Comparison of Growth Kinetics of Various Pathogenic E. coli on Fresh Perilla Leaf


Abstract
:1. Introduction
2. Experimental Section
2.1. Bacterial Strains
2.2. Inoculation of Strain in Perilla Leaves
2.3. Primary Modeling
2.4. Secondary Modeling

2.5. Performance Evaluation of Perilla Leaves Model
2.6. Statistical Analysis
3. Results and Discussion
3.1. Development of Growth Model for E. coli O157:H7 in Perilla Leaves

{kind=link}
| Parameter | 13 °C | 17 °C | 24 °C | 30 °C | 36 °C | Secondary model equation | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | ||
| LT x | 45.84 e | 0.16 | 20.40 d | 0.23 | 4.80 c | 0.10 | 2.64 b | 0.10 | 2.40 a | 0.32 | LT = 0.6688 + (−42.71/T) + (763.7/T2) |
| SGR y | 0.034 a | 0.00 | 0.055 b | 0.01 | 0.236 c | 0.02 | 0.359 d | 0.05 | 0.548 e | 0.00 | SGR = [0.1183(T − 5.182)]2 |
| MPD z | 5.73 a | 0.11 | 5.81 a | 0.10 | 6.73 b | 0.12 | 7.01 b | 0.03 | 7.02 b | 0.04 | MPD = 3.717 + 0.1769T − 0.002378T2 |
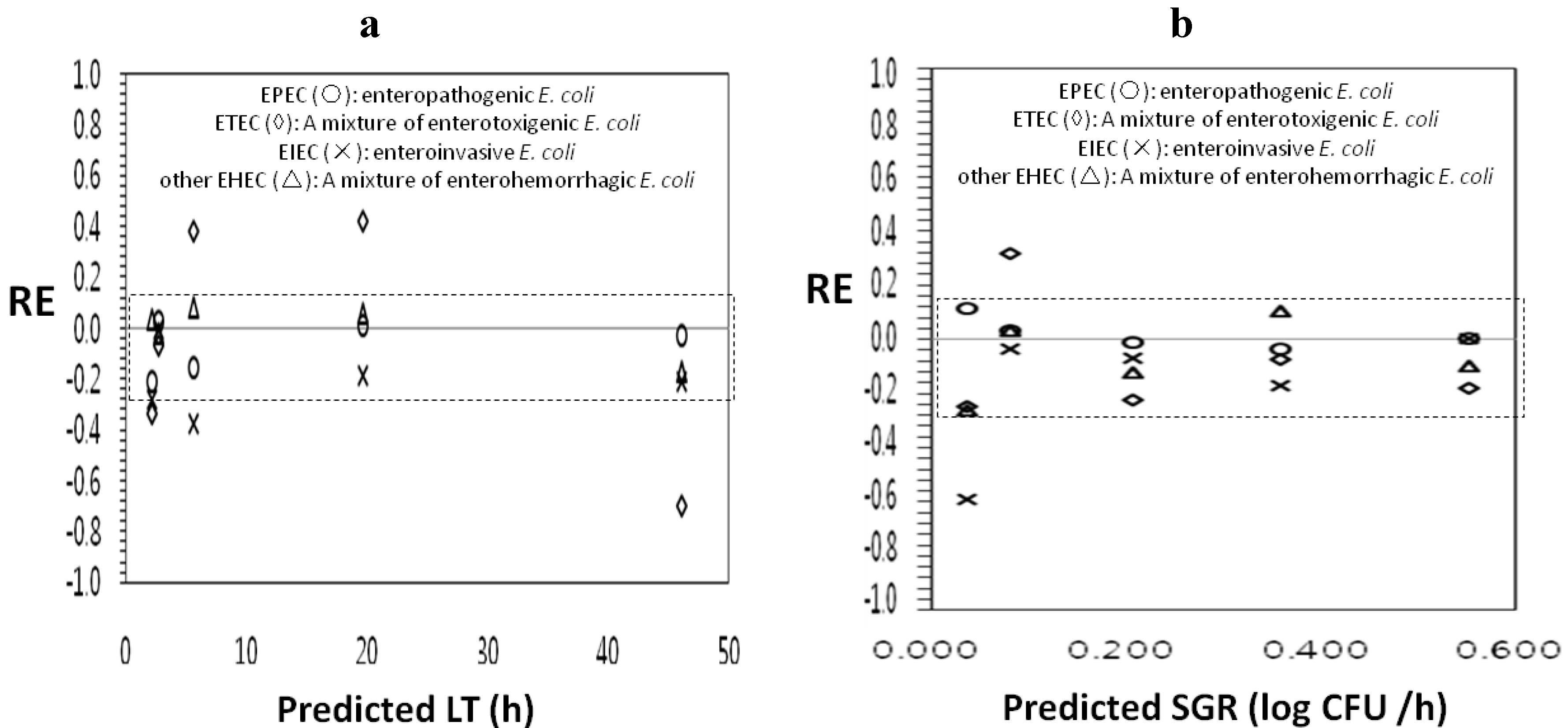
3.2. Evaluation of Model Performance
| Parameter | Strains | 13 °C | 17 °C | 24 °C | 30 °C | 36 °C | |||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | ||
| LT x | O157:H7 | 45.84 a | 0.00 | 19.68 d | 0.00 | 5.52 b | 0.00 | 2.64 a | 0.00 | 2.16 a | 0.00 |
| EPEC k | 47.52 b | 1.18 | 19.44 c | 1.76 | 6.48 b | 0.43 | 2.73 a | 0.06 | 2.64 b | 0.01 | |
| ETEC l | 78.48 e | 0.85 | 11.52 a | 0.29 | 3.60 a | 0.30 | 2.89 b | 0.20 | 2.88 c | 0.07 | |
| EIEC m | 55.92 c | 1.06 | 23.28 e | 0.55 | 7.68 c | 0.01 | 2.74 a | 0.11 | 2.87 c | 0.04 | |
| EHEC n | 54.24 d | 1.42 | 18.48 b | 0.24 | 5.04 b | 0.05 | 2.88 b | 0.07 | 2.12 a | 0.04 | |
