, P.; Karlsson, O.; Zhang, X.; Svenningsson, P.; Andren, P.E. Pyrylium salts as reactive matrices for MALDI-MS imaging of biologically active primary amines. J. Am. Soc. Mass Spectrom. 2015, 26, 934–939. [Google Scholar] [CrossRef] [PubMed]
- Shimma, S.; Takeo, E.; Fukusaki, E. Protocol for quantitative imaging mass spectrometry. Bunseki Kagaku 2016, 65, 745–750. [Google Scholar] [CrossRef]
- Esteve, C.; Tolner, E.A.; Shyti, R.; van den Maagdenberg, A.M.J.M.; McDonnell, L.A. Mass spectrometry imaging of amino neurotransmitters: A comparison of derivatization methods and application in mouse brain tissue. Metabolomics 2016, 12, 30. [Google Scholar] [CrossRef]
- Biala, G.; Budzynska, B. Effects of acute and chronic nicotine on elevated plus maze in mice: Involvement of calcium channels. Life Sci. 2006, 79, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Imidacloprid Pesticide Abstracts (in Japanese), Bayer CropScience K.K. 2017. Available online: https://www.acis.famic.go.jp/syouroku/imidacloprid/index.htm (accessed on 25 February 2021).
- Bruhwyler, J. Anxiolytic potential of a microgram dose of chlordiazepoxide in the open-field test. Eur. J. Pharmacol. 1990, 187, 547–549. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Yoshioka, M. Evaluation for anxiety-related behaviors in rodents. Folia Pharmacol. Jpn. 2007, 130, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Togashi, H.; Matsumoto, M.; Yoshioka, M. Evaluation of anxiety-related behavior in elevated plus-maze test and its applications. Folia Pharmacol. Jpn. 2005, 126, 99–105. [Google Scholar] [CrossRef]
- Merdas, M.; Lagarrigue, M.; Vanbellingen, Q.; Umbdenstock, T.; da Violante, G.; Pineau, C. On-tissue chemical derivatization reagents for matrix-assisted laser desorption/ionization mass spectrometry imaging. J. Mass Spectrom. 2021, 56, e4731. [Google Scholar] [CrossRef]
- Maeda, N.; Sato, M.; Haeno, S.; Yokota, H. Simultaneous Determination of the Neurotransmitters and Free Amino Acids in Rat Organs by LC-MS Analysis. Sci. J. Grad. Sch. Rakuno Gakuen Univ. 2016, 2016, 1–7. [Google Scholar]
- Huang, F.; Li, J.; Shi, H.l.; Wang, T.t.; Muhtar, W.; Du, M.; Zhang, B.b.; Wu, H.; Yang, L.; Hu, Z.b.; et al. Simultaneous quantification of seven hippocampal neurotransmitters in depression mice by LC-MS/MS. J. Neurosci. Methods 2014, 229, 8–14. [Google Scholar] [CrossRef]
- Tareke, E.; Bowyer, J.F.; Doerge, D.R. Quantification of rat brain neurotransmitters and metabolites using liquid chromatography/electrospray tandem mass spectrometry and comparison with liquid chromatography/electrochemical detection. Rapid Commun. Mass Spectrom. 2007, 21, 3898–3904. [Google Scholar] [CrossRef] [PubMed]
