

Fu Brick Tea Protects the Intestinal Barrier and Ameliorates Colitis in Mice by Regulating Gut Microbiota

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Test Materials and Equipment
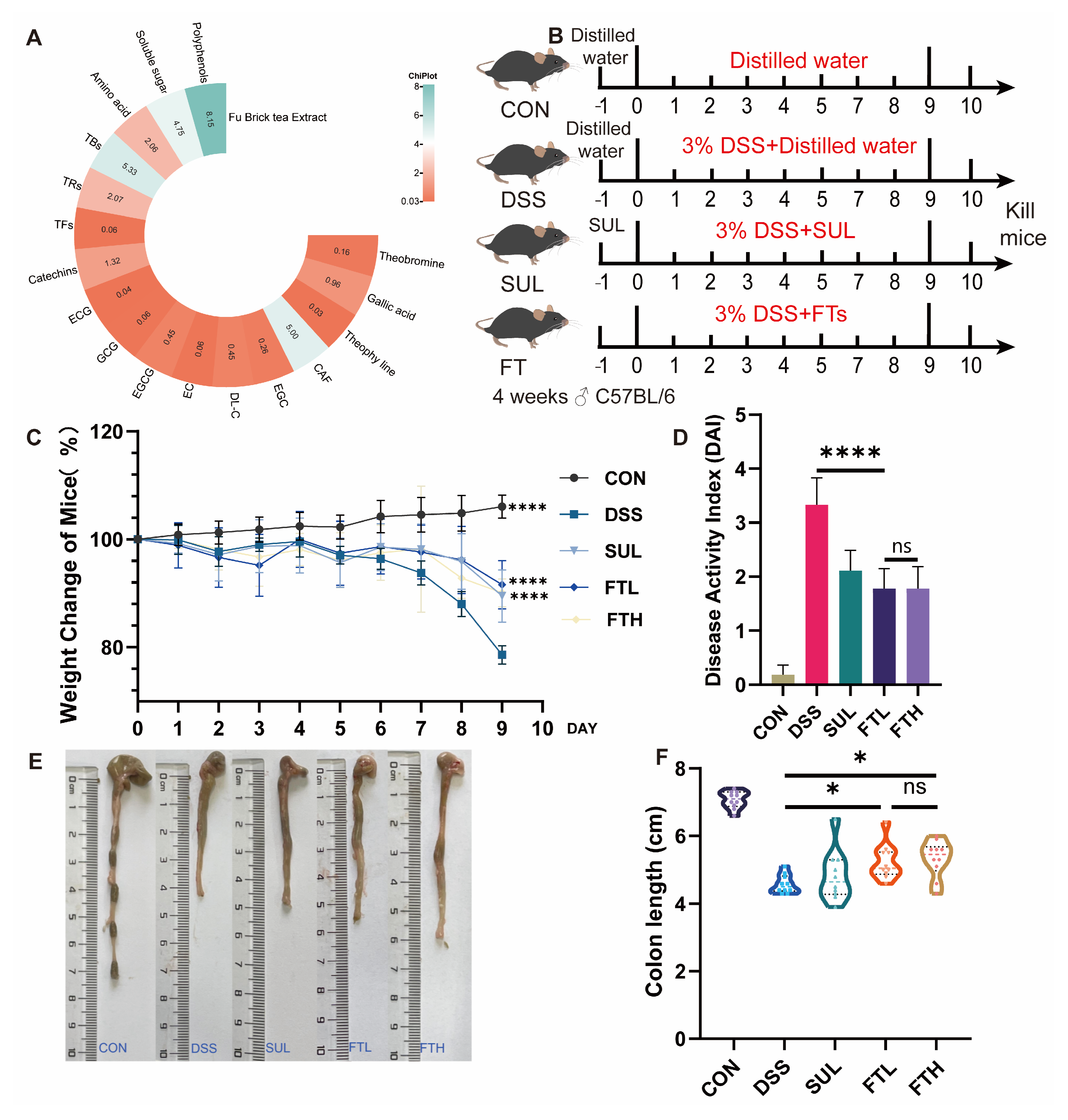
2.2. Animal Experiment Design
2.3. Disease Activity Index Evaluation
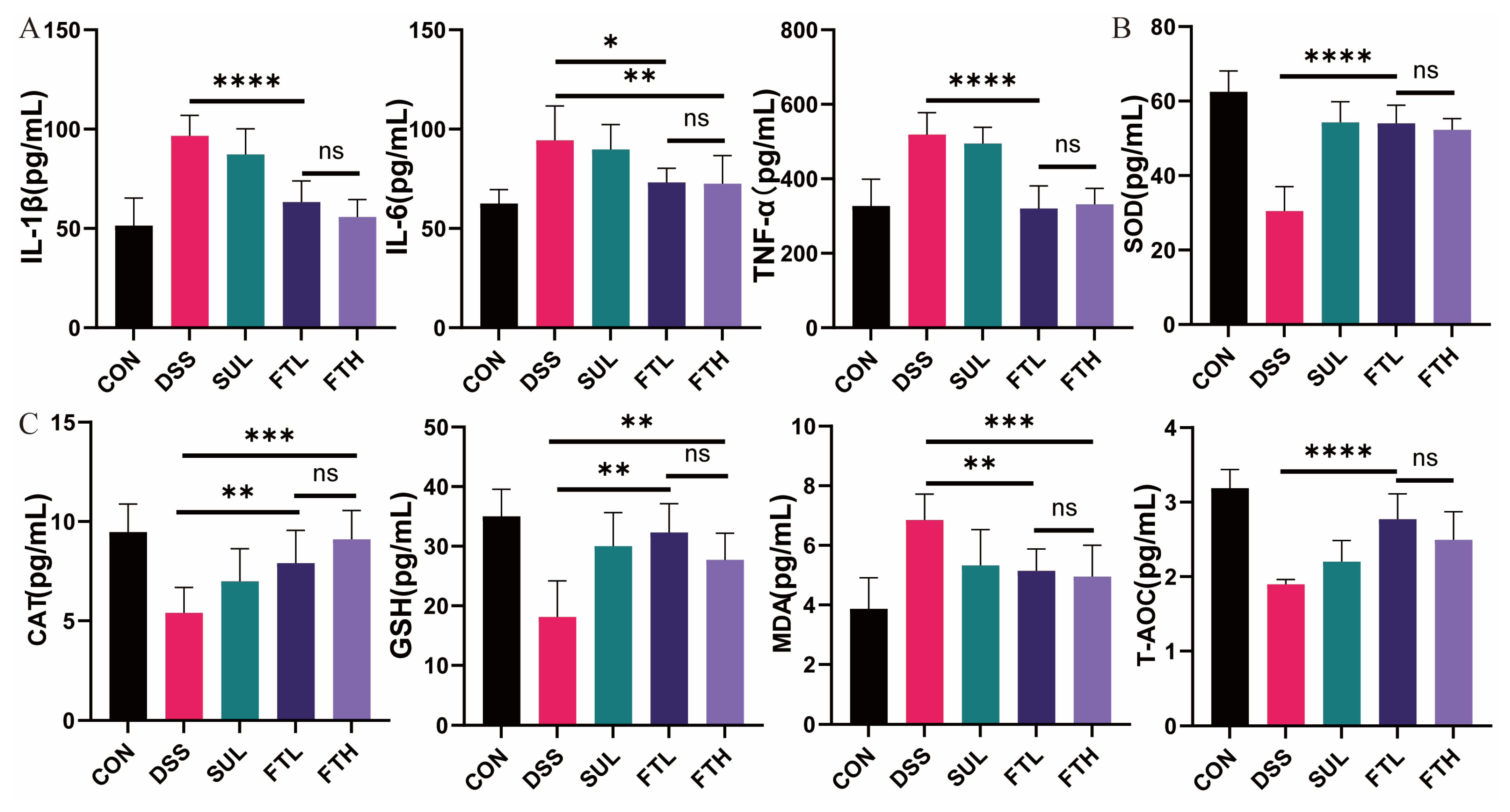
2.4. Detection of Inflammatory Factors in Serum
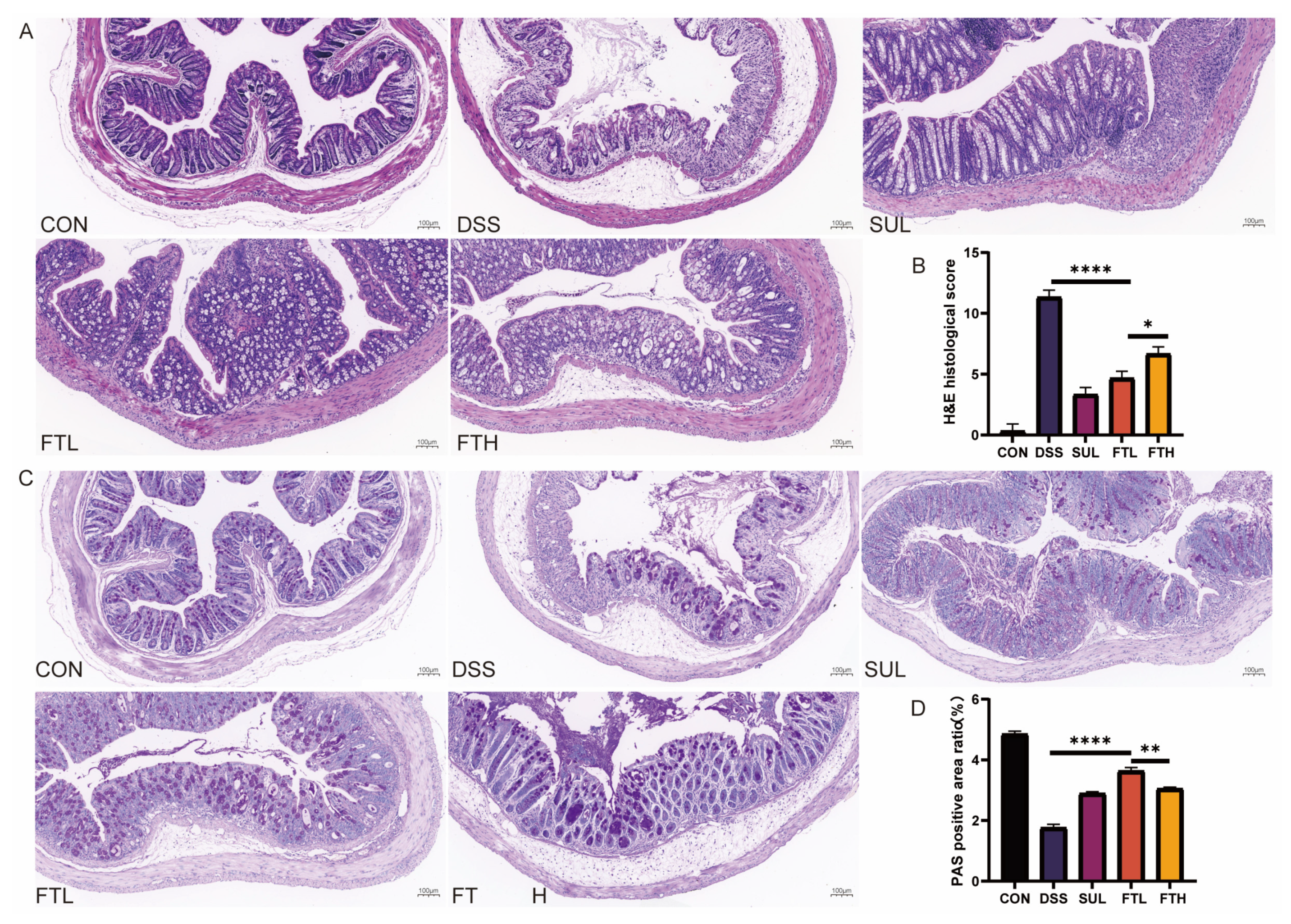
2.5. Pathological Observation of Mice Colon Tissue Sections
2.6. Immunofluorescence Detection of ZO-1 Expression in Colon Tissue
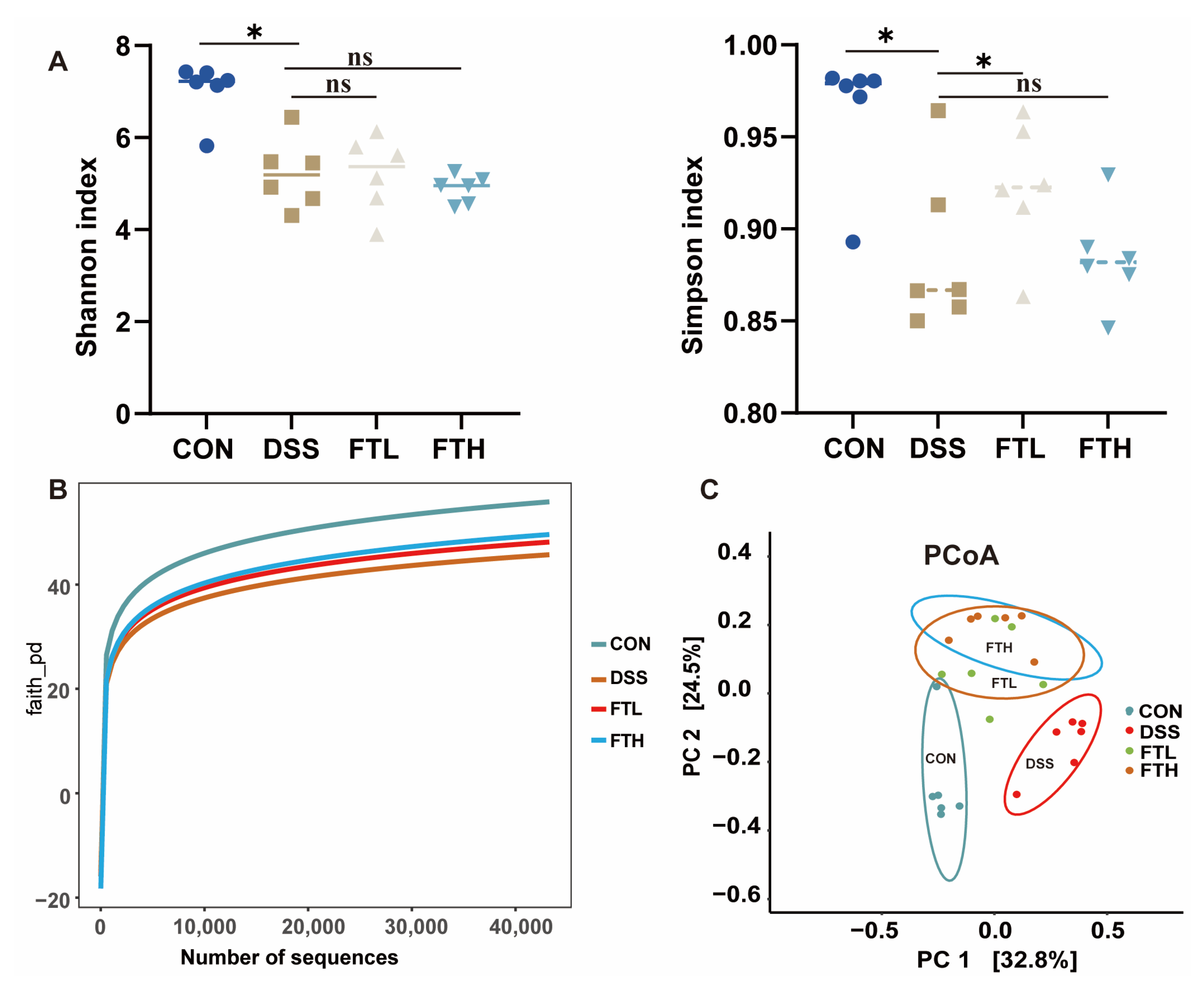
2.7. Gut Microbiota 16sRNA Detection and Analysis
2.8. Detection and Analysis of SCFAs in Intestinal Contents
2.9. Data Statistics
3. Results
3.1. Analysis of Functional Components of FBT
3.2. Effect of FBT on Disease Activity Index of Colitis Mice
3.3. Effects of FBT on Inflammatory/Oxidative Factors of Colitis Mice
3.4. Effects of FBT on Colon Morphology and Structure in Colitis Mice
3.5. Effect of FBT on Intestinal Barrier in Colitis Mice
3.6. Effects of FBT on the Diversity of Gut Microbiota in Mice with Colitis
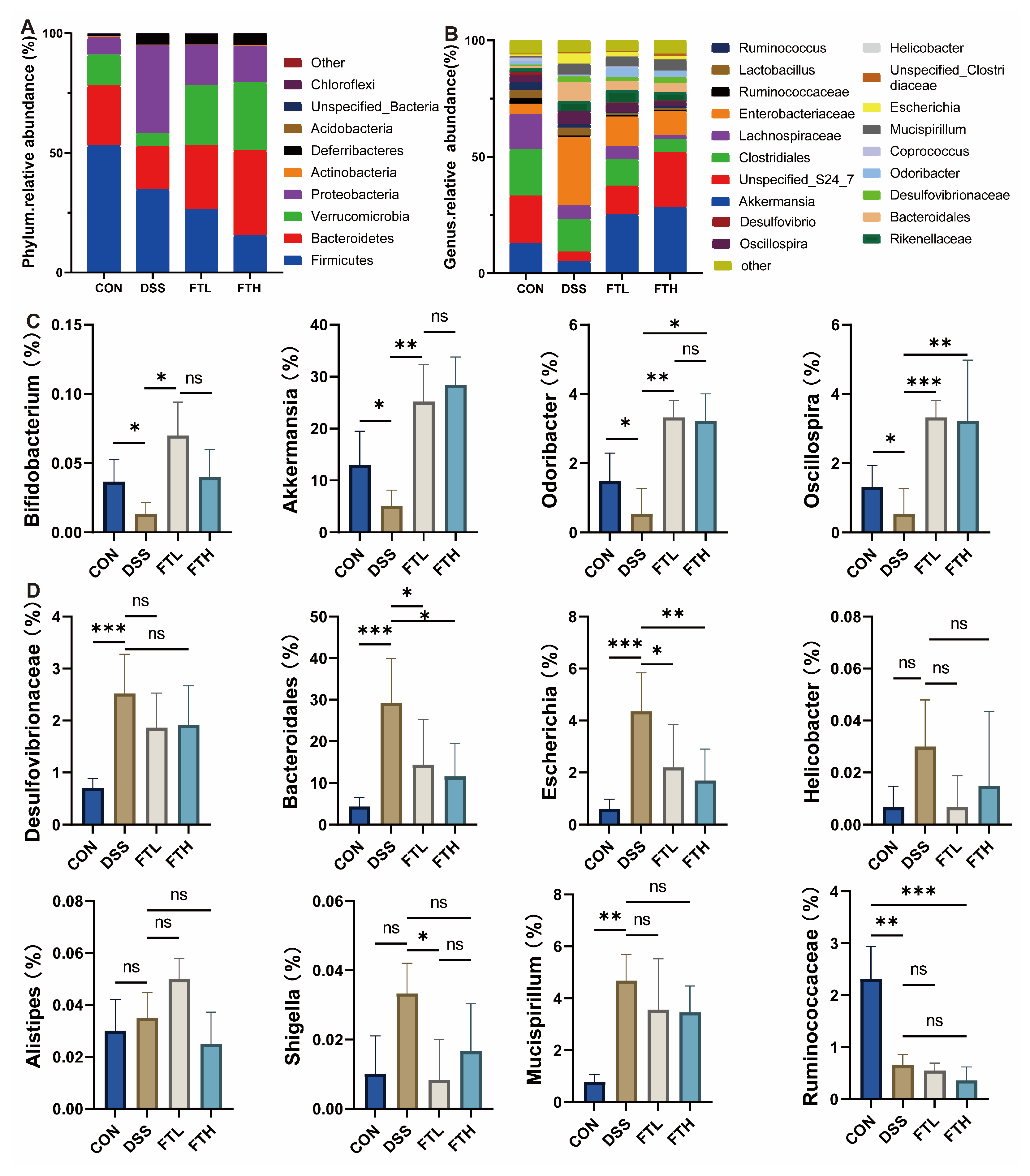
3.7. Effects of FBT on Gut Microbiota in Colitis Mice
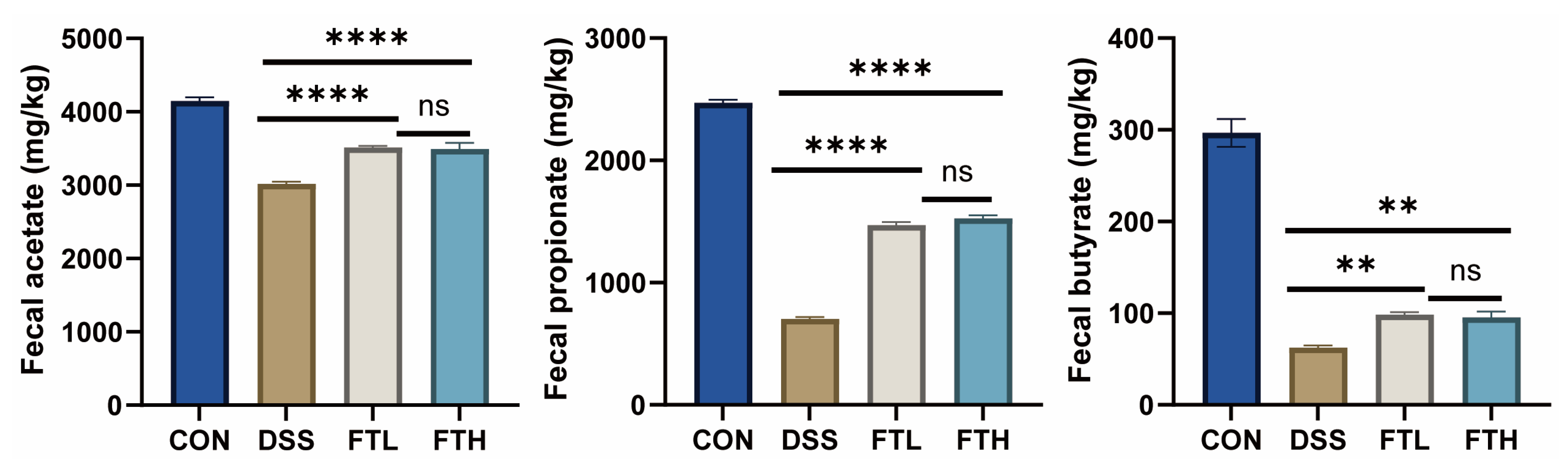
3.8. Analysis of Key Flora Abundance and SCFA Content
3.9. Correlation Analysis Between Gut Microbiota Species and Key Indicators of Colitis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FBT | Fu Brick tea |
| ZO-1 | Intestinal barrier protein |
| IBD | Inflammatory bowel disease |
