Composition, Protein Profile and Rheological Properties of Pseudocereal-Based Protein-Rich Ingredients

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cereal and Pseudocereal Flour Ingredients
2.2. Chemical Composition
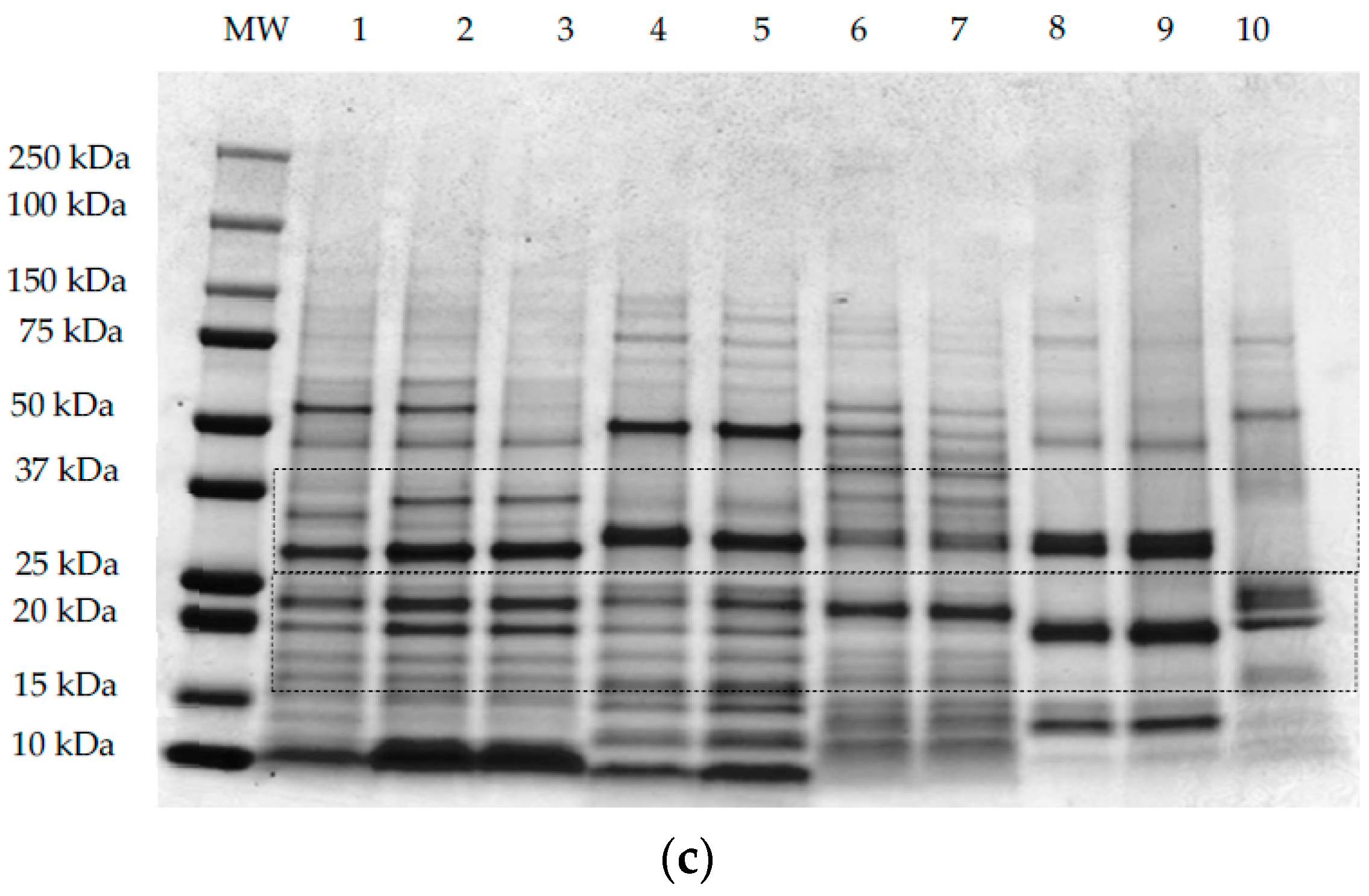
2.3. Electrophoretic Protein Profile Analysis
2.4. Microstructural Analysis
2.5. Pasting Behaviour
2.6. Statistical Analysis
3. Results and Discussion
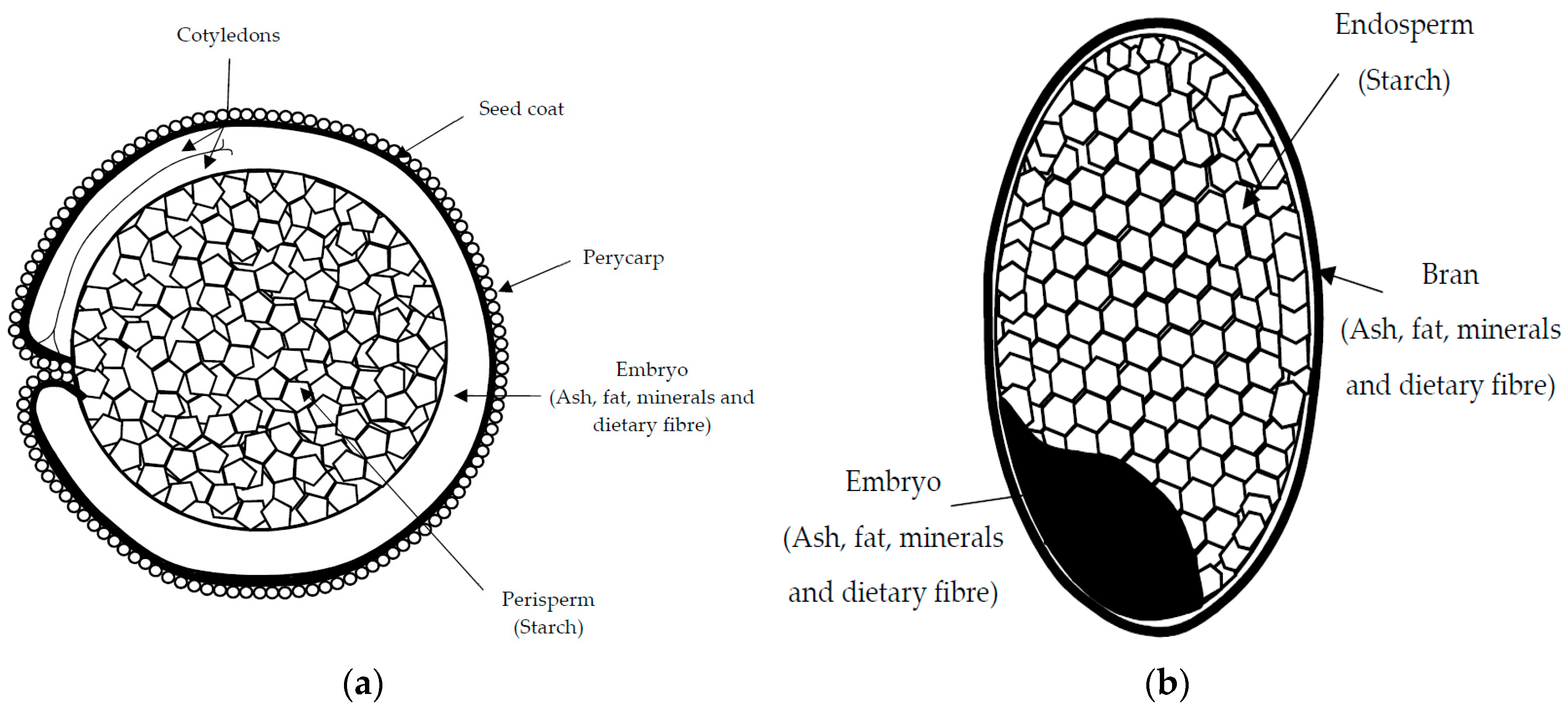
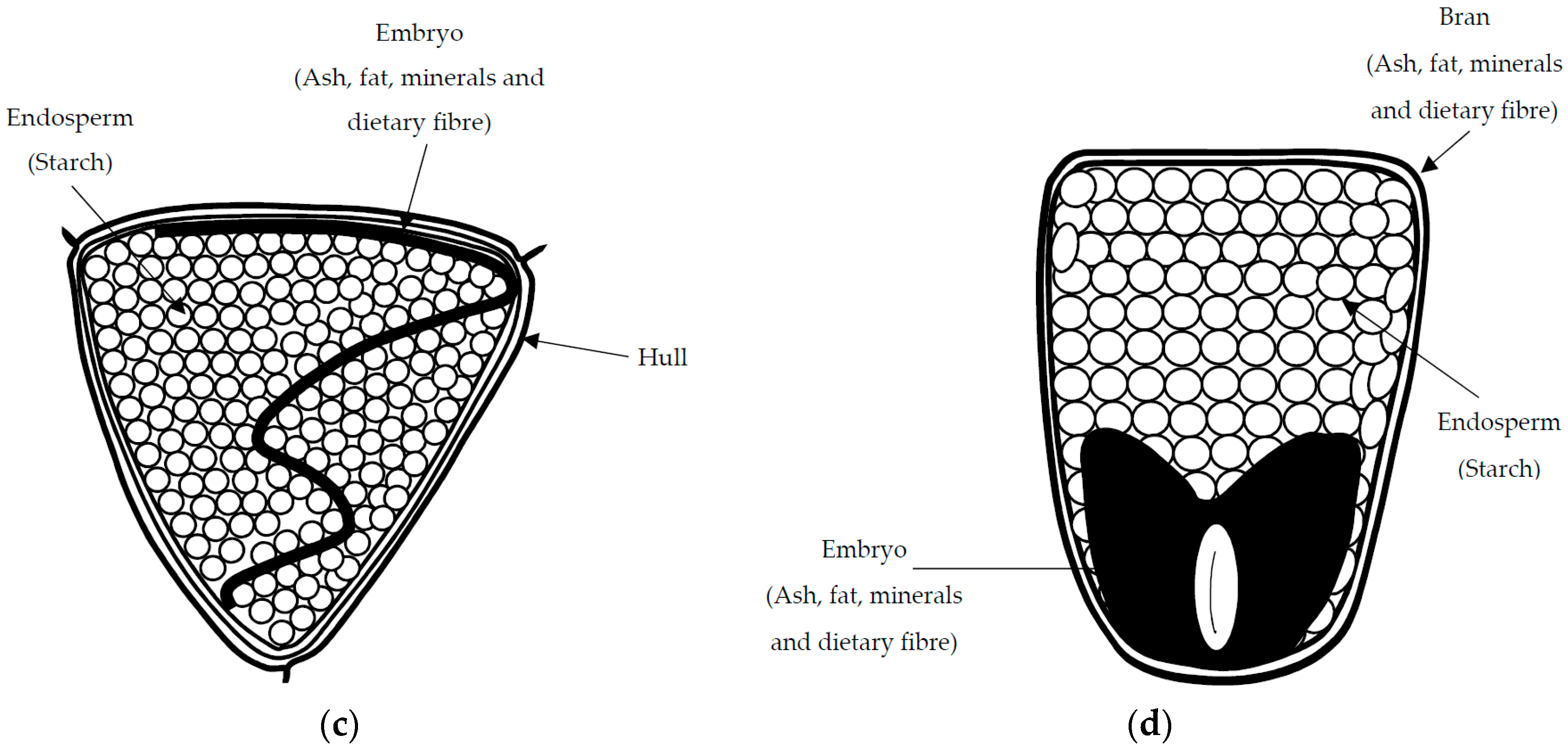
3.1. Chemical Composition
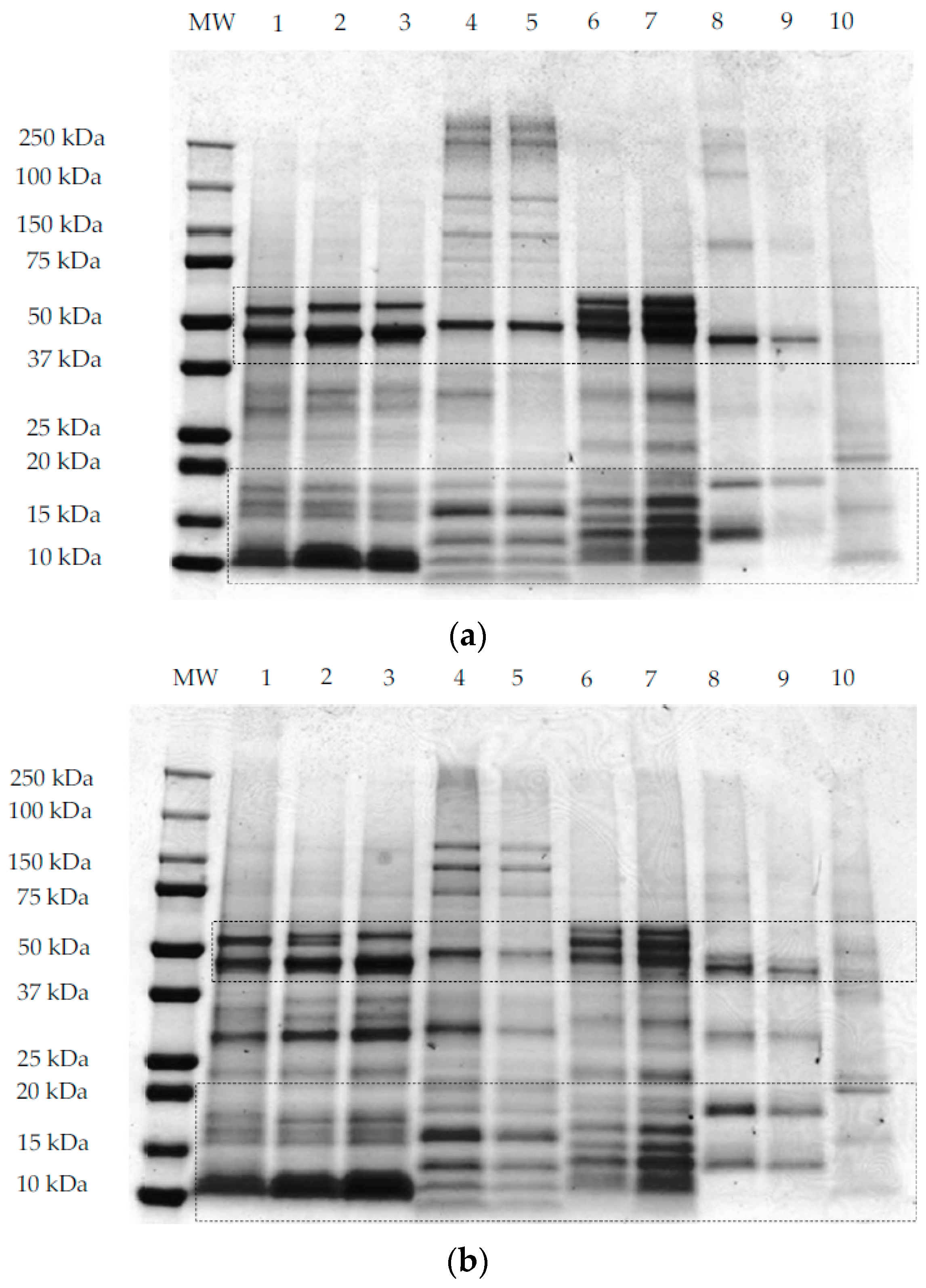
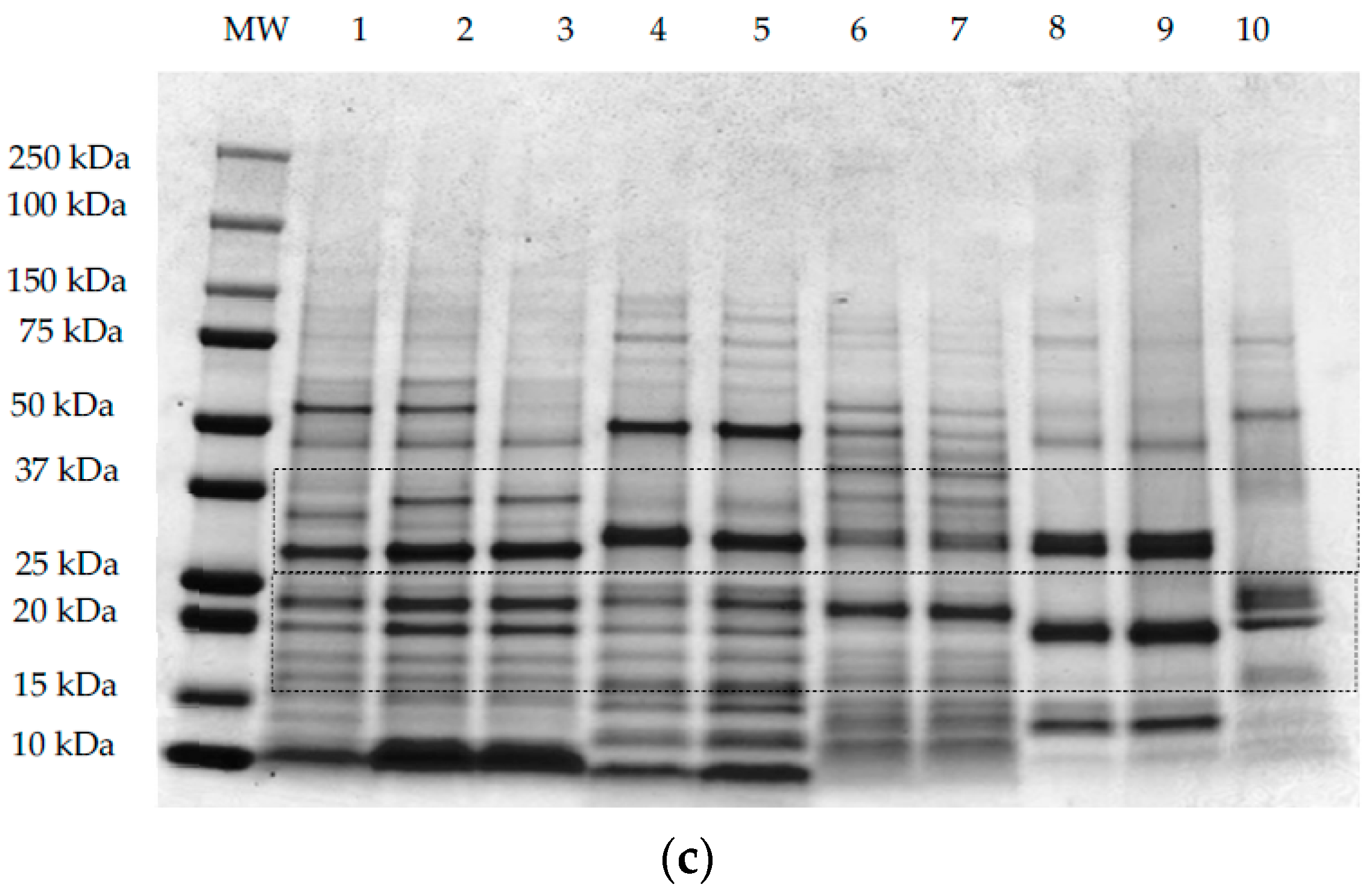
3.2. Protein Profile by SDS-PAGE Electrophoresis
3.3. Starch Granules: Shape and Size
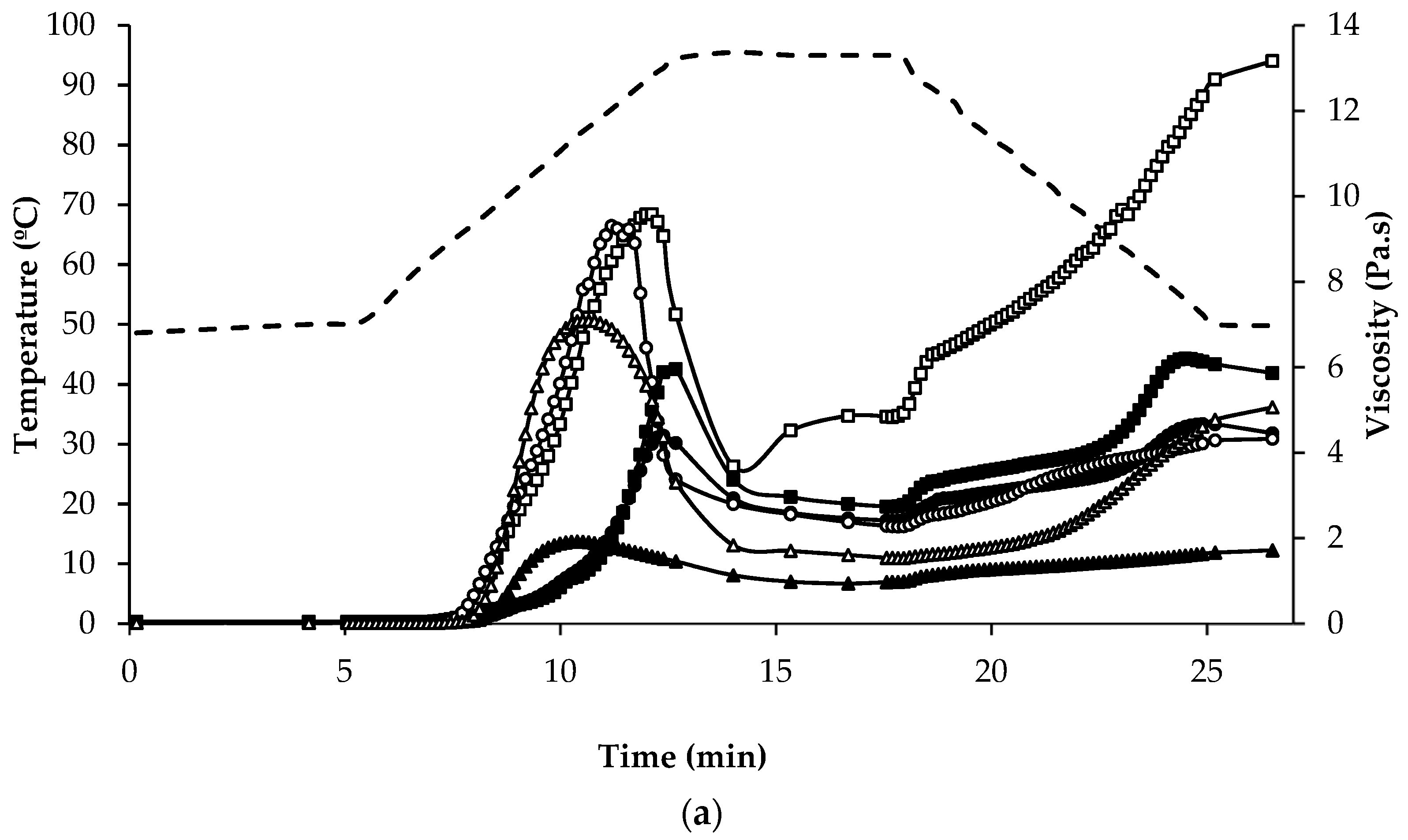
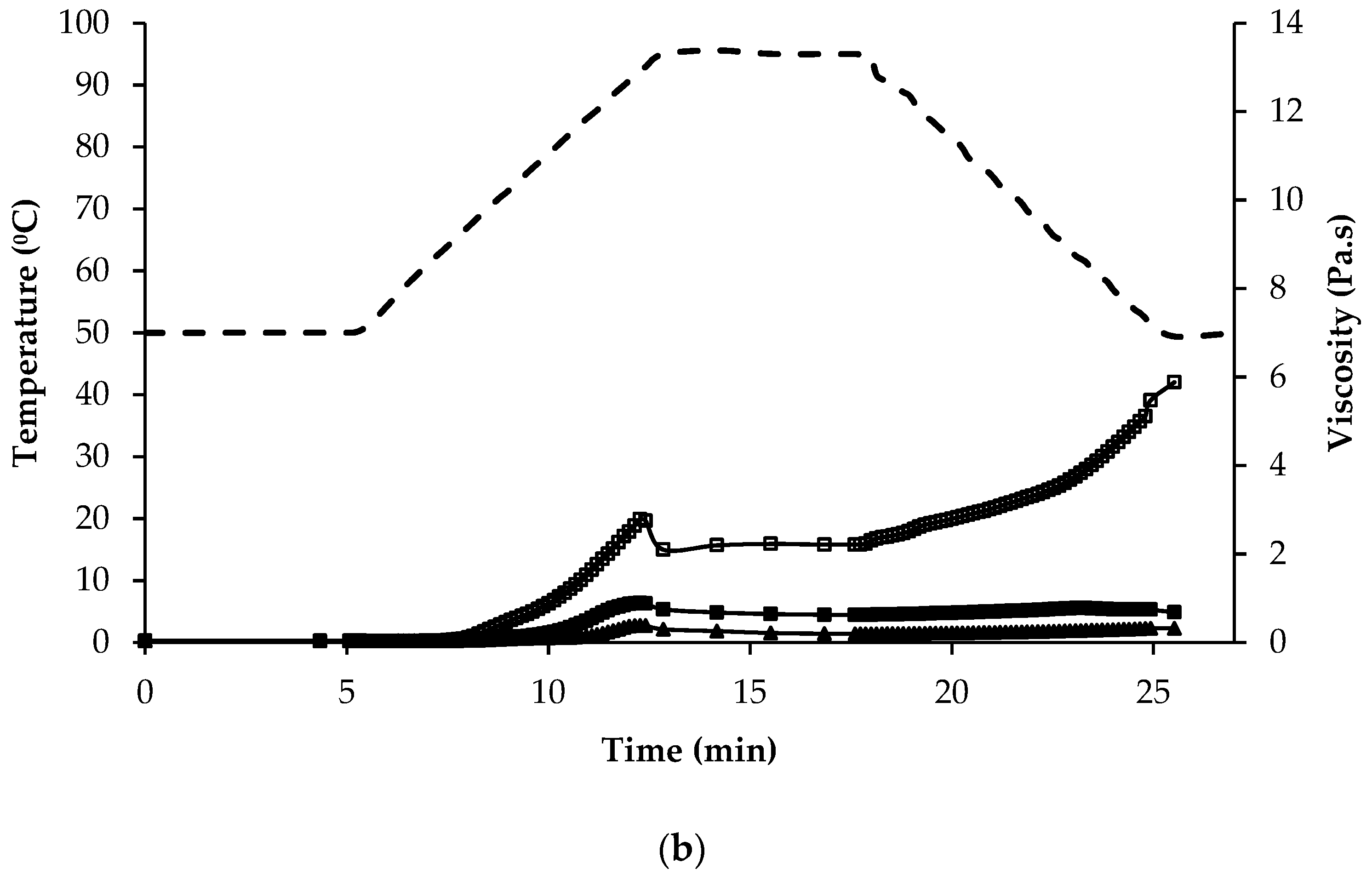
3.4. Pasting Properties
4. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Henchion, M.; Hayes, M.; Mullen, A.M.; Fenelon, M.; Tiwari, B. Future protein supply and demand: strategies and factors influencing a sustainable equilibrium. Foods 2017, 6, 53. [Google Scholar] [CrossRef] [PubMed]
- Day, L. Proteins from land plants—Potential resources for human nutrition and food security. Trends Food Sci. Technol. 2013, 32, 25–42. [Google Scholar] [CrossRef]
- Nations, U. Revision of World Population Prospects; United Nations: New York, NY, USA, 2015. [Google Scholar]
- Aiking, H. Future protein supply. Trends Food Sci. Technol. 2011, 22, 112–120. [Google Scholar] [CrossRef]
- Schutyser, M.A.I.; Pelgrom, P.J.M.; van der Goot, A.J.; Boom, R.M. Dry fractionation for sustainable production of functional legume protein concentrates. Trends Food Sci. Technol. 2015, 45, 327–335. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the functional properties of enzymatic plant protein hydrolysates in food systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef]
- Alting, A.C.; Van De Velde, F. Proteins as clean label ingredients in foods and beverages. In Natural Food Additives, Ingredients and Flavourings; Baines, D., Seal, R., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2012; pp. 197–211. ISBN 978-1-84-569811-9. [Google Scholar]
- Jacobsen, S.E.; Sørensen, M.; Pedersen, S.M.; Weiner, J. Feeding the world: Genetically modified crops versus agricultural biodiversity. Agron. Sustain. Dev. 2013, 33, 651–662. [Google Scholar] [CrossRef]
- Haros, C.M.; Schoenlechner, R. Pseudocereals: Chemistry and Technology, 1st ed.; Wiley Blackwell: Chichester, UK, 2017; ISBN 978-1-11-893825-6. [Google Scholar]
- Schoenlechner, R.; Siebenhandl, S.; Berghofer, E. Pseudocereals. In Gluten-Free Cereal Product and Beverages; Arendt, E.K., Dal Bello, F., Eds.; Elsevier: New York, NY, USA, 2008; pp. 149–190. ISBN 978-0-12-373739-7. [Google Scholar]
- Taylor, J.; Awika, J. Gluten-Free Ancient Grains. Cereals, Pseudocereals, and Legumes: Sustainable, Nutritious, and Health-Promoting Foods for the 21st Century, 1st ed.; Taylor, J., Awika, J., Eds.; Woodhead Publishing Limited: Duxford, UK, 2017; ISBN 978-0-08-100866-9. [Google Scholar]
