
The Transcriptome Analysis Provides New Insights into Signaling for Bamboo Shoot Development of Sympodial Bamboo


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction, Library Construction, and RNA Sequencing
2.3. Functional Annotation and Enrichment Analysis of Differentially Expressed Genes
2.4. Differential Expression Analysis
2.5. WGCNA Analysis
2.6. RT-qPCR Analysis
3. Result
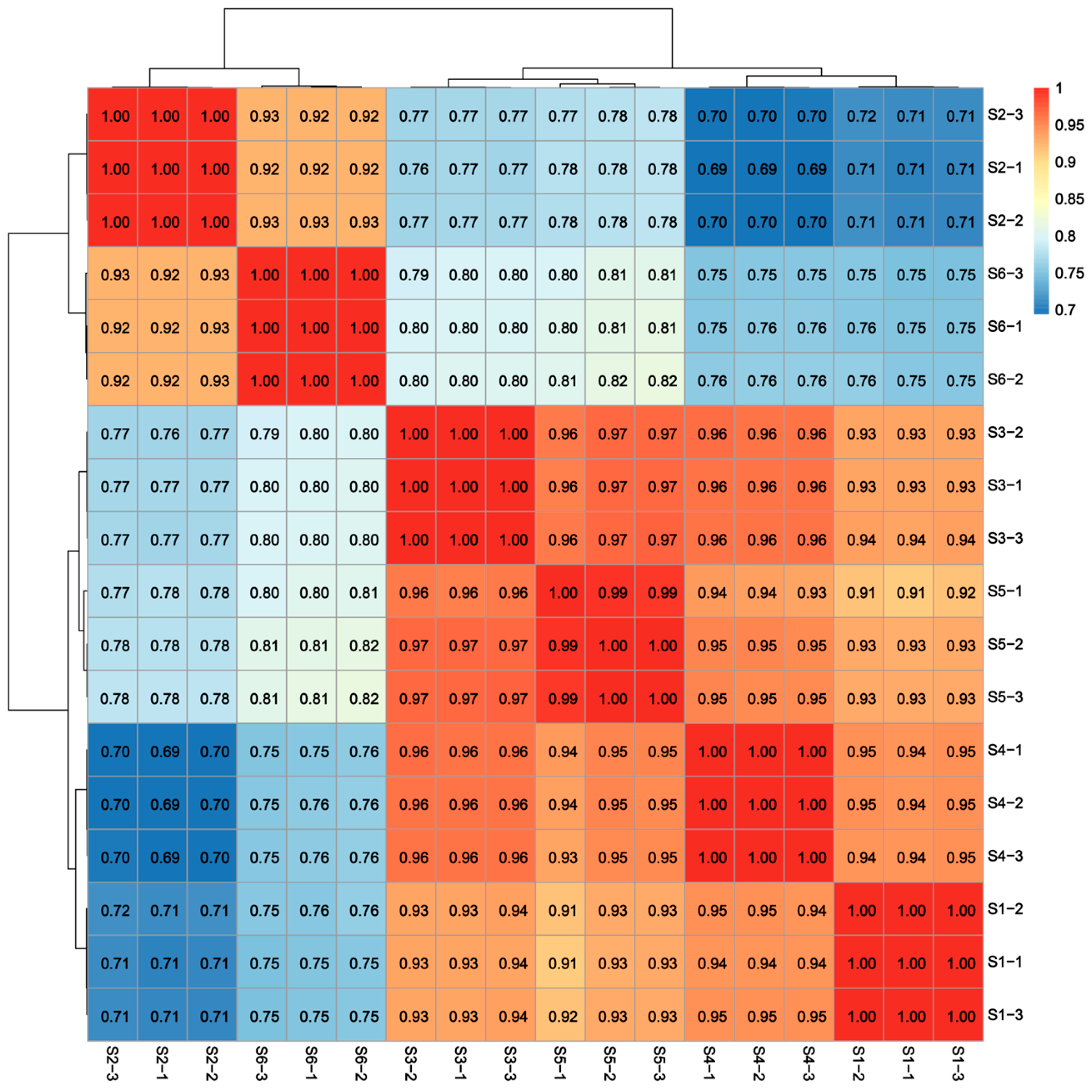
3.1. Transcriptome Data Analysis
3.2. Function Annotation of Bamboo Shoot Tips
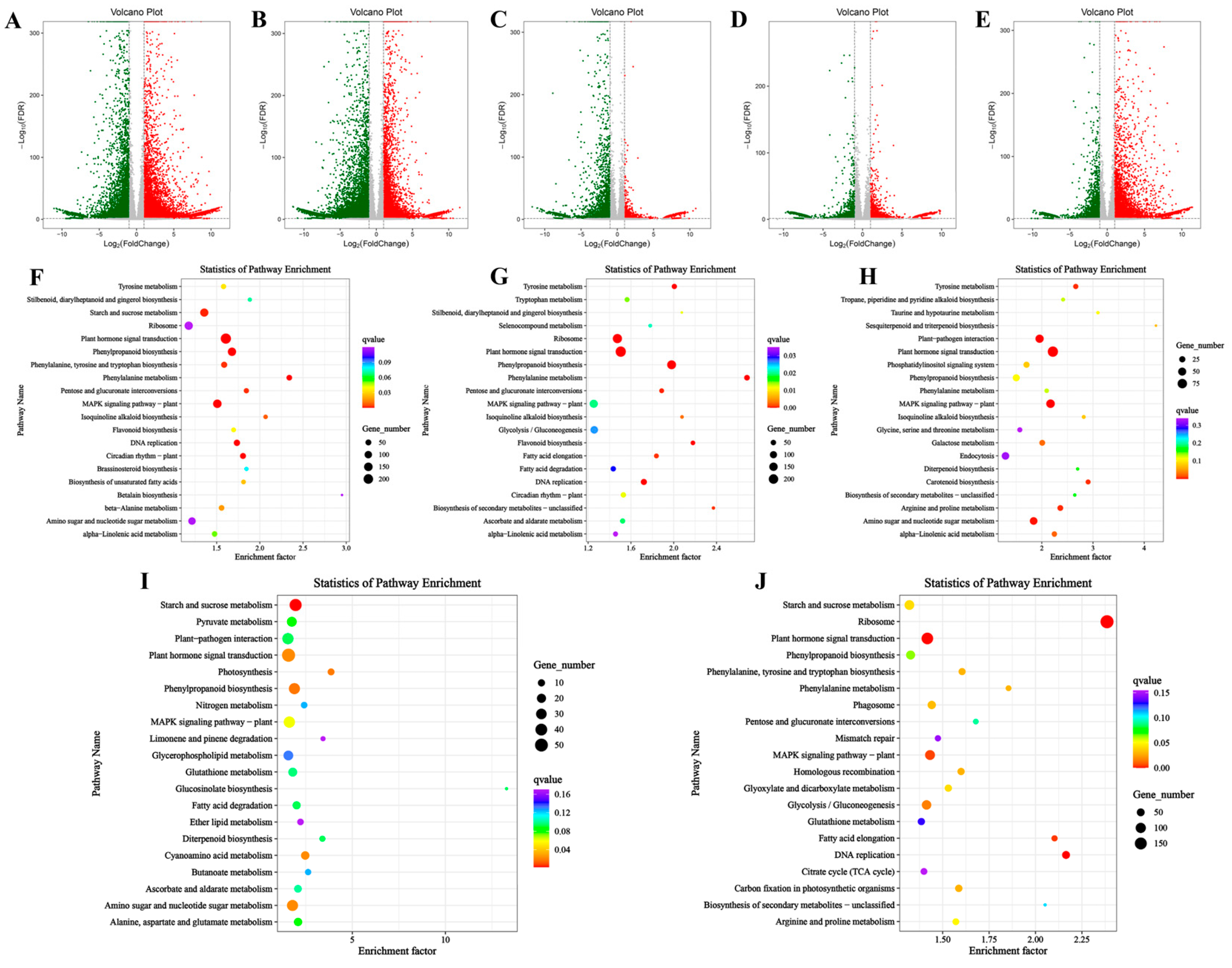
3.3. GO and KEGG Analysis of Differentially Expressed Genes in Bamboo Shoot Tips at Different Developmental Stages
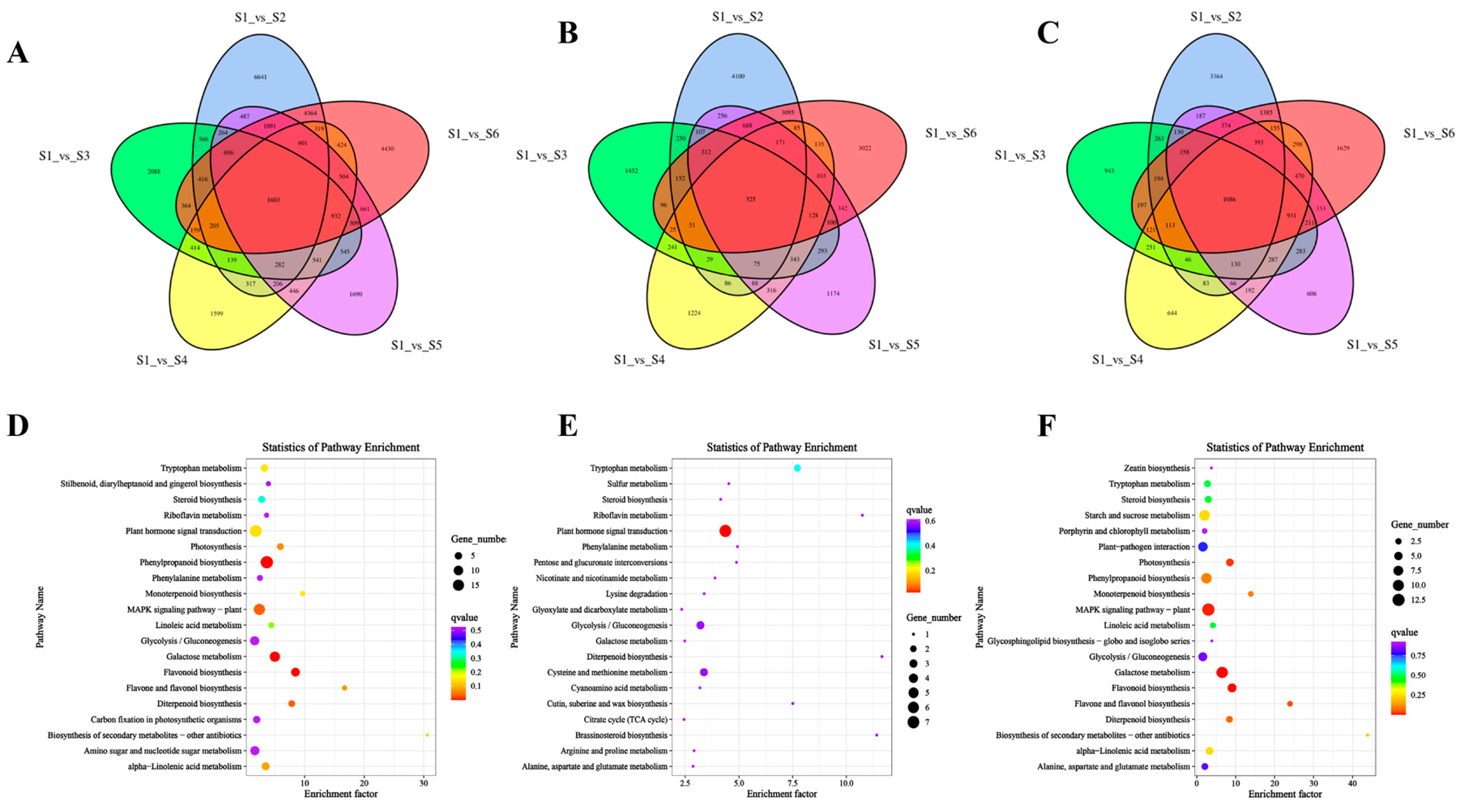
3.4. GO and KEGG Enrichment Analysis of Common DEGs at Different Developmental Stages
3.5. Co-Expression Clustering Analysis of Genes at Different Development Stages
3.6. Co-Expression Network Analysis Based on WGCNA
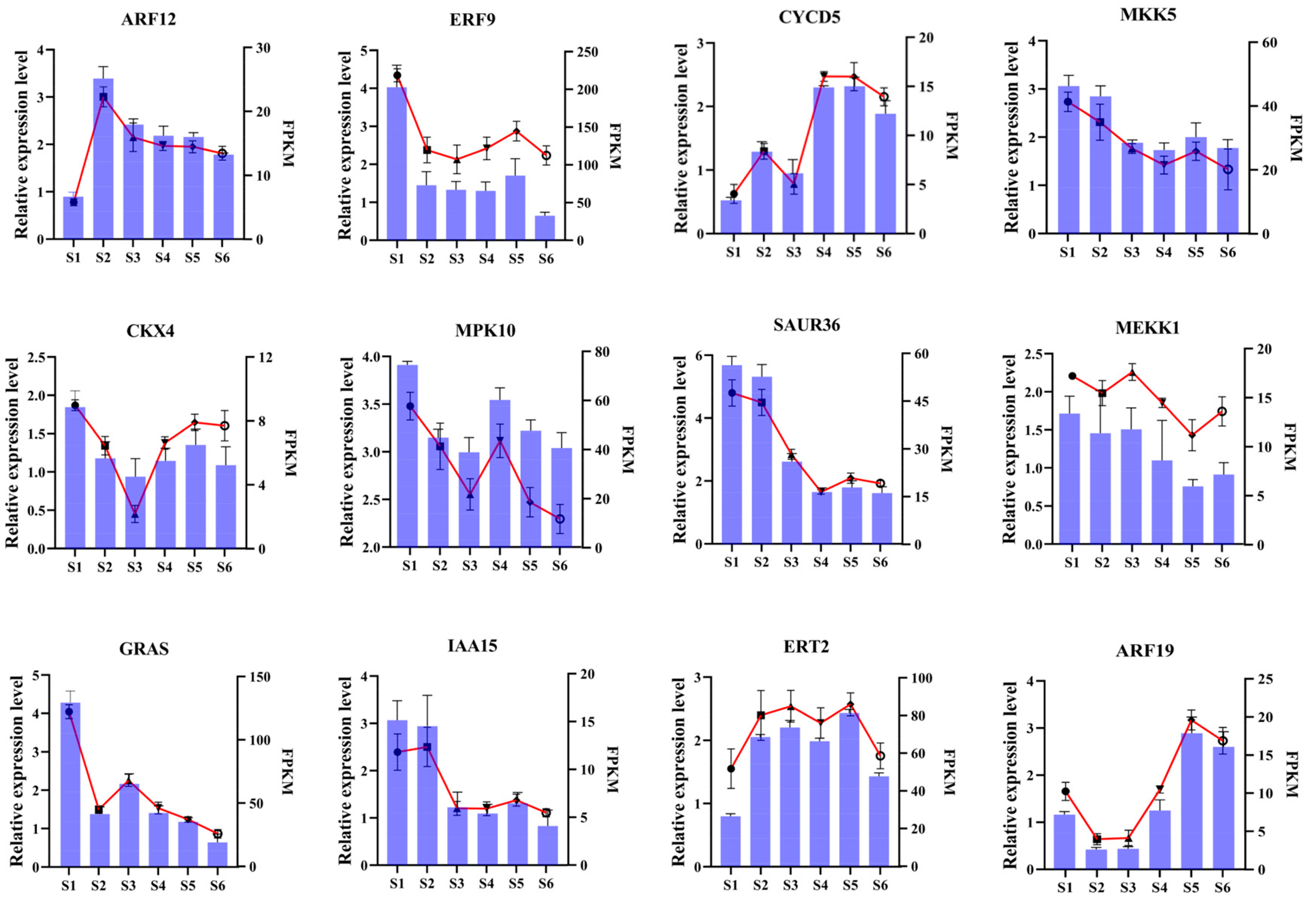
3.7. RT-qPCR Validation of DEGs
4. Discussion
4.1. Identification of Hormone Signaling Related DEGs During Bamboo Shoot Tips Development
4.2. MAPK Signaling Pathway Regulates Bamboo Shoot Tips Development
4.3. Glycolysis/Gluconeogenesis Pathway Is Involved in Bamboo Shoot Development
4.4. Association Between Key Signal Pathways in Early Development of Bamboo Shoots and Edible Traits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Soreng, R.J.; Peterson, P.M.; Zuloaga, F.O.; Romaschenko, K.; Clark, L.G.; Teisher, J.K.; Gillespie, L.J.; Barberá, P.; Welker, C.A.D.; Kellogg, E.A.; et al. A worldwide phylogenetic classification of the Poaceae (Gramineae) III: An update. J. Syst. Evol. 2022, 60, 476–521. [Google Scholar] [CrossRef]
- Chen, M.; Guo, L.; Ramakrishnan, M.; Fei, Z.; Vinod, K.K.; Ding, Y.; Jiao, C.; Gao, Z.; Zha, R.; Wang, C.; et al. Rapid growth of Moso bamboo (Phyllostachys edulis): Cellular roadmaps, transcriptome dynamics, and environmental factors. Plant Cell 2022, 34, 3577–3610. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, E.; Tanaka, C.; Mori, N.; Kuwahara, Y.; Tsuda, M. Phenylpropanoid amides of serotonin accumulate in witches’ broom diseased bamboo. Phytochemistry 2003, 64, 965–969. [Google Scholar] [CrossRef]