| UC | Ulcerative colitis |
| CD | Crohn’s disease |
| ROS | Reactive oxygen species |
| IL-1β | Interleukin-1 beta |
| IL-6 | Interleukin-6 |
| TNF-α | Tumor necrosis factor-α |
| SOD | Superoxide dismutase |
| CAT | Catalase |
| GSH | Glutathione |
| MDA | Malondialdehyde |
| T-AOC | Total Antioxidant Capacity |
| DSS | Dextran sodium sulfate |
| CON | Normal control group |
| DSS | DSS enteritis model group |
| SUL | Positive control group (treated with sulfasalazine) |
| FTL | 200 mg/kg FBT group |
| FTH | 400 mg/kg FBT group |
| LDA | Linear discriminant analysis |
| F/B | The ratio of Firmicutes/Bacteroidetes |
| SCFAs | Short-chain fatty acids |
| HPLC | High-performance liquid chromatography |
| H&E | Hematoxylin and eosin staining |
| PAS | Periodic acid-Schiff staining |
| FID | Flame ionization detector |
| ANOVA | One-way analysis of variance |
References
- Kotsiliti, E. Phage therapy suppresses gut inflammation in IBD. Nat. Biotechnol. 2022, 9, 1327. [Google Scholar] [CrossRef]
- Li, D.; Feng, Y.; Tian, M.; Ji, J.; Hu, X.; Chen, F. Gut microbiota-derived inosine from dietary barley leaf supplementation attenuates colitis through PPARγ signaling activation. Microbiome 2021, 1, 83. [Google Scholar] [CrossRef]
- Fitzpatrick, J.; Melton, S.; Yao, C.; Gibson, P.; Halmos, E. Dietary management of adults with IBD—The emerging role of dietary therapy. Nature reviews. Gastroenterol. Hepatol. 2022, 19, 652–669. [Google Scholar] [CrossRef]