- Mota, C.; Santos, M.; Mauro, R.; Samman, N.; Matos, A.S.; Torres, D.; Castanheira, I. Protein content and amino acids profile of pseudocereals. Food Chem. 2016, 193, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Tester, R.F.; Karkalas, J.; Qi, X. Starch—Composition, fine structure and architecture. J. Cereal Sci. 2004, 39, 151–165. [Google Scholar] [CrossRef]
- Steadman, K.J.J.; Burgoon, M.S.S.; Lewis, B.A.A.; Edwardson, S.E.E.; Obendorf, R.L.L. Buckwheat seed milling fractions: Description, macronutrient composition and dietary fibre. J. Cereal Sci. 2001, 33, 271–278. [Google Scholar] [CrossRef]
- Schirmer, M.; Höchstötter, A.; Jekle, M.; Arendt, E.; Becker, T. Physicochemical and morphological characterization of different starches with variable amylose/amylopectin ratio. Food Hydrocoll. 2013, 32, 52–63. [Google Scholar] [CrossRef]
- Horstmann, S.; Belz, M.M.; Heitmann, M.; Zannini, E.; Arendt, E. Fundamental study on the impact of gluten-free starches on the quality of gluten-free model breads. Foods 2016, 5, 30. [Google Scholar] [CrossRef] [PubMed]
- Janssen, F.; Pauly, A.; Rombouts, I.; Jansens, K.J.A.; Deleu, L.J.; Delcour, J.A. Proteins of amaranth (Amaranthus spp.), buckwheat (Fagopyrum spp.), and quinoa (Chenopodium spp.): A food science and technology perspective. Compr. Rev. Food Sci. Food Saf. 2017, 16, 39–58. [Google Scholar] [CrossRef]
- Mäkinen, O.E.; Zannini, E.; Koehler, P.; Arendt, E.K. Heat-denaturation and aggregation of quinoa (Chenopodium quinoa) globulins as affected by the pH value. Food Chem. 2016, 196, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Venskutonis, P.R.; Kraujalis, P. Nutritional components of amaranth seeds and vegetables: A review on composition, properties, and uses. Compr. Rev. Food Sci. Food Saf. 2013, 12, 381–412. [Google Scholar] [CrossRef]
- Amagliani, L.; O’Regan, J.; Kelly, A.L.; O’Mahony, J.A. Composition and protein profile analysis of rice protein ingredients. J. Food Compos. Anal. 2016, 59, 18–26. [Google Scholar] [CrossRef]
- DeVries, J.W. Dietary Fibre: New Frontiers for Food and Health; van der Kamp, J.W., Jones, J.M., McCleary, B.V., Topping, D.L., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2010; ISBN 978-9-08-686128-6. [Google Scholar]
- Codex Alimentarius Commission. Report of the 27th Session of the Codex Committee on Nutrition and Foods for Special Dietary Uses; Codex Alimentarius Commission: Rome, Italy, 2005. [Google Scholar]
- Foschia, M.; Peressini, D.; Sensidoni, A.; Brennan, C.S. The effects of dietary fibre addition on the quality of common cereal products. J. Cereal Sci. 2013, 58, 216–227. [Google Scholar] [CrossRef]
- Alvarez-Jubete, L.; Arendt, E.K.; Gallagher, E. Nutritive value and chemical composition of pseudocereals as gluten-free ingredients. Int. J. Food Sci. Nutr. 2009, 60, 240–257. [Google Scholar] [CrossRef] [PubMed]
- Burrieza, H.P.; Lopez-Fernandez, M.P.; Maldonado, S. Analogous reserve distribution and tissue characteristics in quinoa and grass seeds suggest convergent evolution. Front. Plant Sci. 2014, 5, 546. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Jubete, L.; Arendt, E.K.; Gallagher, E. Nutritive value of pseudocereals and their increasing use as functional gluten-free ingredients. Trends Food Sci. Technol. 2010, 21, 106–113. [Google Scholar] [CrossRef]
