
Stimulatory Effects of Cinnamon Extract (Cinnamomum cassia) during the Initiation Stage of 3T3-L1 Adipocyte Differentiation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Cell Culture
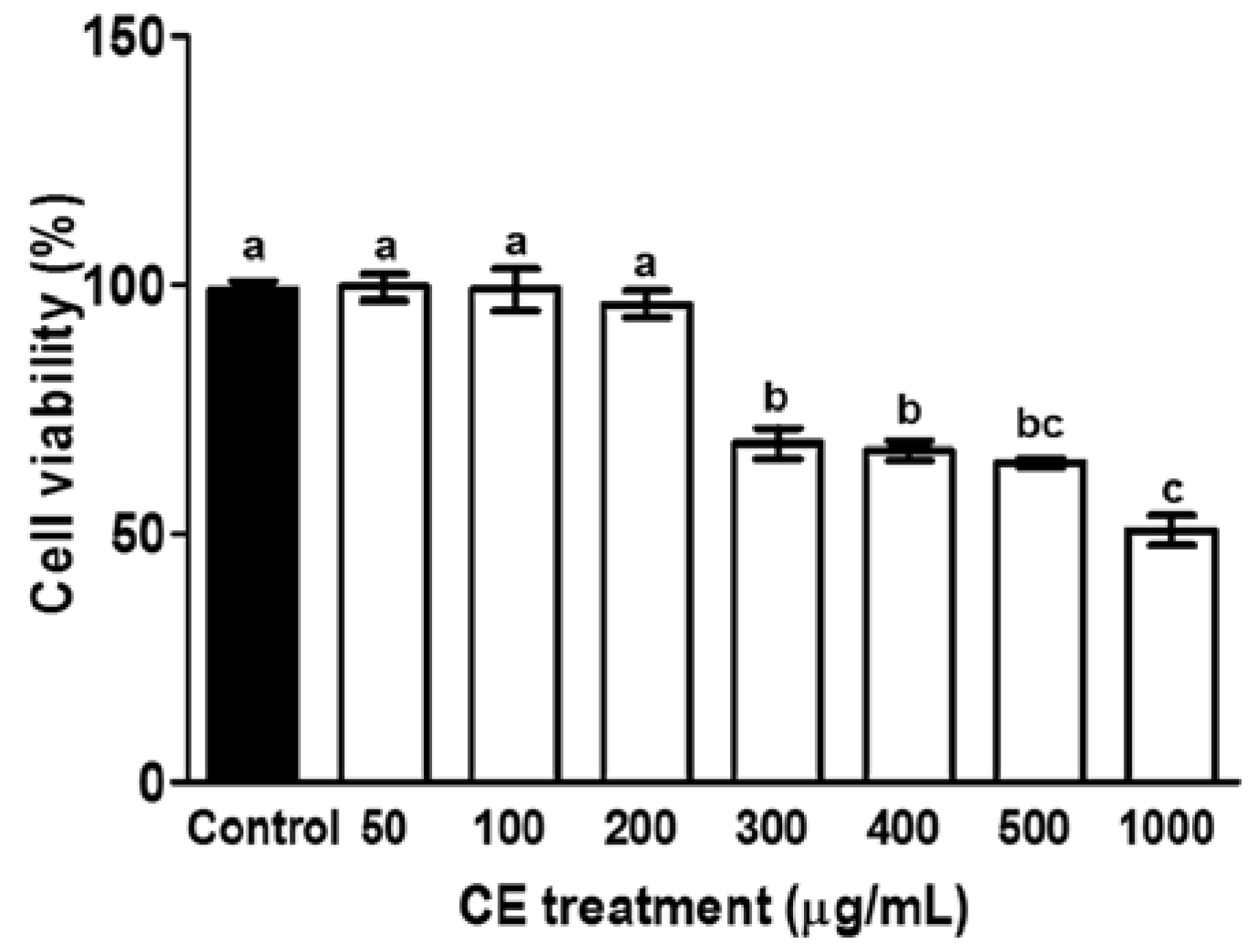
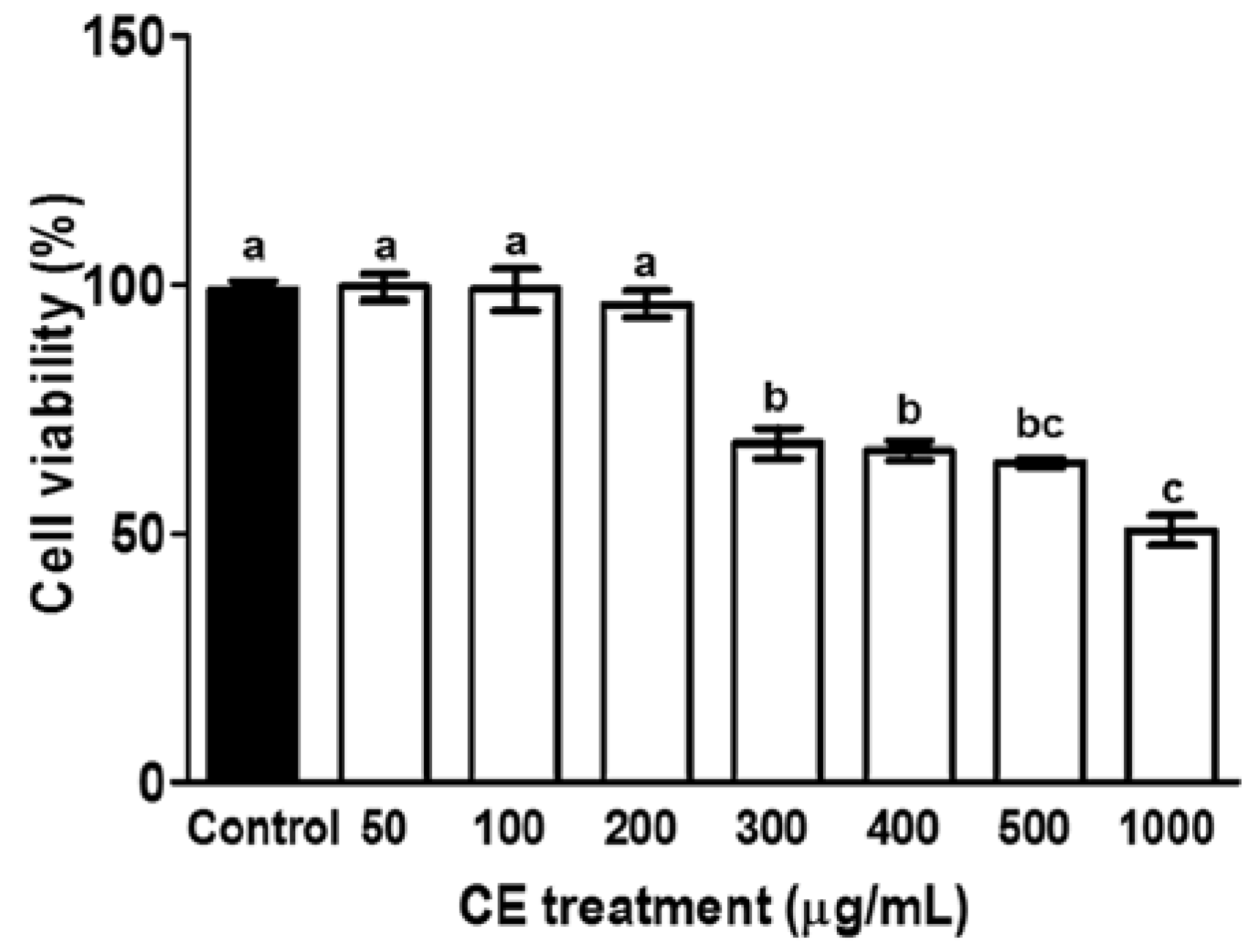
2.3. Cell Viability Assay
2.4. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.5. Oil-Red O Staining
2.6. Data Analysis
3. Results
3.1. Cytotoxicity of CE in Preadipocytes
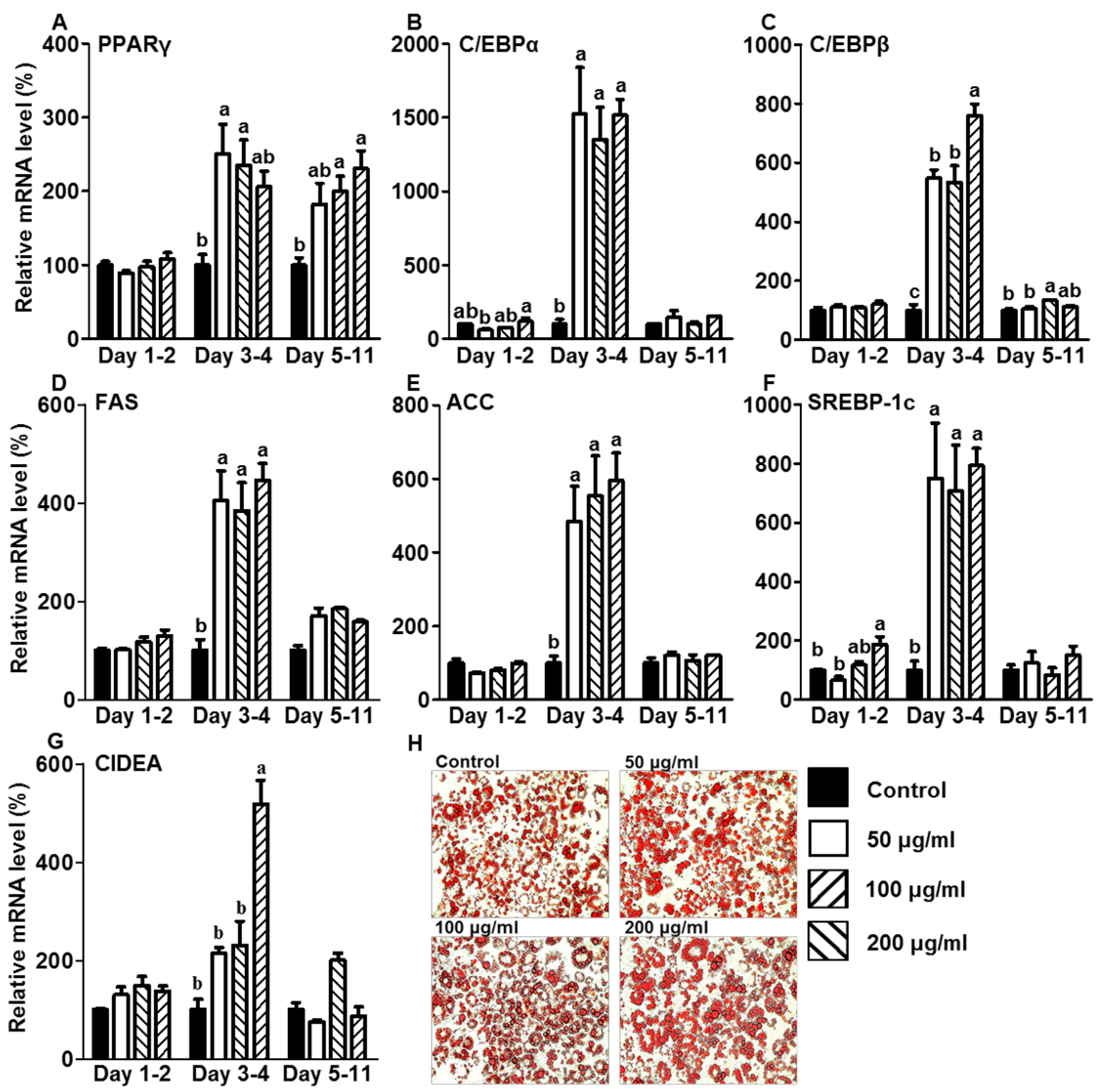
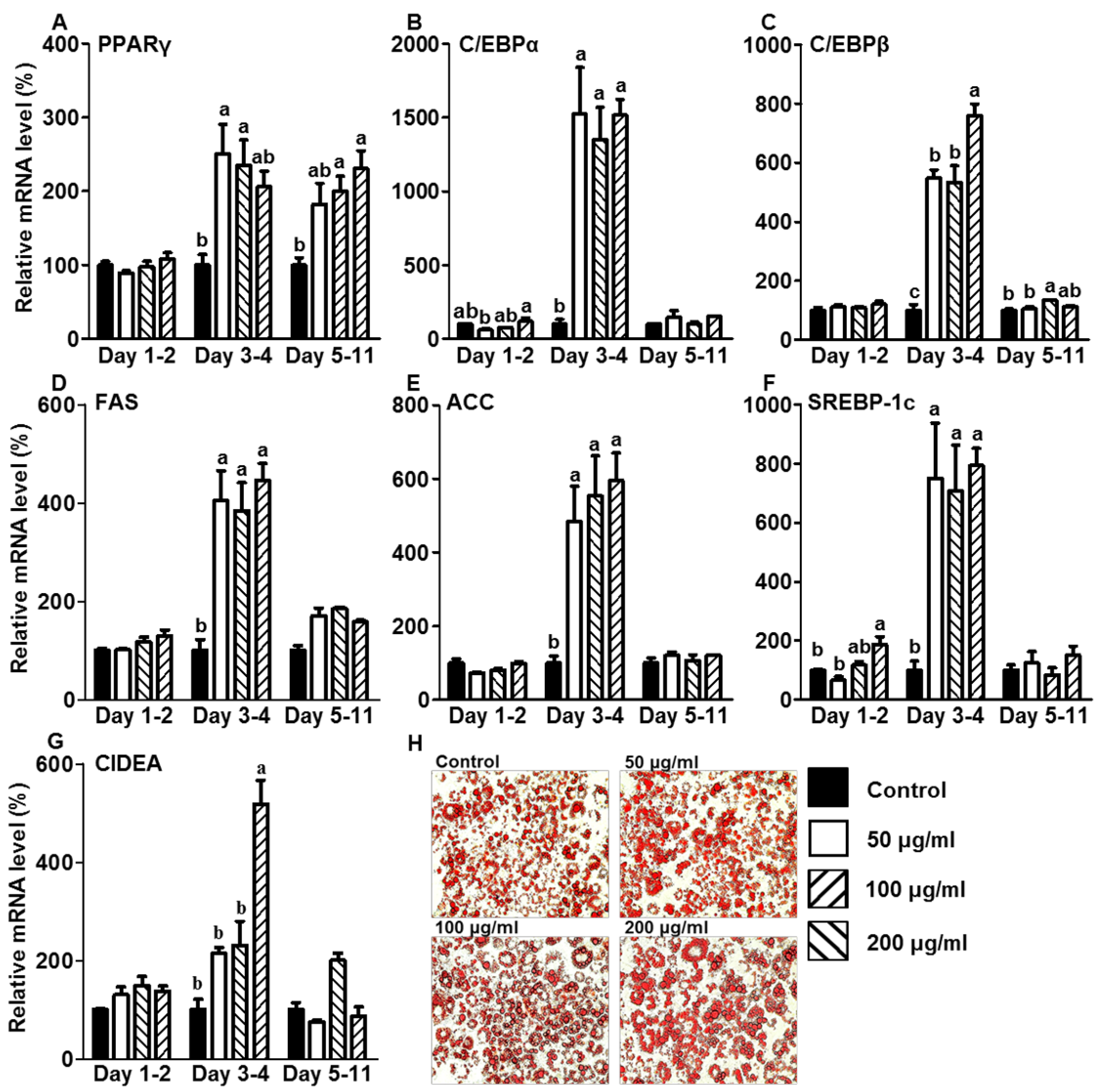
3.2. CE Increased Lipid Accumulation by Increasing the Expression of Adipogenic and Lipogenic Genes during the Initiation Stage of Differentiation
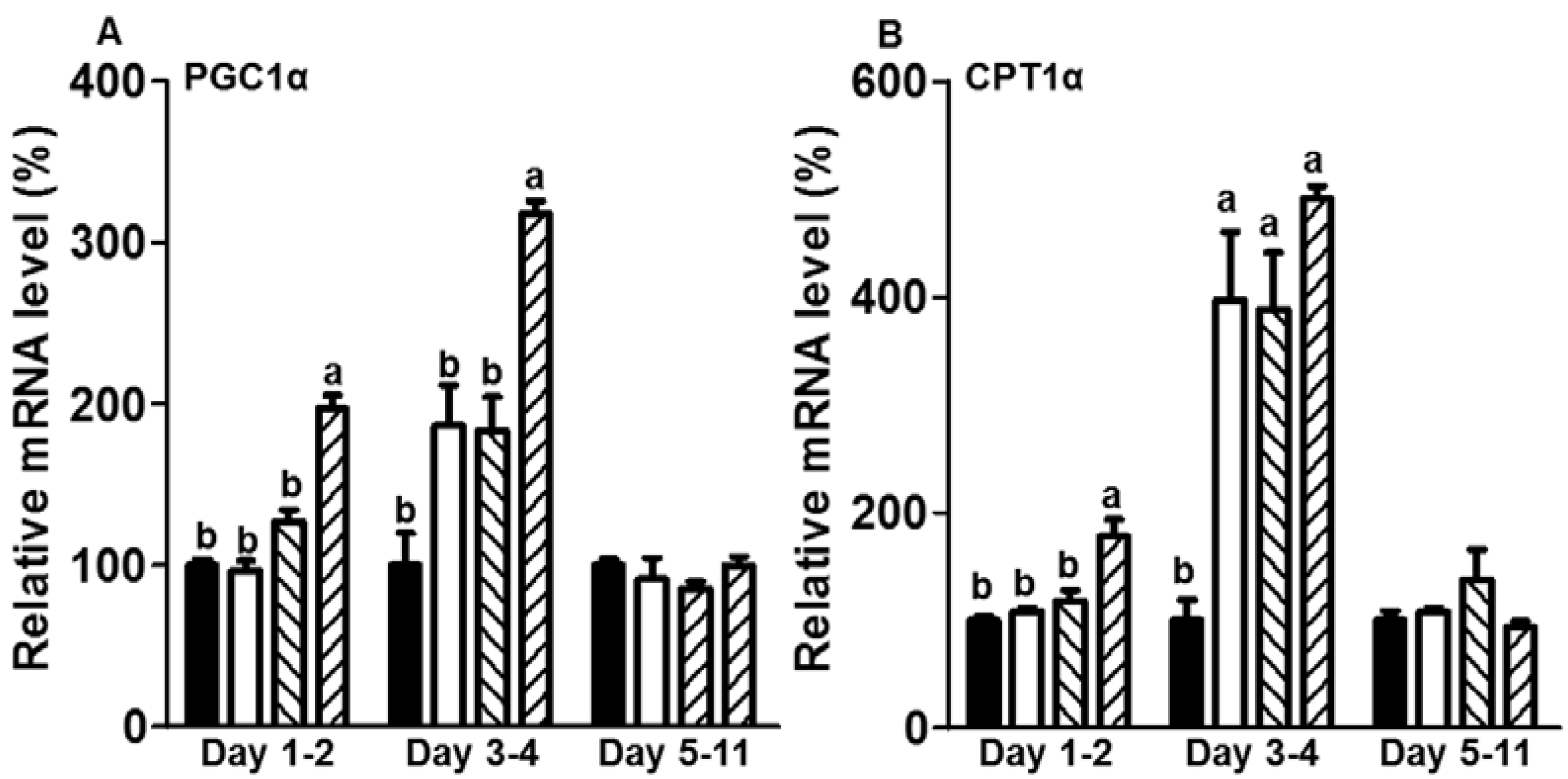
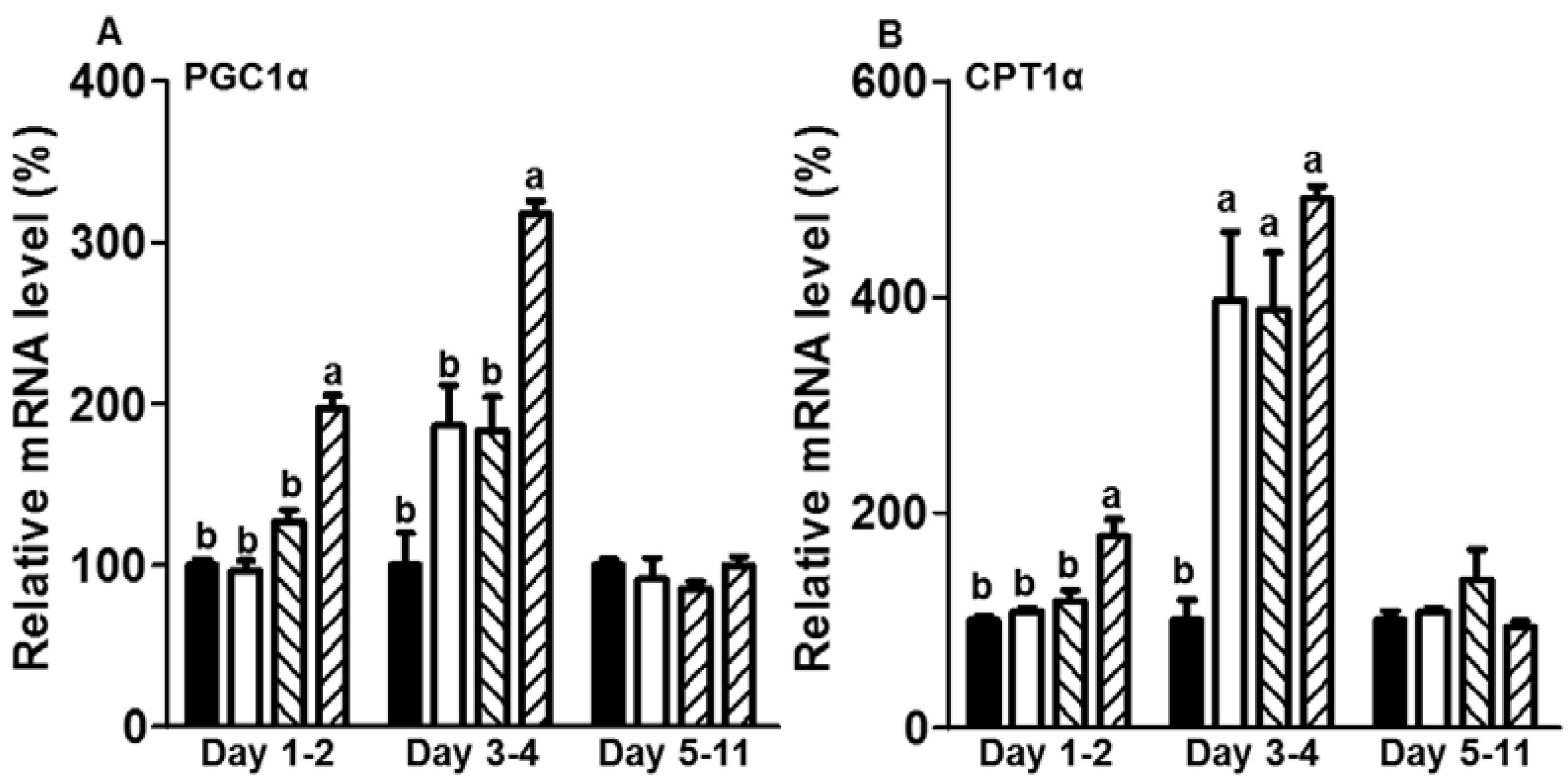
3.3. CE Elevated the mRNA Expression Levels of Genes Related to Fatty Acid Oxidation in the Initiation Stage of Differentiation during Adipogenesis
4. Discussion