- Bai, Y.; Zeng, Z.; Xie, Z.; Chen, G.; Chen, D.; Sun, Y.; Zeng, X.; Liu, Z. Effects of polysaccharides from Fuzhuan brick tea on immune function and gut microbiota of cyclophosphamide-treated mice. J. Nutr. Biochem. 2022, 101, 108947. [Google Scholar] [CrossRef]
- Iliev, I. Mycobiota-host immune interactions in IBD: Coming out of the shadows. Nat. Rev. Gastroenterol. Hepatol. 2022, 2, 91–92. [Google Scholar] [CrossRef]
- Adolph, T.; Zhang, J. Diet fuelling inflammatory bowel diseases: Preclinical and clinical concepts. Gut 2022, 71, 2574–2586. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Pei, G.; Sun, X.; Xiao, Y.; Miao, C.; Zhou, L.; Wang, B.; Yang, L.; Yu, M.; Zhang, Z.; et al. RhoB affects colitis through modulating cell signaling and intestinal microbiome. Microbiome 2022, 1, 149. [Google Scholar] [CrossRef]
- Wu, Z.; Huang, S.; Li, T.; Li, N.; Han, D.; Zhang, B.; Xu, Z.; Zhang, S.; Pang, J.; Wang, S.; et al. Gut microbiota from green tea polyphenol-dosed mice improves intestinal epithelial homeostasis and ameliorates experimental colitis. Microbiome 2021, 1, 184. [Google Scholar] [CrossRef]
- Scarallo, L.; Griffiths, A. Medical therapy of paediatric inflammatory bowel disease. Nature reviews. Gastroenterol. Hepatol. 2022, 19, 685–686. [Google Scholar] [CrossRef]
- Manichanh, C.; Borruel, N.; Casellas, F.; Guarner, F. The gut microbiota in IBD. Nat. Rev. Gastroenterol. Hepatol. 2012, 10, 599–608. [Google Scholar] [CrossRef]
- Olson, C.A.; Vuong, H.E.; Yano, J.M.; Liang, Q.Y.; Nusbaum, D.J.; Hsiao, E.Y. The Gut Microbiota Mediates the Anti-Seizure Effects of the Ketogenic Diet. Cell 2018, 7, 1728–1741.e13. [Google Scholar] [CrossRef]