- Nosworthy, M.G.; Tulbek, M.C.; House, J.D. Does the concentration, isolation, or deflavoring of pea, lentil, and faba bean protein alter protein quality? Cereal Foods World 2017, 62, 139–142. [Google Scholar] [CrossRef]
- Avila Ruiz, G.; Arts, A.; Minor, M.; Schutyser, M. A hybrid dry and aqueous fractionation method to obtain protein-rich fractions from quinoa (Chenopodium quinoa Willd). Food Bioprocess Technol. 2016, 9, 1502–1510. [Google Scholar] [CrossRef]
- Berghout, J.A.M.; Pelgrom, P.J.M.; Schutyser, M.A.I.; Boom, R.M.; van der Goot, A.J. Sustainability assessment of oilseed fractionation processes: A case study on lupin seeds. J. Food Eng. 2015, 150, 117–124. [Google Scholar] [CrossRef]
- Boukid, F.; Folloni, S.; Sforza, S.; Vittadini, E.; Prandi, B. Current trends in ancient grains-based foodstuffs: insights into nutritional aspects and technological applications. Compr. Rev. Food Sci. Food Saf. 2018, 17, 123–136. [Google Scholar] [CrossRef]
- Pelgrom, P.J.M.; Boom, R.M.; Schutyser, M.A.I. Functional analysis of mildly refined fractions from yellow pea. Food Hydrocoll. 2015, 44, 12–22. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis of the Association of Official Analytical Chemists, 18th ed.; AOAC: Washington, DC, USA, 2010. [Google Scholar]
- Nascimento, A.C.; Mota, C.; Coelho, I.; Gueifão, S.; Santos, M.; Matos, A.S.; Gimenez, A.; Lobo, M.; Samman, N.; Castanheira, I. Characterisation of nutrient profile of quinoa (Chenopodium quinoa), amaranth (Amaranthus caudatus), and purple corn (Zea mays L.) consumed in the North of Argentina: Proximates, minerals and trace elements. Food Chem. 2014, 148, 420–426. [Google Scholar] [CrossRef] [PubMed]
- Abugoch James, L.E. Quinoa (Chenopodium quinoa Willd.): Composition, chemistry, nutritional, and functional properties. Adv. Food Nutr. Res. 2009, 58, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Wang, S.; Zhu, F. Physicochemical properties of quinoa starch. Carbohydr. Polym. 2016, 137, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Bradley, R.L. Moisture and Total Solids Analysis. In Food Analysis; Nielsen, S., Ed.; Springer: West Lafayette, IN, USA, 2010; pp. 85–104. ISBN 1441914781. [Google Scholar]
- Roudaut, G.; Debeaufort, F. Moisture loss, gain and migration in foods. In Food and Beverage Stability and Shelf Life; Elsevier: New York, NY, USA, 2011; pp. 63–105. ISBN 978-1-84-569701-3. [Google Scholar]
- Schuck, P.; Dolivet, A.; Jeantet, R. Determination of Dry Matter and Total Dry Matter. In Analytical Methods for Food and Dairy Powders; Schuck, P., Dolivet, A., Jeantet, R., Eds.; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2012; pp. 45–57. ISBN 978-1-11-830739-7. [Google Scholar]
- Tyler, R.T.; Youngs, C.G.; Sosulski, F.W. Air Classification of legumes. I. Separation efficiency, yield, and composition of the starch and protein fractions. Cereal Chem. 1981, 58, 144–148. [Google Scholar]