- Bal, L.M.; Singhal, P.; Satya, S.; Naik, S.N.; Kar, A. Bamboo shoot preservation for enhancing its business potential and local economy: A review. Crit. Rev. Food Sci. Nutr. 2012, 52, 804–814. [Google Scholar] [CrossRef]
- Yang, K.; Li, L.; Lou, Y.; Zhu, C.; Li, X.; Gao, Z. A regulatory network driving shoot lignification in rapidly growing bamboo. Plant Physiol. 2021, 187, 900–916. [Google Scholar] [CrossRef]
- Cui, K.; He, C.Y.; Zhang, J.G.; Duan, A.G.; Zeng, Y.F. Temporal and spatial profiling of internode elongation-associated protein expression in rapidly growing culms of bamboo. J. Proteome Res. 2012, 11, 2492–2507. [Google Scholar] [CrossRef]
- Herde, M.; Koo, A.J.; Howe, G.A. Elicitation of jasmonate-mediated defense responses by mechanical wounding and insect herbivory. Methods Mol. Biol. 2013, 1011, 51–61. [Google Scholar] [CrossRef]
- Peng, Z.; Zhang, C.; Zhang, Y.; Hu, T.; Mu, S.; Li, X.; Gao, J. Transcriptome sequencing and analysis of the fast growing shoots of moso bamboo (Phyllostachys edulis). PLoS ONE 2013, 8, e78944. [Google Scholar] [CrossRef]
- Peng, Z.; Lu, T.; Li, L.; Liu, X.; Gao, Z.; Hu, T.; Yang, X.; Feng, Q.; Guan, J.; Weng, Q.; et al. Genome-wide characterization of the biggest grass, bamboo, based on 10,608 putative full-length cDNA sequences. Bmc Plant Biol. 2010, 10, 116. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, X.; Ding, Y.; Fei, Z.; Jiao, C.; Fan, M.; Yao, B.; Xin, P.; Chu, J.; Wei, Q. Cellular and molecular characterization of a thick-walled variant reveal a pivotal role of shoot apical meristem in transverse development of bamboo culm. J. Exp. Bot. 2019, 70, 3911–3926. [Google Scholar] [CrossRef]
- Li, L.; Hu, T.; Li, X.; Mu, S.; Cheng, Z.; Ge, W.; Gao, J. Genome-wide analysis of shoot growth-associated alternative splicing in moso bamboo. Mol. Genet. Genomics 2016, 291, 1695–1714. [Google Scholar] [CrossRef]
- Wei, Q.; Jiao, C.; Ding, Y.; Gao, S.; Guo, L.; Chen, M.; Hu, P.; Xia, S.; Ren, G.; Fei, Z. Cellular and molecular characterizations of a slow-growth variant provide insights into the fast growth of bamboo. Tree Physiol. 2018, 38, 641–654. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Yu, A.; Hu, W.; Zhang, Z.; Wang, Z.; Meng, Y.; Yang, B.; Kuang, H. Extraction, purification, structural characteristic, health benefit, and product application of the polysaccharides from bamboo shoot: A review. Int. J. Biol. Macromol. 