- Jubair, W.K.; Hendrickson, J.D.; Severs, E.L.; Schulz, H.M.; Adhikari, S.; Ir, D.; Pagan, J.D.; Anthony, R.M.; Robertson, C.E.; Frank, D.N.; et al. Modulation of Inflammatory Arthritis in Mice by Gut Microbiota Through Mucosal Inflammation and Autoantibody Generation. Arthritis Rheumatol. 2018, 8, 1220–1233. [Google Scholar] [CrossRef] [PubMed]
- Thaiss, C.A.; Zmora, N.; Levy, M.; Elinav, E. The microbiome and innate immunity. Nature 2016, 7610, 65–74. [Google Scholar] [CrossRef]
- Sinha, A.; Li, Y.; Mirzaei, M.K.; Shamash, M.; Samadfam, R.; King, I.L.; Maurice, C.F. Transplantation of bacteriophages from ulcerative colitis patients shifts the gut bacteriome and exacerbates the severity of DSS colitis. Microbiome 2022, 10, 105. [Google Scholar] [CrossRef]
- Shao, X.; Sun, S.; Zhou, Y.; Wang, H.; Yu, Y.; Hu, T.; Yao, Y.; Zhou, C. Bacteroides fragilis restricts colitis-associated cancer via negative regulation of the NLRP3 axis. Cancer Lett. 2021, 523, 170–181. [Google Scholar] [CrossRef]
- Gevers, D.; Kugathasan, S.; Denson, L.A.; Vázquez-Baeza, Y.; Van Treuren, W.; Ren, B.; Schwager, E.; Knights, D.; Song, S.J.; Yassour, M.; et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host Microbe 2014, 3, 382–392. [Google Scholar] [CrossRef]
- Lu, X.; Jing, Y.; Zhang, N.; Cao, Y. Eurotium cristatum, a Probiotic Fungus from Fuzhuan Brick Tea, and Its Polysaccharides Ameliorated DSS-Induced Ulcerative Colitis in Mice by Modulating the Gut Microbiota. J. Agric. Food Chem. 2022, 70, 2957–2967. [Google Scholar] [CrossRef]