| SGR y | O157:H7 | 0.036 d | 0.00 | 0.081 b | 0.00 | 0.207 e | 0.00 | 0.359 d | 0.00 | 0.554 c | 0.00 |
| EPEC | 0.040 e | 0.01 | 0.084 c | 0.00 | 0.204 d | 0.01 | 0.347 c | 0.02 | 0.555 d | 0.07 | |
| ETEC | 0.027 c | 0.00 | 0.107 d | 0.01 | 0.161 a | 0.00 | 0.332 b | 0.01 | 0.455 a | 0.04 | |
| EIEC | 0.015 a | 0.00 | 0.079 a | 0.09 | 0.192 c | 0.01 | 0.298 a | 0.01 | 0.555 d | 0.00 | |
| EHEC | 0.026 b | 0.02 | 0.084 c | 0.00 | 0.182 b | 0.04 | 0.396 e | 0.02 | 0.499 b | 0.02 | |
| Strain | Model | Bfa | MRE b | Afc | MARE d | %RE e |
|---|---|---|---|---|---|---|
| EPEC f | LT j | 0.94 | −0.03 | 1.08 | 0.09 | 100 |
| SGR k | 1.02 | 0.14 | 1.04 | 0.04 | 100 | |
| ETEC g | LT | 1.03 | −0.07 | 1.46 | 0.38 | 33.3 |
| SGR | 0.90 | −17.92 | 1.24 | 0.21 | 83.3 | |
| EIEC h | LT | 0.83 | −0.21 | 1.21 | 0.21 | 66.7 |
| SGR | 0.79 | −7.20 | 1.27 | 0.17 | 83.3 | |
| EHEC i | LT | 1.00 | 0.04 | 1.08 | 0.08 | 100 |
| SGR | 0.92 | −9.87 | 1.14 | 0.12 | 100 |
4. Conclusions
Conflicts of Interest
References
- Myron, M.L. Escherichia coli that cause diarrhea: Enterotoxigenic, enteropathogenic, enteroinvasive, enterohemorrhagic, and enteroadherent. J. Infect. Dis. 1987, 155, 377–389. [Google Scholar] [CrossRef]
- Cooley, M.; Carychao, D.; Crawford-Miksza, L.; Jay, M.T.; Myers, C.; Rose, C.; Keys, C.; Farrar, J.; Mandrell, R.E. Incidence and tracking of Escherichia coli O157:H7 in a major produce production region in California. PloS One 2007, 2, e1159. [Google Scholar] [CrossRef]
- News, F.Q. Germany Finally Confirms Source of Deadly E. coli Outbreak. Available online: http://www.foodproductiondaily.com/Safety-Regulation/Germany-finally-confirms-source-of-deadly-E.coli-outbreak (accessed on 15 March 2013).