2024, 271, 132581. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.; Wang, H.; Liao, S.; Tang, Q.; Li, L.; Cui, Y.; He, Y. Transcriptome Sequencing and Analysis for Culm Elongation of the World’s Largest Bamboo (Dendrocalamus sinicus). PLoS ONE 2016, 11, e0157362. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Lian, C.; Liu, R.; Zhang, S.; Yang, F.; Fei, B. Comparison of metaxylem vessels and pits in four sympodial bamboo species. Sci. Rep. 2019, 9, 10876. [Google Scholar] [CrossRef]
- Li, L.; Li, J.; Wang, X.; Wang, W.; Leung, F.; Liu, X.; Wang, C. Growth reduction and alteration of nonstructural carbohydrate (NSC) allocation in a sympodial bamboo (Indocalamus decorus) under atmospheric O3 enrichment. Sci. Total Environ. 2022, 826, 154096. [Google Scholar] [CrossRef]
- Teng, J.; Xiang, T.; Huang, Z.; Wu, J.; Jiang, P.; Meng, C.; Li, Y.; Fuhrmann, J.J. Spatial distribution and variability of carbon storage in different sympodial bamboo species in China. J. Environ. Manage 2016, 168, 46–52. [Google Scholar] [CrossRef]
- Kaushal, R.; Kumar, A.; Alam, N.M.; Singh, I.; Mandal, D.; Tomar, J.M.S.; Mehta, H.; Lepcha, S.T.S.; Long, T.T.; Durai, J. Assessment of eco-hydrological parameters for important sympodial bamboo species in Himalayan foothills. Environ. Monit. Assess. 2021, 193, 468. [Google Scholar] [CrossRef]
- Zheng, Y.; Yang, D.; Rong, J.; Chen, L.; Zhu, Q.; He, T.; Chen, L.; Ye, J.; Fan, L.; Gao, Y.; et al. Allele-aware chromosome-scale assembly of the allopolyploid genome of hexaploid Ma bamboo (Dendrocalamus latiflorus Munro). J. Integr. Plant Biol. 2022, 64, 649–670. [Google Scholar] [CrossRef]
- Kalderis, D.; Seifi, A.; Kieu Trang, T.; Tsubota, T.; Anastopoulos, I.; Manariotis, I.; Pashalidis, I.; Khataee, A. Bamboo-derived adsorbents for environmental remediation: A review of recent progress. Environ. Res. 2023, 224, 115533. [Google Scholar] [CrossRef]
- Gan, J.; Chen, M.; Semple, K.; Liu, X.; Dai, C.; Tu, Q. Life cycle assessment of bamboo products: Review and harmonization. Sci. Total Environ. 2022, 849, 157937. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Mo, W.; Lv, B.; Wang, W.; He, H.; Jian, C.; Liu, X.; Li, S.; Guo, Y. A Review of the Nutritional Composition, Storage Challenges, Processing Technology and Widespread Use of Bamboo Shoots. Foods 2024, 13, 3539. [Google Scholar] [CrossRef] [PubMed]