- González, R.R.; Fernández, R.F.; Vidal, J.L.M.; Frenich, A.G.; Pérez, M.L.G. Development and validation of an ultra-high performance liquid chromatography-tandem mass-spectrometry (UHPLC-MS/MS) method for the simultaneous determination of neurotransmitters in rat brain samples. J. Neurosci. Methods 2011, 198, 187–194. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.H.; Choi, J.; Kim, H.G.; Kim, H.R. Quantification of neurotransmitters in mouse brain tissue by using liquid chromatography coupled electrospray tandem mass spectrometry. J. Anal. Methods Chem. 2014, 2014, 506870. [Google Scholar] [CrossRef] [PubMed]
- Hagino, Y.; Kasai, S.; Fujita, M.; Setogawa, S.; Yamaura, H.; Yanagihara, D.; Hashimoto, M.; Kobayashi, K.; Meltzer, H.Y.; Ikeda, K. Involvement of Cholinergic System in Hyperactivity in Dopamine-Deficient Mice. Neuropsychopharmacology 2015, 40, 1141–1150. [Google Scholar] [CrossRef]
- Prus, A.J.; Vann, R.E.; Rosecrans, J.A.; James, J.R.; Pehrson, A.L.; O’Connell, M.M.; Philibin, S.D.; Robinson, S.E. Acute nicotine reduces and repeated nicotine increases spontaneous activity in male and female Lewis rats. Pharmacol. Biochem. Behav. 2008, 91, 150–154. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akinola, L.S.; McKiver, B.; Toma, W.; Zhu, A.Z.X.; Tyndale, R.F.; Kumar, V.; Damaj, M.I. C57BL/6 substrain differences in pharmacological effects after acute and repeated nicotine administration. Brain Sci. 2019, 9, 244. [Google Scholar] [CrossRef]
- Robinson, S.E.; James, J.R.; Lapp, L.N.; Vann, R.E.; Gross, D.F.; Philibin, S.D.; Rosecrans, J.A. Evidence of cellular nicotinic receptor desensitization in rats exhibiting nicotine-induced acute tolerance. Psychopharmacology 2006, 184, 306–313. [Google Scholar] [CrossRef]
- Cachope, R.; Cheer, J.F. Local control of striatal dopamine release. Front. Behav. Neurosci. 2014, 8, 188. [Google Scholar] [CrossRef]
- Kimura-Kuroda, J.; Komuta, Y.; Kuroda, Y.; Hayashi, M.; Kawano, H. Nicotine-like effects of the neonicotinoid insecticides acetamiprid and imidacloprid on cerebellar neurons from neonatal rats. PLoS ONE 2012, 7, e32432. [Google Scholar] [CrossRef]
- Li, P.; Ann, J.; Akk, G. Activation and modulation of human α4β2 nicotinic acetylcholine receptors by the neonicotinoids clothianidin and imidacloprid. J. Neurosci. Res. 2011, 89, 1295–1301. [Google Scholar] [CrossRef]
- Pignatelli, A.; Kobayashi, K.; Okano, H.; Belluzzi, O. Functional properties of dopaminergic neurones in the mouse olfactory bulb. J. Physiol. 2005, 564, 501–514. [Google Scholar] [CrossRef] [PubMed]
- Doty, R.L. Olfaction in Parkinson’s disease and related disorders. Neurobiol. Dis. 2012, 46, 527–552. [Google Scholar] [CrossRef] [PubMed]