- Arendt, E.K.; Zannini, E. Cereal Grains for the Food and Beverage Industries, 1st ed.; Woodhead Publishing Limited: Cambridge, UK, 2013; ISBN 978-0-85-709413-1. [Google Scholar]
- Gamel, T.H.; Mesallam, A.S.; Damir, A.A.; Shekib, L.A.; Linssen, J.P. Characterization of amaranth seeds oil. J. Food Lipids 2007, 14, 323–334. [Google Scholar] [CrossRef]
- Navruz-Varli, S.; Sanlier, N. Nutritional and health benefits of quinoa (Chenopodium quinoa Willd.). J. Cereal Sci. 2016, 69, 371–376. [Google Scholar] [CrossRef]
- Barrera, G.N.; Bustos, M.C.; Iturriaga, L.; Flores, S.K.; Leon, A.E.; Ribotta, P.D. Effect of damaged starch on the rheological properties of wheat starch suspensions. J. Food Eng. 2013, 116, 233–239. [Google Scholar] [CrossRef]
- Nowak, V.; Du, J.; Charrondière, U.R. Assessment of the nutritional composition of quinoa (Chenopodium quinoa Willd.). Food Chem. 2016, 193, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Ando, H.; Chen, Y.-C.; Tang, H.; Mayumi, S.; Watanabe, K.; Mitsunaga, T. Food components in fractions of quinoa seed. Food Sci. Technol. Res. 2002, 8, 80–84. [Google Scholar] [CrossRef]
- Repo-Carrasco, R.; Peña, J.; Kallio, H.; Salminen, S. Dietary fiber and other functional components in two varieties of crude and extruded kiwicha (Amaranthus caudatus). J. Cereal Sci. 2009, 49, 219–224. [Google Scholar] [CrossRef]
- Ruales, J.; Grijalva, Y.; Lopez-Jaramillo, P.; Nair, B. The nutritional quality of an infant food from quinoa and its effect on the plasma level of insulin-like growth factor-1 (IGF-1) in undernourished children. Int. J. Food Sci. Nutr. 2002, 53, 143–154. [Google Scholar] [CrossRef] [PubMed]
- Repo-Carrasco, R.; Astuhuaman Serna, L. Quinoa (Chenopodium quinoa, Willd.) as a source of dietary fiber and other functional components. Food Sci. Technol. 2009, 31, 225–230. [Google Scholar] [CrossRef]
- Mir, N.A.; Riar, C.S.; Singh, S. Nutritional constituents of pseudo cereals and their potential use in food systems: A review. Trends Food Sci. Technol. 2018, 75, 170–180. [Google Scholar] [CrossRef]
- Vasco-Méndez, N.L.; Paredes-López, O. Antigenic homology between amaranth glutelins and other storage proteins. Food Biochem. 1994, 18, 227–238. [Google Scholar] [CrossRef]
- Milisavljević, M.D.; Timotijević, G.S.; Radović, S.R.; Brkljačić, J.M.; Konstantinović, M.M.; Maksimović, V.R. Vicilin-like storage globulin from buckwheat (Fagopyrum esculentum Moench) seeds. J. Agric. Food Chem. 2004, 52, 5258–5262. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.M.; Ma, C.Y. Conformational study of globulin from common buckwheat (Fagopyrum esculentum Moench) by fourier transform infrared spectroscopy and differential scanning calorimetry. J. Agric. Food Chem. 2005, 53, 8046–8053. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.J.; Lamsa, B.P. Zein extraction from corn, corn products, and coproducts and modifications for various applications: A review. Cereal Chem. 2011, 88, 159–173. [Google Scholar] [CrossRef]
- Valencia-Chamorro, S.A. Quinoa. In Encyclopedia of Food Sciences and Nutrition; Elsevier: Amsterdam, The Netherlands, 2003; pp. 4895–4902. [Google Scholar]