- Singhal, P.; Bal, L.M.; Satya, S.; Sudhakar, P.; Naik, S.N. Bamboo shoots: A novel source of nutrition and medicine. Crit. Rev. Food Sci. Nutr. 2013, 53, 517–534. [Google Scholar] [CrossRef]
- Behera, P.; Balaji, S. Health Benefits of Fermented Bamboo Shoots: The Twenty-First Century Green Gold of Northeast India. Appl. Biochem. Biotechnol. 2021, 193, 1800–1812. [Google Scholar] [CrossRef]
- Hu, S.K.; Zhang, M.; Yang, Y.Q.; Xuan, W.; Zou, Z.W.; Arkorful, E.; Chen, Y.; Ma, Q.P.; Jeyaraj, A.; Chen, X.; et al. A novel insight into nitrogen and auxin signaling in lateral root formation in tea plant [Camellia sinensis (L.) O. Kuntze]. Bmc Plant Biol. 2020, 20, 232. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinformatics 2011, 12, 323. [Google Scholar] [CrossRef]
- Liu, S.; Wang, Z.; Zhu, R.; Wang, F.; Cheng, Y.; Liu, Y. Three Differential Expression Analysis Methods for RNA Sequencing: Limma, EdgeR, DESeq2. J. Vis. Exp. 2021, 175, e62528. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Lan, Y.; Wu, L.; Wu, M.; Liu, H.; Gao, Y.; Zhang, K.; Xiang, Y. Transcriptome analysis reveals key genes regulating signaling and metabolic pathways during the growth of moso bamboo (Phyllostachys edulis) shoots. Physiol. Plant 2021, 172, 91–105. [Google Scholar] [CrossRef]
- Shen, Z.; Zhang, Y.H.; Zhang, L.; Li, Y.; Sun, Y.D.; Li, Z.Y. Changes in the distribution of endogenous hormones in Phyllostachys edulis ‘Pachyloen’ during bamboo shooting. PLoS ONE 2020, 15, e0241806. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Jiao, C.; Guo, L.; Ding, Y.; Cao, J.; Feng, J.; Dong, X.; Mao, L.; Sun, H.; Yu, F.; et al. Exploring key cellular processes and candidate genes regulating the primary thickening growth of Moso underground shoots. New Phytol. 2017, 214, 81–96. [Google Scholar] [CrossRef] [PubMed]
- Cai, M.; Cheng, W.; Bai, Y.; Mu, C.; Zheng, H.; Cheng, Z.; Gao, J. PheGRF4e initiated auxin signaling during moso bamboo shoot development. Mol. Biol. Rep. 2022, 49, 8815–8825. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Cheng, Z.; Ma, Y.; Bai, Q.; Li, X.; Cao, Z.; Wu, Z.; Gao, J. The association of hormone signalling genes, transcription and changes in shoot anatomy during moso bamboo growth. Plant Biotechnol. J. 2018, 16, 72–85. [Google Scholar] [CrossRef]