- Yang, W.; Ren, D.; Zhao, Y.; Liu, L.; Yang, X. Fuzhuan Brick Tea Polysaccharide Improved Ulcerative Colitis in Association with Gut Microbiota-Derived Tryptophan Metabolism. J. Agric. Food Chem. 2021, 69, 8448–8459. [Google Scholar] [CrossRef]
- Liu, Y.; Luo, L.; Luo, Y.; Zhang, J.; Wang, X.; Sun, K.; Zeng, L. Prebiotic Properties of Green and Dark Tea Contribute to Protective Effects in Chemical-Induced Colitis in Mice: A Fecal Microbiota Transplantation Study. J. Agric. Food Chem. 2020, 23, 6368–6380. [Google Scholar] [CrossRef]
- Zhang, Y.; Lin, Y.; Xiong, Y.; Huang, J.; Liu, Z. An Analysis of the Intestinal Microbiome Combined with Metabolomics to Explore the Mechanism of How Jasmine Tea Improves Depression in CUMS-Treated Rats. Foods 2024, 13, 2636. [Google Scholar] [CrossRef]
- Chen, D.; Chen, G.; Chen, C.; Zeng, X.; Ye, H. Prebiotics effects in vitro of polysaccharides from tea flowers on gut microbiota of healthy persons and patients with inflammatory bowel disease. Int. J. Biol. Macromol. 2020, 158, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.; Chen, X.Y.; An, L.Y.; Li, C.C.; Zhao, N.; Yang, F.; You, S.T.; Hou, C.Z.; Li, K.; Jiang, C.; et al. Development of Novel Tetrahydroquinoline Inhibitors of NLRP3 Inflammasome for Potential Treatment of DSS-Induced Mice Colitis. J. Med. Chem. 2021, 1, 871–889. [Google Scholar] [CrossRef]