- Gorinstein, S.; Pawelzik, E.; Delgado-Licon, E.; Haruenkit, R.; Weisz, M.; Trakhtenberg, S. Characterisation of pseudocereal and cereal proteins by protein and amino acid analyses. J. Sci. Food Agric. 2002, 82, 886–891. [Google Scholar] [CrossRef]
- Radovic, R.S.; Maksimovic, R.V.; Brkljacic, M.J.; Varkonji Gasic, I.E.; Savic, P.A. 2S albumin from buckwheat (Fagopyrum esculentum Moench) seeds. J. Agric. Food Chem. 1999, 47, 1467–1470. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Zhu, F. Molecular structure of quinoa starch. Carbohydr. Polym. 2017, 158, 124–132. [Google Scholar] [CrossRef] [PubMed]
- Christa, K.; Soral-Smietana, M.; Lewandowicz, G. Buckwheat starch: Structure, functionality and enzyme in vitro susceptibility upon the roasting process. Int. J. Food Sci. Nutr. 2009, 60, 140–154. [Google Scholar] [CrossRef] [PubMed]
- Debet, M.R.; Gidley, M.J. Three classes of starch granule swelling: Influence of surface proteins and lipids. Carbohydr. Polym. 2006, 64, 452–465. [Google Scholar] [CrossRef]
- Nienke, L.; Chang, P.R.; Tyler, R.T. Analytical, biochemical and physicochemical aspects of starch granule size, with emphasis on small granule starches: A review. Starch Stärke 2004, 56, 89–99. [Google Scholar] [CrossRef]
- Rayner, M.; Timgren, A.; Sjoo, M.; Dejmek, P. Quinoa starch granules: A candidate for stabilising food-grade pickering emulsions. J. Sci. Food Agric. 2012, 92, 1841–1847. [Google Scholar] [CrossRef] [PubMed]
- Ragaee, S.; Abdel-Aal, E.S.M. Pasting properties of starch and protein in selected cereals and quality of their food products. Food Chem. 2006, 95, 9–18. [Google Scholar] [CrossRef]
- Bulut-Solak, B.; Alonso-Miravalles, L.; O’Mahony, J.A. Composition, morphology and pasting properties of Orchis anatolica tuber gum. Food Hydrocoll. 2016, 69, 483–490. [Google Scholar] [CrossRef]
- Wang, S.; Li, C.; Copeland, L.; Niu, Q.; Shuo, W. Starch retrogradation: A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2015, 14, 568–585. [Google Scholar] [CrossRef]
- Yoshimoto, Y.; Egashira, T.; Hanashiro, I.; Ohinata, H.; Takase, Y.; Takeda, Y. Molecular structure and some physicochemical properties of buckwheat starches. Cereal Chem. 2004, 81, 515–520. [Google Scholar] [CrossRef]
- Izydorczyk, M.S.; McMillan, T.; Bazin, S.; Kletke, J.; Dushnicky, L.; Dexter, J. Canadian buckwheat: A unique, useful and under-utilized crop. Can. J. Plant Sci. 2013, 94, 509–524. [Google Scholar] [CrossRef]
- Lu, L.; Murphy, K.; Baik, B.K. Genotypic variation in nutritional composition of buckwheat groats and husks. Cereal Chem. 2013, 90, 132–137. [Google Scholar] [CrossRef]
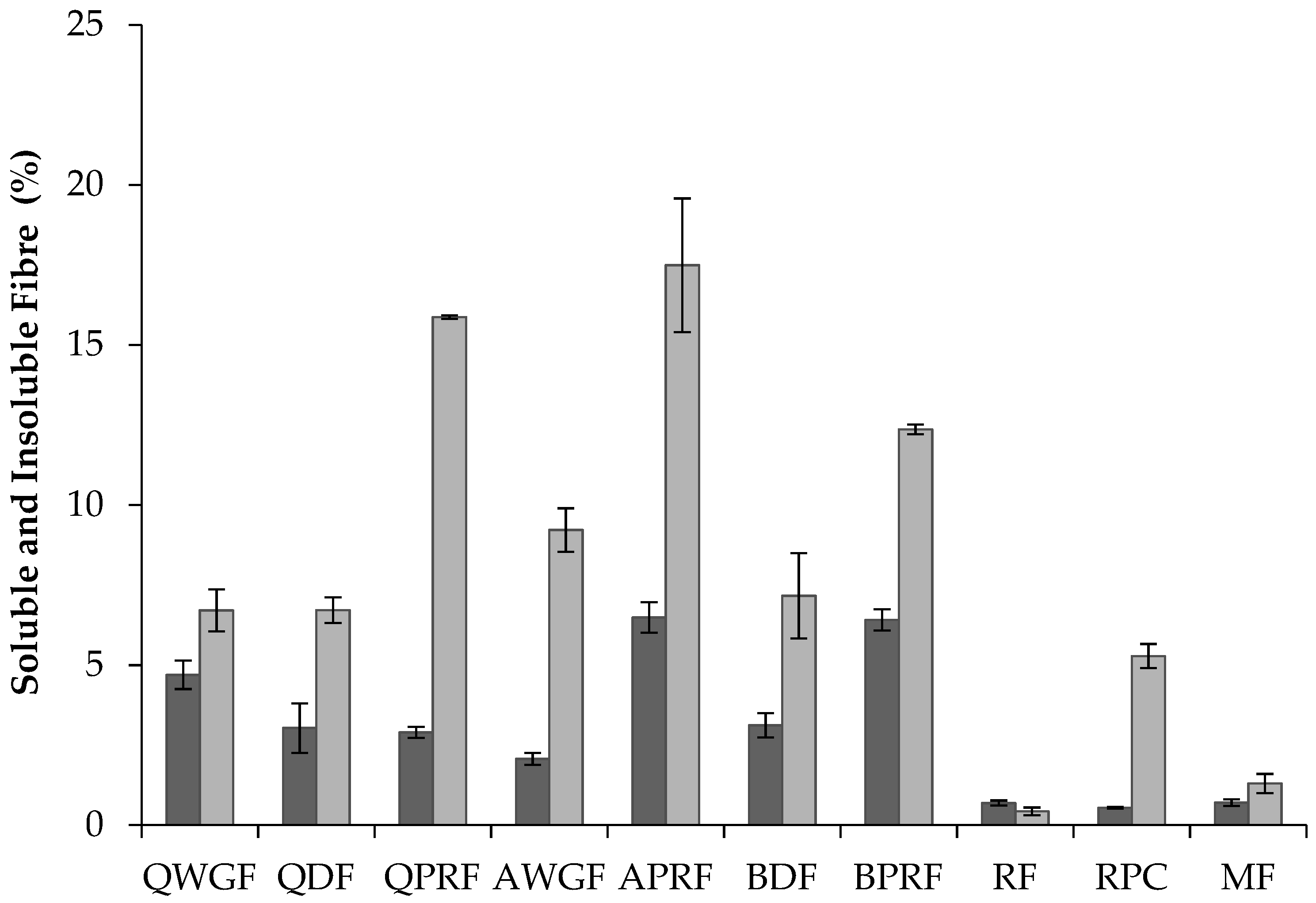
 ) and insoluble (
) and insoluble (  ) dietary fibre content (% w/w) of quinoa wholegrain flour (QWGF), quinoa dehulled flour (QDF), quinoa protein-rich flour (QPRF), amaranth wholegrain flour (AWGF), amaranth protein-rich flour (APRF), buckwheat dehulled flour (BDF), buckwheat protein-rich flour (BPRF), rice flour (RF), rice protein concentrate (RPC), and maize flour (MF).