- Guo, L.; Sun, X.; Li, Z.; Wang, Y.; Fei, Z.; Jiao, C.; Feng, J.; Cui, D.; Feng, X.; Ding, Y.; et al. Morphological dissection and cellular and transcriptome characterizations of bamboo pith cavity formation reveal a pivotal role of genes related to programmed cell death. Plant Biotechnol. J. 2019, 17, 982–997. [Google Scholar] [CrossRef]
- Zhou, B.; Zhang, X.; Zhang, H.; Yao, W.; Li, L.; Lin, S.; Cao, F. Effects of retaining different number of mother bamboo on the growth and NSCs allocation of new-born seedlings in Phyllostachys edulis at different age. Bmc Plant Biol. 2025, 25, 60. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, D.; Zhang, S.; Lou, Y.; An, X.; Jiang, Z.; Gao, Z. Transcriptome and miRNAome analysis reveals components regulating tissue differentiation of bamboo shoots. Plant Physiol. 2022, 188, 2182–2198. [Google Scholar] [CrossRef]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Ding, L.N.; Li, Y.T.; Wu, Y.Z.; Li, T.; Geng, R.; Cao, J.; Zhang, W.; Tan, X.L. Plant Disease Resistance-Related Signaling Pathways: Recent Progress and Future Prospects. Int. J. Mol. Sci. 2022, 23, 16200. [Google Scholar] [CrossRef]
- Zhang, Y.; Berman, A.; Shani, E. Plant Hormone Transport and Localization: Signaling Molecules on the Move. Annu. Rev. Plant Biol. 2023, 74, 453–479. [Google Scholar] [CrossRef]
- Mathura, S.R.; Sutton, F.; Bowrin, V. Genome-wide identification, characterization, and expression analysis of the sweet potato (Ipomoea batatas [L.] Lam.) ARF, Aux/IAA, GH3, and SAUR gene families. Bmc Plant Biol. 2023, 23, 622. [Google Scholar] [CrossRef]
- Stortenbeker, N.; Bemer, M. The SAUR gene family: The plant’s toolbox for adaptation of growth and development. J. Exp. Bot. 2019, 70, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Ori, N. Dissecting the Biological Functions of ARF and Aux/IAA Genes. Plant Cell 2019, 31, 1210–1211. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Feng, S.; Pang, J.; Mao, L.; Shou, H.; Xie, J. Effect of heat treatment on lignification of postharvest bamboo shoots (Phyllostachys praecox f. prevernalis). Food Chem. 2012, 135, 2182–2187. [Google Scholar] [CrossRef]