- Argmann, C.; Hou, R.; Ungaro, R.; Irizar, H.; Al-Taie, Z.; Huang, R.; Kosoy, R.; Venkat, S.; Song, W.; Di’Narzo, A.; et al. Biopsy and blood-based molecular biomarker of inflammation in IBD. Gut 2022, 72, 1271–1287. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.Y.; Wu, X.Y.; Li, M.Y.; Li, X.T.; Huang, R.M.; Sun, Y.M.; Xu, Z.L. Noni (Morinda citrifolia L.) Fruit Polysaccharides Regulated IBD Mice Via Targeting Gut Microbiota: Association of JNK/ERK/NF-κB Signaling Pathways. J. Agric. Food Chem. 2021, 35, 10151–10162. [Google Scholar] [CrossRef]
- Feng, Z.; Jiao, L.; Wu, Z.; Xu, J.; Gu, P.; Xu, S.; Liu, Z.; Hu, Y.; Liu, J.; Wu, Y.; et al. A Novel Nanomedicine Ameliorates Acute Inflammatory Bowel Disease by Regulating Macrophages and T-Cells. Mol. Pharm. 2021, 9, 3484–3495. [Google Scholar] [CrossRef]
- Bona, M.; Torres, C.; Lima, S.; Morais, A.; Lima, A.; Maciel, B. Intestinal Barrier Permeability in Obese Individuals with or without Metabolic Syndrome: A Systematic Review. Nutrients 2022, 14, 3649. [Google Scholar] [CrossRef]
- Zhang, J.; Cen, L.; Zhang, X.; Tang, C.; Chen, Y.; Zhang, Y.; Yu, M.; Lu, C.; Li, M.; Li, S.; et al. MPST deficiency promotes intestinal epithelial cell apoptosis and aggravates inflammatory bowel disease via AKT. Redox Biol. 2022, 56, 102469. [Google Scholar] [CrossRef]
- Feng, J.; Wang, Y.; Lv, Y.; Fang, S.; Ren, M.; Yao, M.; Lan, M.; Zhao, Y.; Gao, F. XA pH-Responsive and Colitis-Targeted Nanoparticle Loaded with Shikonin for the Oral Treatment of Inflammatory Bowel Disease in Mice. Mol. Pharm. 2022, 19, 4157–4170. [Google Scholar] [CrossRef]
- Wang, Z.; Klipfell, E.; Bennett, B.J.; Koeth, R.; Levison, B.S.; Dugar, B.; Feldstein, A.E.; Britt, E.B.; Fu, X.; Chung, Y.M. Gut Flora Metabolism Of Phosphatidylcholine Promotes Cardiovascular Disease. Nature 2011, 7341, 57–63. [Google Scholar] [CrossRef]