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Schuster, D.P. Obesity and the development of type 2 diabetes: The effects of fatty tissue inflammation. Diabetes Metab. Syndr. Obes. 2010, 3, 253–262. [Google Scholar] [CrossRef] [PubMed]
- Konige, M.; Wang, H.; Sztalryd, C. Role of adipose specific lipid droplet proteins in maintaining whole body energy homeostasis. Biochim. Biophys. Acta 2014, 1842, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef] [PubMed]
- Sears, B.; Perry, M. The role of fatty acids in insulin resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Gregoire, F.M.; Smas, C.M.; Sul, H.S. Understanding adipocyte differentiation. Physiol. Rev. 1998, 78, 783–809. [Google Scholar] [PubMed]
- Bermudez, V.; Finol, F.; Parra, N.; Parra, M.; Perez, A.; Penaranda, L.; Vilchez, D.; Rojas, J.; Arraiz, N.; Velasco, M. Ppar-gamma agonists and their role in type 2 diabetes mellitus management. Am. J. Ther. 2010, 17, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Hauner, H. The mode of action of thiazolidinediones. Diabetes Metab. Res. Rev. 2002, 18 (Suppl. 2), S10–S15. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Waltenberger, B.; Pferschy-Wenzig, E.M.; Blunder, M.; Liu, X.; Malainer, C.; Blazevic, T.; Schwaiger, S.; Rollinger, J.M.; Heiss, E.H.; et al. Natural product agonists of peroxisome proliferator-activated receptor gamma (ppargamma): A review. Biochem. Pharmacol. 2014, 92, 73–89. [Google Scholar] [CrossRef] [PubMed]
- Sangal, A. Role of cinnamon as beneficial antidiabetic food adjunct: A review. Adv. Appl. Sci. Res. 2011, 2, 440–450. [Google Scholar]
- Khan, A.; Sadafar, M.; Ali Khan, M.M.; Khattak, K.N.; Anderson, R.A. Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care 2003, 26, 3215–3218. [Google Scholar] [CrossRef] [PubMed]
- Crawford, P. Effectiveness of cinnamon for lowering hemoglobin A1C in patients with type 2 diabetes: A randomized, controlled trial. J. Am. Board Fam. Med. 2009, 22, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Li, J.E.; Futawaka, K.; Yamamoto, H.; Kasahara, M.; Tagami, T.; Liu, T.H.; Moriyama, K. Cinnamaldehyde