- Xie, X.; Hu, S.; Liu, L.; Pan, H.; Huang, H.; Cao, X.; Qiao, G.; Han, X.; Qiu, W.; Lu, Z.; et al. Genome-Wide Analysis of HECT E3 Ligases Members in Phyllostachys edulis Provides Insights into the Role of PeHECT1 in Plant Abiotic Stress Response. Int. J. Mol. Sci. 2024, 25, 11896. [Google Scholar] [CrossRef] [PubMed]
- Su, S.; Xuan, X.; Tan, J.; Yu, Z.; Jiao, Y.; Zhang, Z.; Ramakrishnan, M. Analysis of the CHS Gene Family Reveals Its Functional Responses to Hormones, Salinity, and Drought Stress in Moso Bamboo (Phyllostachys edulis). Plants 2025, 14, 161. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, S. Mitogen-activated protein kinase cascades in plant signaling. J. Integr. Plant Biol. 2022, 64, 301–341. [Google Scholar] [CrossRef]
- Xu, J.; Zhang, S. Mitogen-activated protein kinase cascades in signaling plant growth and development. Trends Plant Sci. 2015, 20, 56–64. [Google Scholar] [CrossRef]
- Shao, Y.; Yu, X.; Xu, X.; Li, Y.; Yuan, W.; Xu, Y.; Mao, C.; Zhang, S.; Xu, J. The YDA-MKK4/MKK5-MPK3/MPK6 Cascade Functions Downstream of the RGF1-RGI Ligand-Receptor Pair in Regulating Mitotic Activity in Root Apical Meristem. Mol. Plant 2020, 13, 1608–1623. [Google Scholar] [CrossRef]
- Wu, M.; Wang, S.; Ma, P.; Li, B.; Hu, H.; Wang, Z.; Qiu, Q.; Qiao, Y.; Niu, D.; Lukowitz, W.; et al. Dual roles of the MPK3 and MPK6 mitogen-activated protein kinases in regulating Arabidopsis stomatal development. Plant Cell 2024, 36, 4576–4593. [Google Scholar] [CrossRef]
- Liu, Y.; Leary, E.; Saffaf, O.; Frank Baker, R.; Zhang, S. Overlapping functions of YDA and MAPKKK3/MAPKKK5 upstream of MPK3/MPK6 in plant immunity and growth/development. J. Integr. Plant Biol. 2022, 64, 1531–1542. [Google Scholar] [CrossRef]
- Chen, L.; Dou, P.; Li, L.; Chen, Y.; Yang, H. Transcriptome-wide analysis reveals core transcriptional regulators associated with culm development and variation in Dendrocalamus sinicus, the strongest woody bamboo in the world. Heliyon 2022, 8, e12600. [Google Scholar] [CrossRef]
- Fernie, A.R.; Carrari, F.; Sweetlove, L.J. Respiratory metabolism: Glycolysis, the TCA cycle and mitochondrial electron transport. Curr. Opin. Plant Biol. 2004, 7, 254–261. [Google Scholar] [CrossRef]
- O’Leary, B.; Plaxton, W.C. Multifaceted functions of post-translational enzyme modifications in the control of plant glycolysis. Curr. Opin. Plant Biol. 2020, 55, 28–37. [Google Scholar] [CrossRef]