) and insoluble ( ) dietary fibre content (% w/w) of quinoa wholegrain flour (QWGF), quinoa dehulled flour (QDF), quinoa protein-rich flour (QPRF), amaranth wholegrain flour (AWGF), amaranth protein-rich flour (APRF), buckwheat dehulled flour (BDF), buckwheat protein-rich flour (BPRF), rice flour (RF), rice protein concentrate (RPC), and maize flour (MF).
) dietary fibre content (% w/w) of quinoa wholegrain flour (QWGF), quinoa dehulled flour (QDF), quinoa protein-rich flour (QPRF), amaranth wholegrain flour (AWGF), amaranth protein-rich flour (APRF), buckwheat dehulled flour (BDF), buckwheat protein-rich flour (BPRF), rice flour (RF), rice protein concentrate (RPC), and maize flour (MF).
) and insoluble ( ) dietary fibre content (% w/w) of quinoa wholegrain flour (QWGF), quinoa dehulled flour (QDF), quinoa protein-rich flour (QPRF), amaranth wholegrain flour (AWGF), amaranth protein-rich flour (APRF), buckwheat dehulled flour (BDF), buckwheat protein-rich flour (BPRF), rice flour (RF), rice protein concentrate (RPC), and maize flour (MF).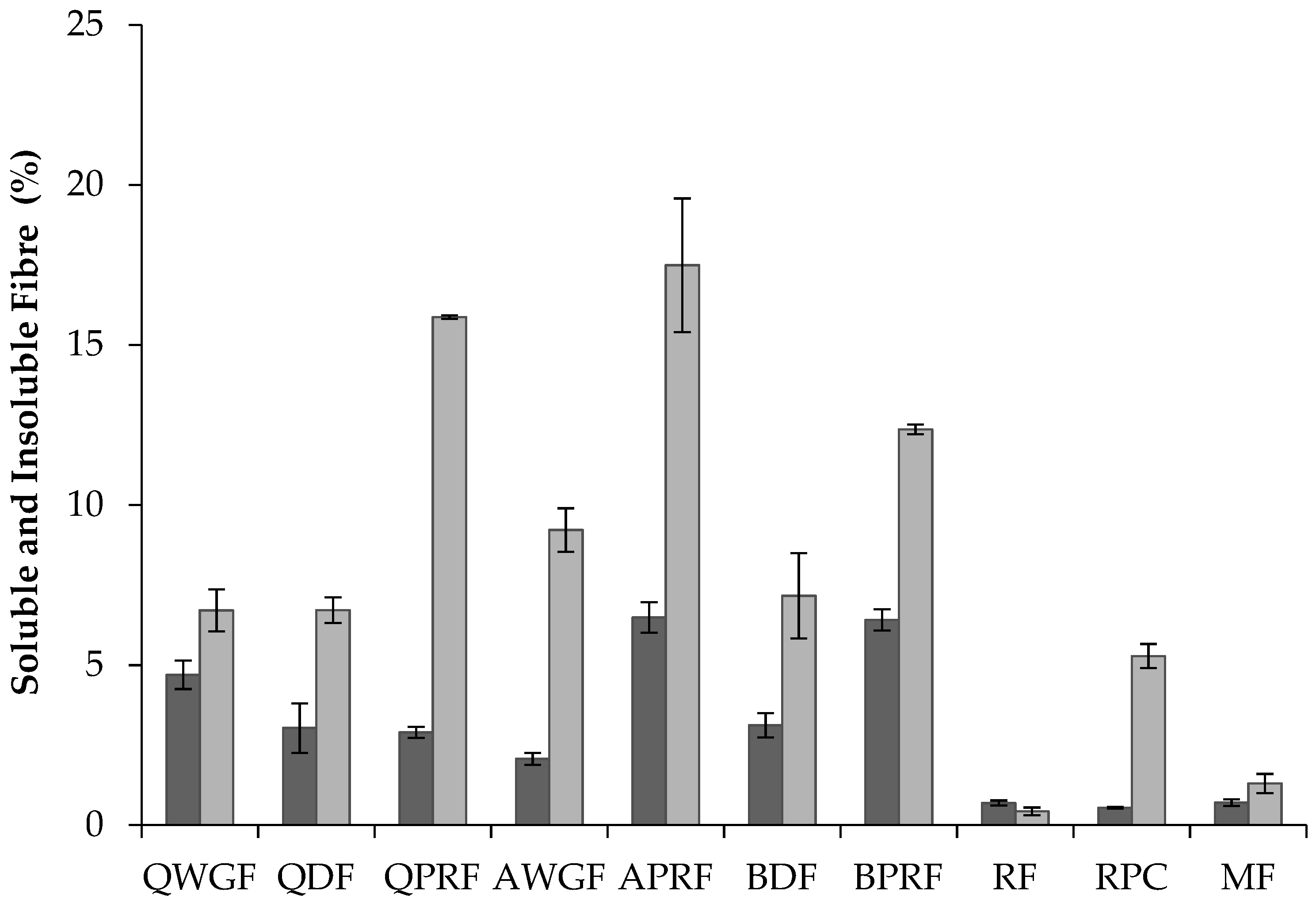