contributes to insulin sensitivity by activating PPARdelta, PPARgamma, and RXR. Am. J. Chin. Med. 2015, 43, 879–892. [Google Scholar] [CrossRef] [PubMed]
- Hasanzade, F.; Toliat, M.; Emami, S.A.; Emamimoghaadam, Z. The effect of cinnamon on glucose of type ii diabetes patients. J. Tradit. Complement. Med. 2013, 3, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Jung, H.W.; Bae, H.S.; Kang, J.S.; Park, Y.K. The extract of cinnamomum cassia twigs inhibits adipocyte differentiation via activation of the insulin signaling pathway in 3T3-L1 preadipocytes. Pharm. Biol. 2013, 51, 961–967. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Yuan, H.D.; Kim, D.Y.; Quan, H.Y.; Chung, S.H. Cinnamaldehyde prevents adipocyte differentiation and adipogenesis via regulation of peroxisome proliferator-activated receptor-gamma (ppargamma) and amp-activated protein kinase (AMPK) pathways. J. Agric. Food Chem. 2011, 59, 3666–3673. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Spiegelman, B.M. Molecular regulation of adipogenesis. Annu. Rev. Cell Dev. Biol. 2000, 16, 145–171. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.M.; Dzamko, N.; Cleasby, M.E.; Hegarty, B.D.; Furler, S.M.; Cooney, G.J.; Kraegen, E.W. Direct demonstration of lipid sequestration as a mechanism by which rosiglitazone prevents fatty-acid-induced insulin resistance in the rat: Comparison with metformin. Diabetologia 2004, 47, 1306–1313. [Google Scholar] [CrossRef] [PubMed]
- Sheng, X.; Zhang, Y.; Gong, Z.; Huang, C.; Zang, Y.Q. Improved insulin resistance and lipid metabolism by cinnamon extract through activation of peroxisome proliferator-activated receptors. PPAR Res. 2008, 2008, 581348. [Google Scholar] [CrossRef] [PubMed]
- Puri, V.; Ranjit, S.; Konda, S.; Nicoloro, S.M.; Straubhaar, J.; Chawla, A.; Chouinard, M.; Lin, C.; Burkart, A.; Corvera, S.; et al. Cidea is associated with lipid droplets and insulin sensitivity in humans. Proc. Natl. Acad. Sci. USA 2008, 105, 7833–7838. [Google Scholar] [CrossRef] [PubMed]