- Kleczkowski, L.A.; Igamberdiev, A.U. Multiple Roles of Glycerate Kinase-From Photorespiration to Gluconeogenesis, C4 Metabolism, and Plant Immunity. Int. J. Mol. Sci. 2024, 25, 3258. [Google Scholar] [CrossRef]
- Walker, R.P.; Benincasa, P.; Battistelli, A.; Moscatello, S.; Tecsi, L.; Leegood, R.C.; Famiani, F. Gluconeogenesis and nitrogen metabolism in maize. Plant Physiol. Biochem. 2018, 130, 324–333. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, F. Glycolysis and gluconeogenesis: A teaching view. J. Biol. Chem. 2021, 296, 100016. [Google Scholar] [CrossRef] [PubMed]
- Ndlovu, N.; Spillane, C.; McKeown, P.C.; Cairns, J.E.; Das, B.; Gowda, M. Genome-wide association studies of grain yield and quality traits under optimum and low-nitrogen stress in tropical maize (Zea mays L.). Theor. Appl. Genet. 2022, 135, 4351–4370. [Google Scholar] [CrossRef]
- Qian, X.; Zhao, X.; Yu, L.; Yin, Y.; Zhang, X.D.; Wang, L.; Li, J.X.; Zhu, Q.; Luo, J.L. Current status of GABA receptor subtypes in analgesia. Biomed. Pharmacother. 2023, 168, 115800. [Google Scholar] [CrossRef]
- Coelho, A.I.; Berry, G.T.; Rubio-Gozalbo, M.E. Galactose metabolism and health. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 422–427. [Google Scholar] [CrossRef]
- Wang, S.; Ito, T.; Uehara, M.; Naito, S.; Takano, J. UDP-D-galactose synthesis by UDP-glucose 4-epimerase 4 is required for organization of the trans-Golgi network/early endosome in Arabidopsis thaliana root epidermal cells. J. Plant Res. 2015, 128, 863–873. [Google Scholar] [CrossRef]
- Liu, B.; Tang, L.; Chen, Q.; Zhu, L.; Zou, X.; Li, B.; Zhou, Q.; Fu, Y.; Lu, Y. Lignin Distribution on Cell Wall Micro-Morphological Regions of Fibre in Developmental Phyllostachys pubescens Culms. Polymers 2022, 14, 312. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Kitamura, S.; Sone, Y.; Itoh, T. Immunohistochemical localization of hemicelluloses and pectins varies during tissue development in the bamboo culm. Histochem. J. 2002, 34, 535–544. [Google Scholar] [CrossRef] [PubMed]
- Scaman, C.H.; Jim, V.J.; Hartnett, C. Free galactose concentrations in fresh and stored apples (Malus domestica) and processed apple products. J. Agric. Food Chem. 2004, 52, 511–517. [Google Scholar] [CrossRef]