- Engelhardt, K.R.; Grimbacher, B. IL-10 in Humans: Lessons from the Gut, IL-10/IL-10 Receptor Deficiencies, and IL-10 Polymorphisms. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar] [CrossRef]
- Gori, A.; Tincati, C.; Rizzardini, G.; Torti, C.; Quirino, T.; Haarman, M.; Amor, K.B.; Van Schaik, J.; Vriesema, A.; Knol, J. Early impairment of gut function and gut flora supporting a role for alteration of gastrointestinal mucosa in human immunodeficiency virus pathogenesis. J. Clin. Microbiol. 2008, 2, 757–758. [Google Scholar] [CrossRef]
- Chiou, Y.-S.; Lee, P.-S.; Pan, M.-H. Food Bioactives and Their Effects on Obesity-Accelerated Inflammatory Bowel Disease. J. Agric. Food Chem. 2018, 66, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Dziarski, R.; Park, S.Y.; Kashyap, D.R.; Dowd, S.E.; Gupta, D. Pglyrp-Regulated Gut Microflora Prevotella falsenii, Parabacteroides distasonis and Bacteroides eggerthii Enhance and Alistipes finegoldii Attenuates Colitis in Mice. PLoS ONE 2016, 11, e0146162. [Google Scholar] [CrossRef]
- Zheng, M.; Mao, K.; Fang, D.; Li, D.; Zhu, J. B cell residency but not T cell–independent IgA switching in the gut requires innate lymphoid cells. Proc. Natl. Acad. Sci. USA 2021, 27, e2106754118. [Google Scholar] [CrossRef]