- Barneda, D.; Planas-Iglesias, J.; Gaspar, M.L.; Mohammadyani, D.; Prasannan, S.; Dormann, D.; Han, G.S.; Jesch, S.A.; Carman, G.M.; Kagan, V.; et al. The brown adipocyte protein cidea promotes lipid droplet fusion via a phosphatidic acid-binding amphipathic helix. eLife 2015, 4, e07485. [Google Scholar] [CrossRef] [PubMed]
- Abreu-Vieira, G.; Fischer, A.W.; Mattsson, C.; de Jong, J.M.; Shabalina, I.G.; Rydén, M.; Laurencikiene, J.; Arner, P.; Cannon, B.; Nedergaard, J.; et al. Cidea improves the metabolic profile through expansion of adipose tissue. Nat. Commun. 2015, 6, 7433. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Yon Toh, S.; Chen, Z.; Guo, K.; Ng, C.P.; Ponniah, S.; Lin, S.C.; Hong, W.; Li, P. CIDEA-deficient mice have lean phenotype and are resistant to obesity. Nat. Genet. 2003, 35, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Hallberg, M.; Morganstein, D.L.; Kiskinis, E.; Shah, K.; Kralli, A.; Dilworth, S.M.; White, R.; Parker, M.G.; Christian, M. A