- Sun, H.; Li, L.; Lou, Y.; Zhao, H.; Yang, Y.; Gao, Z. Cloning and Preliminary Functional Analysis of PeUGE Gene from Moso Bamboo (Phyllostachys edulis). DNA Cell Biol. 2016, 35, 706–714. [Google Scholar] [CrossRef] [PubMed]
- Hoftberger, M.; Althammer, M.; Foissner, I.; Tenhaken, R. Galactose induces formation of cell wall stubs and cell death in Arabidopsis roots. Planta 2022, 256, 26. [Google Scholar] [CrossRef]
- Zhang, X.; Yoshihara, K.; Miyata, N.; Hata, T.; Altaisaikhan, A.; Takakura, S.; Asano, Y.; Izuno, S.; Sudo, N. Dietary tryptophan, tyrosine, and phenylalanine depletion induce reduced food intake and behavioral alterations in mice. Physiol. Behav. 2022, 244, 113653. [Google Scholar] [CrossRef]
- Gomes, G.L.B.; Scortecci, K.C. Auxin and its role in plant development: Structure, signalling, regulation and response mechanisms. Plant Biol. 2021, 23, 894–904. [Google Scholar] [CrossRef]
- Ren, X.; Zou, Q.; Yuan, C.; Chang, R.; Xing, R.; Yan, X. The Dominant Role of Oxygen in Modulating the Chemical Evolution Pathways of Tyrosine in Peptides: Dityrosine or Melanin. Angew. Chem. Int. Ed. Engl. 2019, 58, 5872–5876. [Google Scholar] [CrossRef]
- Ishikawa, S.; Taira, T.; Niki, T.; Takahashi-Niki, K.; Maita, C.; Maita, H.; Ariga, H.; Iguchi-Ariga, S.M. Oxidative status of DJ-1-dependent activation of dopamine synthesis through interaction of tyrosine hydroxylase and 4-dihydroxy-L-phenylalanine (L-DOPA) decarboxylase with DJ-1. J. Biol. Chem. 2009, 284, 28832–28844. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, S.; Dong, M.; Guo, Q. The Transcriptome Analysis Provides New Insights into Signaling for Bamboo Shoot Development of Sympodial Bamboo. Foods 2025, 14, 1647. https://doi.org/10.3390/foods14091647
Hu S, Dong M, Guo Q. The Transcriptome Analysis Provides New Insights into Signaling for Bamboo Shoot Development of Sympodial Bamboo. Foods. 2025; 14(9):1647. https://doi.org/10.3390/foods14091647
Chicago/Turabian StyleHu, Shunkai, Mengran Dong, and Qirong Guo. 2025. "The Transcriptome Analysis Provides New Insights into Signaling for Bamboo Shoot Development of Sympodial Bamboo" Foods 14, no. 9: 1647. https://doi.org/10.3390/foods14091647
APA StyleHu, S., Dong, M., & Guo, Q. (2025). The Transcriptome Analysis Provides New Insights into Signaling for Bamboo Shoot Development of Sympodial Bamboo. Foods, 14(9), 1647. https://doi.org/10.3390/foods14091647