- Höglinger, G.U.; Alvarez-Fischer, D.; Arias-Carrión, O.; Djufri, M.; Windolph, A.; Keber, U.; Borta, A.; Ries, V.; Schwarting, R.K.W.; Scheller, D.; et al. A new dopaminergic nigro-olfactory projection. Acta Neuropathol. 2015, 130, 333–348. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, X.A.; Franco, A.; Fernández-Novoa, L.; Cacabelos, R. Blood levels of histamine, IL-1β, and TNF-α in patients with mild to moderate Alzheimer disease. Mol. Chem. Neuropathol. 1996, 29, 237–252. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Novoa, L.; Cacabelos, R. Histamine function in brain disorders. Behav. Brain Res. 2001, 124, 213–233. [Google Scholar] [CrossRef]
- Coelho, M.H.; Silva, I.J.; Azevedo, M.S.; Manso, C.F. Decrease in blood histamine in drug-treated Parkinsonian patients. Mol. Chem. Neuropathol. 1991, 14, 77–85. [Google Scholar] [CrossRef]
- Gholipoor, P.; Saboory, E.; Roshan-Milani, S.; Fereidoni, J. Effect of hyperthermia on histamine blood level and convulsive behavior in infant rats. Epilepsy Behav. 2013, 29, 269–274. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Brain Region | Control [mg] | Low-Dose [mg] | High-Dose [mg] |
|---|---|---|---|
| Striatum | 3.30 ± 0.20 | 3.09 ± 0.21 | 3.84 ± 0.23 |
| Hippocampus | 6.54 ± 0.55 | 5.84 ± 0.47 | 7.25 ± 0.46 |
| Cortex | 54.29 ± 5.18 | 62.36 ± 5.89 | 69.30 ± 1.04 |
| Cerebellum | 89.14 ± 5.27 | 95.61 ± 3.46 | 88.61 ± 2.08 |
| Brain stem | 65.90 ± 3.12 | 68.86 ± 1.46 | 68.70 ± 1.70 |
| Olfactory bulb | 20.15 ± 1.04 | 18.46 ± 1.68 | 18.69 ± 1.15 |
| Compound | RT [min] | MRM Transition [m/z] | CE [V] | |
|---|---|---|---|---|
| Precursor Ion | Product Ion | |||
| DPP-3-MT | 3.030 | 383.1 | 151.2 | 24 |
| 91.0 | 60 | |||
| DPP-Dopamine | 2.860 | 369.0 | 233.3 | 24 |
| 233.2 | 20 | |||
| DPP-Histamine | 2.060 | 327.0 | 95.1 | 32 |
| 233.4 | 28 | |||
| 68.1 | 72 | |||
| DPP-Serotonin | 2.930 | 392.0 | 160.0 | 32 |
| 232.3 | 20 | |||
| DPP-3-MT-d4 | 3.030 | 387.3 | 156.2 | 24 |
| 96.0 | 68 | |||
| DPP-Dopamine-d4 | 2.860 | 372.1 | 232.1 | 28 |
| 140.9 | 28 | |||
| 94.5 | 56 | |||
| DPP-Histamine-d4 | 2.060 | 330.0 | 232.2 | 24 |
| 99.1 | 36 | |||
| DPP-Serotonin-d4 | 2.930 | 395.0 | 164.2 | 24 |
| 232.1 | 16 | |||
| Compound | IDL [ppt] | MDL [ppt] |
|---|---|---|
| 3-MT | 0.90 | 2.08 |
| Dopamine | 4.48 | 24.9 |
| Histamine | 3.24 | 9.82 |
| Serotonin | 0.91 | 1.51 |
| Brain Region | Control | Low-Dose | High-Dose |
|---|---|---|---|
| 3-MT [ng/g] | |||
| Striatum | 747.3 ± 88.0 | 807.4 ± 137.1 | 595.6 ± 39.9 |
| Hippocampus | 6.1 ± 1.2 | 5.0 ± 0.6 | 4.9 ± 1.0 |
| Cortex | 66.7 ± 15.6 | 69.0 ± 15.8 | 56.5 ± 5.2 ☨ |
| Cerebellum | 3.1 ± 0.2 | 3.7 ± 0.1 | 4.4 ± 0.45 * |
| Brain stem | 13.0 ± 0.9 | 11.9 ± 0.6 | 13.9 ± 1.2 |
| Olfactory bulb | 72.4 ± 5.1 | 62.4 ± 3.3 | 51.8 ± 1.4 **☨ |
| Dopamine [ng/g] | |||