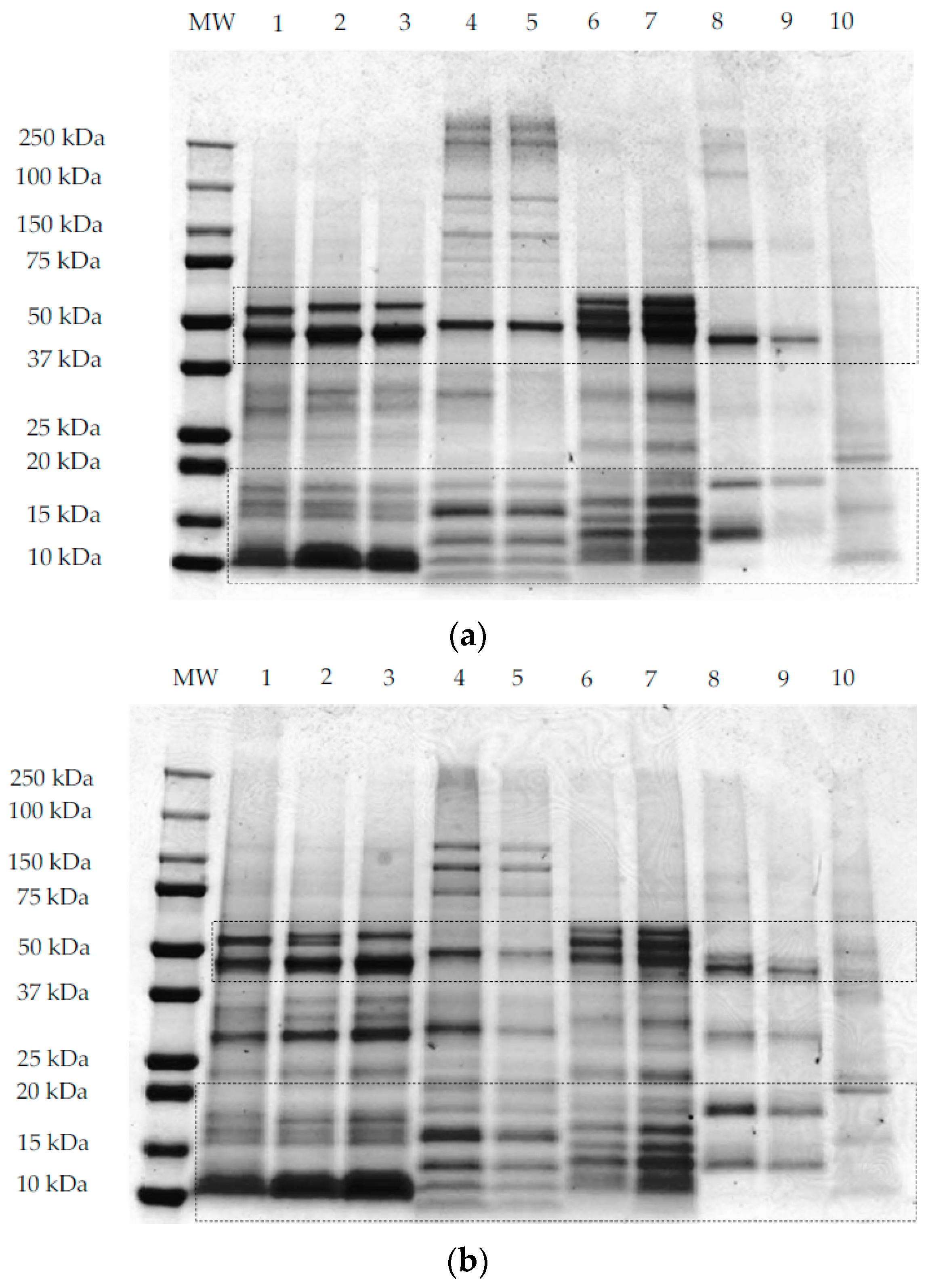

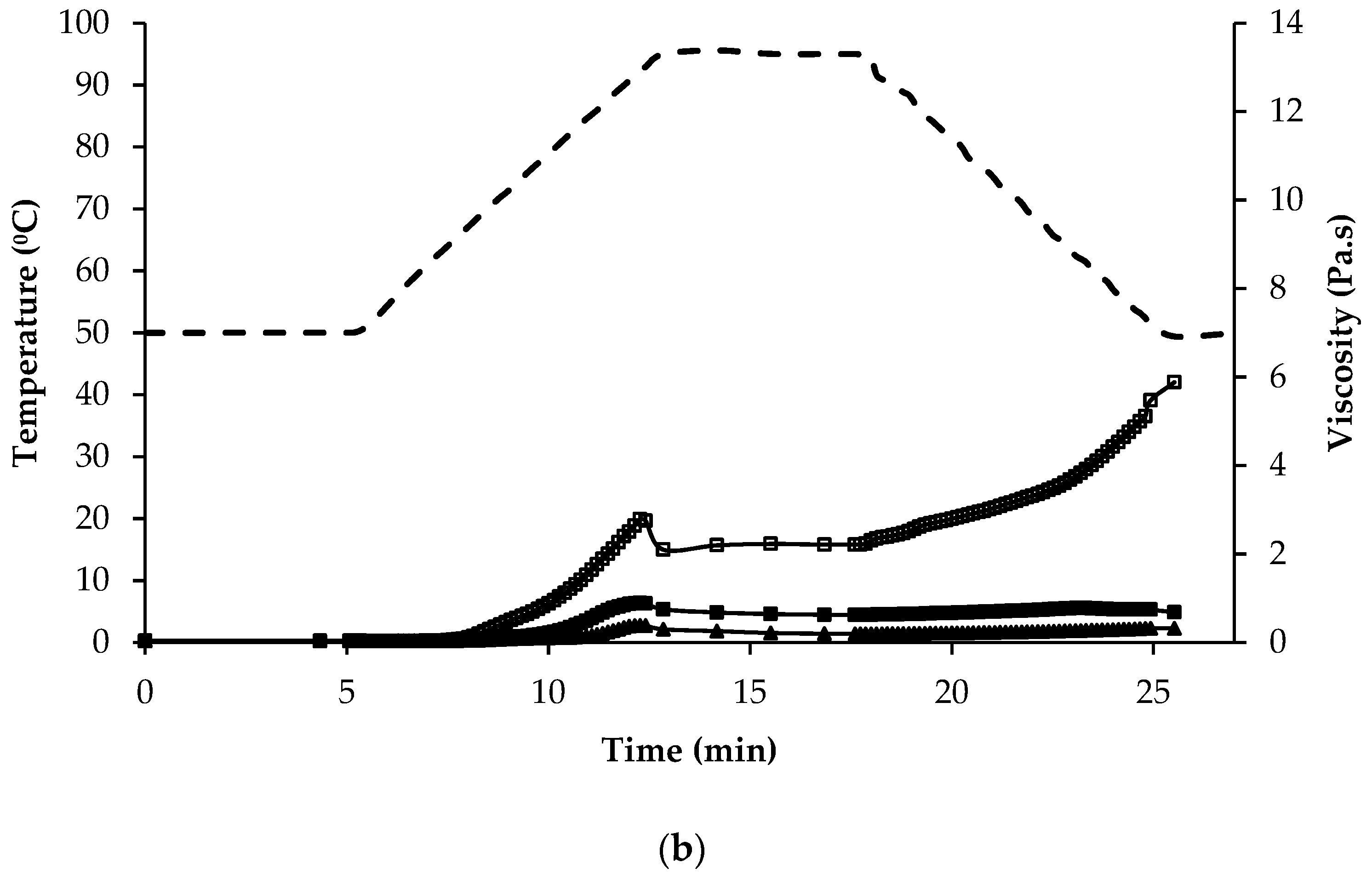
 ), quinoa dehulled flour (
), quinoa dehulled flour (  ), amaranth wholegrain flour (
), amaranth wholegrain flour (  ), buckwheat dehulled flour (BDF) (
), buckwheat dehulled flour (BDF) (  ), rice flour (RF) (
), rice flour (RF) (  ), and maize flour (
), and maize flour (  ); (b) of protein enriched flour ingredients: quinoa protein-rich flour ( ), amaranth protein-rich flour ( ), buckwheat protein-rich flour ( ).
), quinoa dehulled flour ( ), amaranth wholegrain flour ( ), buckwheat dehulled flour (BDF) ( ), rice flour (RF) ( ), and maize flour ( ); (b) of protein enriched flour ingredients: quinoa protein-rich flour ( ), amaranth protein-rich flour ( ), buckwheat protein-rich flour ( ).
); (b) of protein enriched flour ingredients: quinoa protein-rich flour ( ), amaranth protein-rich flour ( ), buckwheat protein-rich flour ( ).