- Korea Centers for Disease Control & Prevention. Available online: http://www.cdc.go.kr/CDC/eng/main.jsp (accessed on 20 Febuary 2013).
- Lee, J.K.; Park, I.H.; Yoon, K.; Kim, H.J.; Cho, J.I.; Lee, S.H.; Hwang, I.G. An analysis of epidemiological investigation reports regarding to pathogenic E. coli outbreaks in Korea from 2009 to 2010. J. Food Hyg. Saf. 2012, 27, 366–374. [Google Scholar]
- Jinneman, K.C.; Trost, P.A.; Hill, W.E.; Weagant, S.D.; Bryant, J.L.; Kaysner, C.A.; Wekell, M.M. Comparison of template preparation methods from foods for amplification of Escherichia coli O157 Shiga-like toxins type I and II DNA by multiplex polymerase chain reaction. J. Food Prot. 1995, 58, 722–726. [Google Scholar]
- Kwon, W.H.; Lee, W.G.; Song, J.E.; Kim, K.Y.; Shim, W.B.; Yoon, Y.H.; Kim, Y.S.; Chung, D.H. Microbiological hazard analysis on perilla leaf farms at the harvesting stage for the application of the Good Agricultural Practices (GAP). J. Food Hyg. Saf. 2012, 27, 295–300. [Google Scholar]
- Choi, J.W.; Park, S.Y.; Yeon, J.H.; Lee, M.J.; Chung, D.H.; Lee, K.H.; Kim, M.G.; Lee, D.H.; Kim, K.S.; Ha, S.D. Microbial contamination levels of fresh vegetables distributed in markets. J. Food Hyg. Saf. 2005, 20, 43–47. [Google Scholar]
- Jung, S.H.; Hur, M.J.; Ju, J.H.; Kim, K.A.; Oh, S.S.; Go, S.M.; Kim, Y.H.; Im, J.S. Microbiological evaluation of raw vegetable. J. Food Hyg. Saf. 2006, 24, 250–257. [Google Scholar]
- Park, S.Y.; Choi, J.W.; Chung, D.H.; Kim, M.G.; Lee, K.H.; Kim, K.S.; Bahk, G.J.; Bae, D.H.; Park, S.K.; Kim, K.Y. Development of a predictive mathematical model for the growth kinetics of Listeria monocytogenes in sesame leaves. Food Sci. Biotechnol. 2007, 16, 238–242. [Google Scholar]
- Oscar, T.P. Extrapolation of a predictive model for growth of a low inoculum size of Salmonella typhimurium DT104 on chicken skin to higher inoculum sizes. J. Food Prot. 2011, 74, 1630–1638. [Google Scholar] [CrossRef]
- Buchanan, R.L.; Bagi, L.K.; Goins, R.V.; Phillips, J.G. Response surface models for the growth kinetics of Escherichia coli O157:H7. Food Microbiol. 1993, 10, 303–315. [Google Scholar] [CrossRef]
- Kovárová, K.; Zehnder, A.J.; Egli, T. Temperature-dependent growth kinetics of Escherichia coli ML 30 in glucose-limited continuous culture. J. Bacteriol. 1996, 178, 4530–4539. [Google Scholar]
- Presser, K.A.; Ratkowsky, D.A.; Ross, T. Modelling the growth rate of Escherichia coli as a function of pH and lactic acid concentration. Appl. Environ. Microbiol. 1997, 63, 2355–2360. [Google Scholar]
- Ross, T.; Ratkowsky, D.A.; Mellefont, L.A.; McMeekin, T.A. Modelling the effects of temperature, water activity, pH and lactic acid concentration on the growth rate of Escherichia coli. Int. J. Food Microbiol. 2003, 82, 33–43. [Google Scholar] [CrossRef]
- Sutherland, J.P.; Bayliss, A.J.; Braxton, D.S.; Beaumont, A.L. Predictive modelling of Escherichia coli O157:H7: Inclusion of carbon dioxide as a fourth factor in a pre-existing model. Int. J. Food Microbiol. 1997, 37, 113–120. [Google Scholar] [CrossRef]
- Sutherland, J.P.; Bayliss, A.J.; Braxton, D.S. Predictive modelling of growth of Escherichia coli O157:H7: The effects of temperature, pH and sodium chloride. Int. J. Food Microbiol. 1995, 25, 29–49. [Google Scholar] [CrossRef]