| Striatum | 9747.0 ± 1580.2 | 9364.5 ± 1458.3 | 8283.8 ± 515.5 ☨ |
| Hippocampus | 67.9 ± 8.7 | 46.3 ± 8.8 | 43.0 ± 9.6 |
| Cortex | 418.0 ± 103.4 | 457.7 ± 116.7 | 294.3 ± 34.7 ☨ |
| Cerebellum | 20.2 ± 1.6 | 16.7 ± 0.7 | 13.7 ± 0.7 ***☨ |
| Brain stem | 123.0 ± 7.3 | 106.6 ± 7.3 | 116.9 ± 8.0 |
| Olfactory bulb | 407.8 ± 20.0 | 331.9 ± 16.7 ** | 304.8 ± 10.1 *** |
| Histamine [ng/g] | |||
| Striatum | 452.8 ± 28.3 | 133.7 ± 28.5 *** | 198.1 ± 12.2 *** |
| Hippocampus | 53.3 ± 6.8 | 48.3 ± 4.8 | 39.6 ± 5.3 |
| Cortex | 64.1 ± 4.9 | 57.9 ± 4.1 | 55.9 ± 4.5 |
| Cerebellum | 60.9 ± 4.0 | 54.7 ± 3.2 | 44.9 ± 4.0 * |
| Brain stem | 65.3 ± 10.4 | 63.5 ± 4.8 | 60.8 ± 5.6 |
| Olfactory bulb | 155.9 ± 16.2 | 112.9± 8.6 * | 115.3 ± 7.6 * |
| Serotonin [ng/g] | |||
| Striatum | 438.0 ± 44.8 | 193.3 ± 33.7 *** | 320.2 ± 14.8 *☨ |
| Hippocampus | 704.0 ± 78.3 | 800.0 ± 51.7 | 651.3 ± 55.8 |
| Cortex | 546.6 ± 40.0 | 565.6 ± 41.9 | 481.3 ± 24.8 |
| Cerebellum | 370.1 ± 36.8 | 404.4 ± 30.5 | 260.8 ± 24.5 |
| Brain stem | 1348.0 ± 56.8 | 1407.9 ± 49.9 | 1288.4 ± 69.4 |
| Olfactory bulb | 666.8 ± 26.8 | 662.2 ± 43.3 | 616.9 ± 38.1 |
| [ng/mL] | Control | Low-Dose | High-Dose |
|---|---|---|---|
| 3-MT | 0.91 ± 0.13 | 0.99 ± 0.078 | 1.05 ± 0.13 |
| Dopamine | N.D. | N.D. | N.D. |
| Histamine | 131 ± 32.78 | 58.90 ± 6.88 * | 45.08 ± 3.43 ** |
| Serotonin | 1235 ± 261 | 1460 ± 580 | 1075 ± 159 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hirai, A.; Yamazaki, R.; Kobayashi, A.; Kimura, T.; Nomiyama, K.; Shimma, S.; Nakayama, S.M.M.; Ishizuka, M.; Ikenaka, Y. Detection of Changes in Monoamine Neurotransmitters by the Neonicotinoid Pesticide Imidacloprid Using Mass Spectrometry. Toxics 2022, 10, 696. https://doi.org/10.3390/toxics10110696
Hirai A, Yamazaki R, Kobayashi A, Kimura T, Nomiyama K, Shimma S, Nakayama SMM, Ishizuka M, Ikenaka Y. Detection of Changes in Monoamine Neurotransmitters by the Neonicotinoid Pesticide Imidacloprid Using Mass Spectrometry. Toxics. 2022; 10(11):696. https://doi.org/10.3390/toxics10110696
Chicago/Turabian StyleHirai, Anri, Ryo Yamazaki, Atsushi Kobayashi, Takashi Kimura, Kei Nomiyama, Shuichi Shimma, Shouta M. M. Nakayama, Mayumi Ishizuka, and Yoshinori Ikenaka. 2022. "Detection of Changes in Monoamine Neurotransmitters by the Neonicotinoid Pesticide Imidacloprid Using Mass Spectrometry" Toxics 10, no. 11: 696. https://doi.org/10.3390/toxics10110696
APA StyleHirai, A., Yamazaki, R., Kobayashi, A., Kimura, T., Nomiyama, K., Shimma, S., Nakayama, S. M. M., Ishizuka, M., & Ikenaka, Y. (2022). Detection of Changes in Monoamine Neurotransmitters by the Neonicotinoid Pesticide Imidacloprid Using Mass Spectrometry. Toxics, 10(11), 696. https://doi.org/10.3390/toxics10110696