), quinoa dehulled flour ( ), amaranth wholegrain flour ( ), buckwheat dehulled flour (BDF) ( ), rice flour (RF) ( ), and maize flour ( ); (b) of protein enriched flour ingredients: quinoa protein-rich flour ( ), amaranth protein-rich flour ( ), buckwheat protein-rich flour ( ).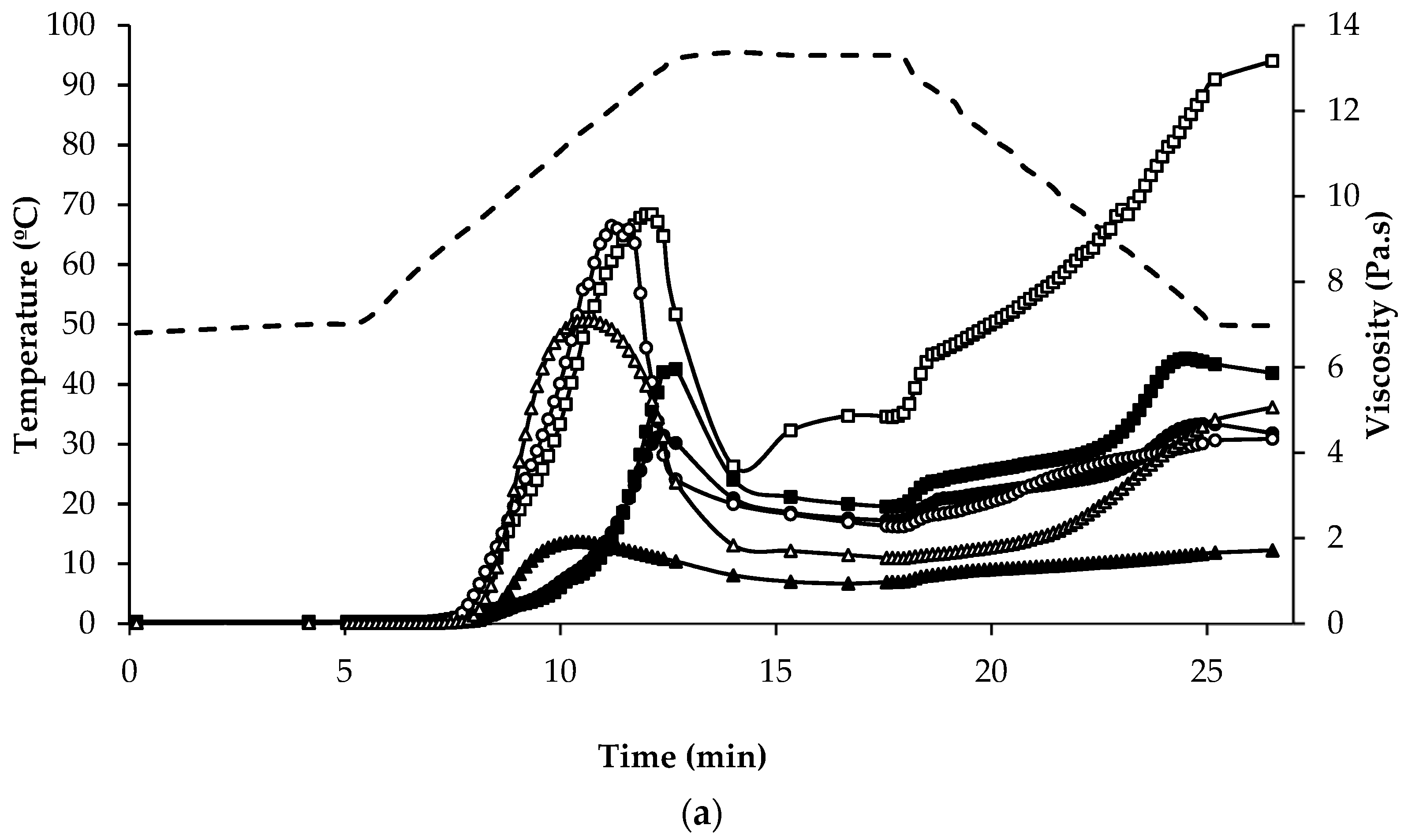

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moisture | Ash | Protein (% w/w) | Fat | Carbohydrate | Starch | Damaged Starch (% Total Starch) | TDF (% w/w) | |
|---|---|---|---|---|---|---|---|---|
| Quinoa | ||||||||
| QWGF | 9.01 ± 0.10 d | 2.30 ± 0.00 c | 13.1 ± 0.10 b | 6.54 ± 0.07 d | 69.0 ± 0.27 d | 60.0 ± 2.58 d | 10.6 ± 0.47 d | 11.4 ± 1.10 b |
| QDF | 8.86 ± 0.25 d | 1.80 ± 0.10 b | 15.7 ± 0.30 b | 5.36 ± 0.61 d | 68.3 ± 1.26 d | 50.5 ± 1.40 bc | 11.7 ± 0.32 e | 9.75 ± 1.17 b |
| QPRF | 5.25 ± 0.25 a | 3.60 ± 0.19 e | 33.3 ± 1.10 d | 12.8 ± 0.73 e | 45.0 ± 2.27 b | 21.4 ± 0.81 a | 10.4 ± 0.40 d | 18.8 ± 0.23 c |
| Amaranth | ||||||||
| AWGF | 8.94 ± 0.05 d | 2.40 ± 0.02 c | 14.6 ± 0.30 b | 6.04 ± 0.10 d | 68.1 ± 0.47 d | 52.8 ± 1.45 c | 12.2 ± 0.35 e | 11.3 ± 0.86 b |
| APRF | 7.76 ± 0.12 b | 6.86 ± 0.18 f | 38.6 ± 1.74 e | 16.6 ± 0.08 f | 30.2 ± 2.12 a | 20.3 ± 0.31 a | 2.61 ± 0.01 b | 24.0 ± 2.56 d |
| Buckwheat | ||||||||
| BDF | 8.75 ± 0.11 d | 1.51 ± 0.31 b | 14.2 ± 0.06 b | 2.77 ± 0.05 bc | 72.8 ± 0.53 e | 61.6 ± 0.12 d | 1.52 ± 0.06 a | 10.3 ± 1.72 b |
| BPRF | 6.86 ± 0.17 c | 3.05 ± 0.10 d | 20.5 ± 0.90 c | 4.76 ± 0.15 cd | 64.8 ± 1.32 c | 47.3 ± 1.20 b | 2.22 ± 0.07 ab | 19.0 ± 0.48 c |
| Rice | ||||||||
| RF | 8.89 ± 0.19 d | 0.85 ± 0.05 a | 8.22 ± 0.14 a | 0.71 ± 0.08 a | 81.3 ± 0.46 f | 78.5 ± 0.82 e | 10.7 ± 0.14 f | 1.12 ± 0.20 a |
| RPC | 6.24 ± 0.08 a | 3.42 ± 0.24 d | 75.0 ± 0.38 f | 0.79 ± 0.00 a | 14.6 ± 0.7 g | 6.50 ± 0.71 f | 88.3 ± 0.11 g | 5.83 ± 0.41 e |
| Maize | ||||||||
| MF | 12.2 ± 0.31 e | 0.74 ± 0.04 a | 6.42 ± 0.21 a | 1.66 ± 0.02 ab | 79.0 ± 0.58 f | 76.0 ± 2.26 e | 7.21 ± 0.25 c | 2.00 ± 0.40 a |
| Stage of Pasting | ||||||
|---|---|---|---|---|---|---|
| Initial Viscosity (mPa∙s) | Peak Viscosity (Pa∙s) | Peak Time (min) | End of Holding at 95 °C (Pa∙s) | End of Cooling to 50 °C (Pa∙s) | Final Paste at 50 °C (Pa∙s) | |
| Quinoa | ||||||
| QWGF | 18.4 ± 0.76 a,b | 6.25 ± 0.18 e | 12.5 | 2.74 ± 0.34 e | 6.07 ± 0.30 e | 5.83 ± 0.43 e |
| QDF | 24.8 ± 1.05 b,c | 4.39 ± 0.16 d | 12.5 | 2.41 ± 0.24 c | 4.68 ± 0.19 c | 4.40 ± 0.22 c |
| QPRF | 24.1 ± 0.12 b,c | 0.91 ± 0.03 ab | 12.5 | 0.63 ± 0.01 a | 0.75 ± 0.01 a | 0.67 ± 0.01 a |
| Amaranth | ||||||
| AWGF | 29.1 ± 1.32 c | 1.92 ± 0.07 b,c | 10.3 | 0.98 ± 0.03 b | 1.64 ± 0.06 b | 1.72 ± 0.07 b |
| APRF | 26.4 ± 0.84 c | 0.29 ± 0.03 a | 12.5 | 0.13 ± 0.01 a | 0.19 ± 0.01 a | 0.19 ± 0.01 a |
| Buckwheat | ||||||
| BDF | 48.4 ± 2.64 e | 9.60 ± 0.46 f | 12.1 | 4.84 ± 0.90 f | 12.5 ± 0.47 f | 13.0 ± 0.35 f |
| BPRF | 37.5 ± 2.21 d | 2.81 ± 0.10 c | 12.5 | 2.23 ± 0.07 d | 5.35 ± 0.24 d | 5.92 ± 0.24 e |
| Rice | ||||||
| RF | 15.5 ± 0.06 a | 9.37 ± 1.43 f | 11.2 | 2.28 ± 0.23 c | 4.24 ± 0.15 c | 4.31 ± 0.13 c |
| RPC | 17.9 ± 0.13 a | n.d. | n.d. | 0.02 ± 0.00 g | 0.02 ± 0.00 g | 0.02 ± 0.00 g |
| Maize | ||||||
| MF | 16.4 ± 0.02 a | 7.11 ± 0.20 e | 10.6 | 1.54 ± 0.15 cd | 4.67 ± 0.30 cd | 5.07 ± 0.33 d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso-Miravalles, L.; O’Mahony, J.A. Composition, Protein Profile and Rheological Properties of Pseudocereal-Based Protein-Rich Ingredients. Foods 2018, 7, 73. https://doi.org/10.3390/foods7050073
Alonso-Miravalles L, O’Mahony JA. Composition, Protein Profile and Rheological Properties of Pseudocereal-Based Protein-Rich Ingredients. Foods. 2018; 7(5):73. https://doi.org/10.3390/foods7050073
Chicago/Turabian StyleAlonso-Miravalles, Loreto, and James A. O’Mahony. 2018. "Composition, Protein Profile and Rheological Properties of Pseudocereal-Based Protein-Rich Ingredients" Foods 7, no. 5: 73. https://doi.org/10.3390/foods7050073
APA StyleAlonso-Miravalles, L., & O’Mahony, J. A. (2018). Composition, Protein Profile and Rheological Properties of Pseudocereal-Based Protein-Rich Ingredients. Foods, 7(5), 73. https://doi.org/10.3390/foods7050073