- Koseki, S.; Isobe, S. Prediction of pathogen growth on iceberg lettuce under real temperature history during distribution from farm to table. Int. J. Food Microbiol. 2005, 104, 239–248. [Google Scholar] [CrossRef]
- McKellar, R.C.; Delaquis, P. Development of a dynamic growth-death model for Escherichia coli O157:H7 in minimally processed leafy green vegetables. Int. J. Food Microbiol. 2011, 151, 7–14. [Google Scholar] [CrossRef]
- Daughtry, B.J.; Davey, K.R.; King, K.D. Temperature dependence of growth kinetics of food bacteria. Food Microbiol. 1997, 14, 21–30. [Google Scholar] [CrossRef]
- Oscar, T.P. Development and validation of a tertiary simulation model for predicting the potential growth of Salmonella typhimurium on cooked chicken. Int. J. Food Microbiol. 2002, 76, 177–190. [Google Scholar] [CrossRef]
- Ratkowsky, D.A.; Olley, J.; McMeekin, T.A.; Ball, A. Relationship between temperature and growth rate of bacterial cultures. J. Bacteriol. 1982, 149, 1–5. [Google Scholar]
- McMeekin, T.A.; Olley, J.; Ross, T. Predictive Microbiology: Theory and Application; John Wiley & Sons Ltd.: Taunton, UK, 1993. [Google Scholar]
- Ross, T. Indices for performance evaluation of predictive models in food microbiology. J. Appl. Microbiol. 1996, 81, 501–508. [Google Scholar]
- Oscar, T.P. Validation of lag time and growth rate models for Salmonella typhimurium: Acceptable prediction zone method. J. Food Sci. 2005, 70, 129–137. [Google Scholar] [CrossRef]
- Abou-Zeid, K.A.; Oscar, T.P.; Schwarz, J.G.; Hashem, F.M.; Whiting, R.C.; Yoon, K. Development and validation of a predictive model for Listeria monocytogenes Scott A as a function of temperature, pH, and commercial mixture of potassium lactate and sodium diacetate. J. Mircobiol. Biotechnol. 2009, 19, 718–726. [Google Scholar]
- Delignette-Muller, M.L.; Rosso, L.; Flandrois, J.P. Accuracy of microbial growth predictions with square root and polynomial models. Int. J. Food Microbiol. 1995, 27, 139–146. [Google Scholar] [CrossRef]
- McKellar, R.C.; Lu, X. Modeling Microbial Responses in Food; CRC Press: Boca Raton, FL, USA, 2003. [Google Scholar]
- Salter, M.A.; Ross, T.; McMeekin, T.A. Applicability of a model for non-pathogenic Escherichia coli for predicting the growth of pathogenic Escherichia coli. J. Appl. Microbiol. 1998, 85, 357–364. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kim, J.; Ro, E.; Yoon, K. Comparison of Growth Kinetics of Various Pathogenic E. coli on Fresh Perilla Leaf. Foods 2013, 2, 364-373. https://doi.org/10.3390/foods2030364
Kim J, Ro E, Yoon K. Comparison of Growth Kinetics of Various Pathogenic E. coli on Fresh Perilla Leaf. Foods. 2013; 2(3):364-373. https://doi.org/10.3390/foods2030364
Chicago/Turabian StyleKim, Juhui, Eunyoung Ro, and Kisun Yoon. 2013. "Comparison of Growth Kinetics of Various Pathogenic E. coli on Fresh Perilla Leaf" Foods 2, no. 3: 364-373. https://doi.org/10.3390/foods2030364
APA StyleKim, J., Ro, E., & Yoon, K. (2013). Comparison of Growth Kinetics of Various Pathogenic E. coli on Fresh Perilla Leaf. Foods, 2(3), 364-373. https://doi.org/10.3390/foods2030364