functional interaction between RIP140 and PGC-1alpha regulates the expression of the lipid droplet protein CIDEA. Mol. Cell Biol. 2008, 28, 6785–6795. [Google Scholar] [CrossRef] [PubMed]
- Karamanlidis, G.; Karamitri, A.; Docherty, K.; Hazlerigg, D.G.; Lomax, M.A. C/EBPbeta reprograms white 3T3-L1 preadipocytes to a brown adipocyte pattern of gene expression. J. Biol. Chem. 2007, 282, 24660–24669. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer | Reverse Primer |
|---|---|---|
| ACC | TGCATTCTGACCTTCACGAC | ACATCCACTTCCACACACGA |
| C/EBPα | GGACAAGAACAGCAACGAGTA | GCAGTTGCCATGGCCTTGA |
| C/EBPβ | TGGACAAGCTGAGCGACGAG | TGTGCTGCGTCTCCAGGTTG |
| CIDEA | ATCACAACTGGCCTGGTTACG | TACTACCCGGTGTCCATTTCT |
| CPT1α | TTTGACTTTGAGAAATACCCTGATA | TGGATGAAATTCTCTCCCACAATAA |
| FAS | TGGGTTCTAGCCAGCAGAGT | ACCACCAGAGACCGTTATGC |
| PGC1α | TGCCCAGATCTTCCTGAACT | TCTGTGAGAACCGCTAGCAA |
| PPARγ | TTTGACTTTGAGAAATACCC | TGGATGAAATTCTCTCCAC |
| RPL32 | CACCAGTCAGACCGATAT | TTCTCCGCACCCTGTTG |
| SREBP-1c | GAACAGACACTGGCCGAGAT | GAGGCCAGAGAAGCAGAAGAG |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.G.; Siaw, J.A.; Kang, H.W. Stimulatory Effects of Cinnamon Extract (Cinnamomum cassia) during the Initiation Stage of 3T3-L1 Adipocyte Differentiation. Foods 2016, 5, 83. https://doi.org/10.3390/foods5040083
Lee SG, Siaw JA, Kang HW. Stimulatory Effects of Cinnamon Extract (Cinnamomum cassia) during the Initiation Stage of 3T3-L1 Adipocyte Differentiation. Foods. 2016; 5(4):83. https://doi.org/10.3390/foods5040083
Chicago/Turabian StyleLee, Sang Gil, Joanna A. Siaw, and Hye Won Kang. 2016. "Stimulatory Effects of Cinnamon Extract (Cinnamomum cassia) during the Initiation Stage of 3T3-L1 Adipocyte Differentiation" Foods 5, no. 4: 83. https://doi.org/10.3390/foods5040083
APA StyleLee, S. G., Siaw, J. A., & Kang, H. W. (2016). Stimulatory Effects of Cinnamon Extract (Cinnamomum cassia) during the Initiation Stage of 3T3-L1 Adipocyte Differentiation. Foods, 5(4), 83. https://doi.org/10.3390/foods5040083