- Akritidou, T.; Akkermans, S.; Smet, C.; Gaspari, S.; Sharma, C.; Matthews, E.; Van Impe, J.F.M. Gut microbiota of the small intestine as an antimicrobial barrier against foodborne pathogens: Impact of diet on the survival of S. Typhimurium and L. monocytogenes during in vitro digestion. Food Res. Int. 2023, 173, 113292. [Google Scholar] [CrossRef] [PubMed]
- Xiang, Y.T.; Mei, L. Research progress in pathogenesis of IBD: The role of gut flora. Prog. Physiol. Sci. 2013, 4, 247–252. [Google Scholar]
- Scaldaferri, F.; Gerardi, V.; Lopetuso, L.R.; Zompo, F.D.; Gasbarrini, A. Gut Microbial Flora, Prebiotics, and Probiotics in IBD: Their Current Usage and Utility. BioMed Res. Int. 2013, 2, 435268. [Google Scholar] [CrossRef]
- Liu, M.-J.; Yang, J.-Y.; Yan, Z.-H.; Hu, S.; Li, J.-Q.; Xu, Z.-X.; Jian, Y.-P. Recent findings in Akkermansia muciniphila-regulated metabolism and its role in intestinal diseases. Clin. Nutr. 2022, 10, 2333–2344. [Google Scholar] [CrossRef]
- Ellul, S.; Rausch, P.; Pisani, A.; Bang, C.; Ellul, P.; Franke, A. P688 Association of Akkermansia muciniphila with a healthy gut microbiome. J. Crohn’s Colitis 2021, 15 (Suppl. S1), S606–S607. [Google Scholar] [CrossRef]
- Derrien, M.; Belzer, C.; de Vos, W.M. Akkermansia Muciniphila and Its Role in Regulating Host Functions. Microb. Pathog. 2016, 106, 171–181. [Google Scholar] [CrossRef]
- Amandine, E.; Clara, B.; Lucie, G.; Janneke, P.O.; Celine, D.; Laure, B.B.; Yves, G.; Muriel, D.; Giulio, G.M.; Nathalie, M.D.; et al. Cross-talk Between Akkermansia muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef]
- Bian, X.; Wu, W.; Yang, L.; Lv, L.; Wang, Q.; Li, Y.; Ye, J.; Fang, D.; Wu, J.; Jiang, X.; et al. Administration of Akkermansia muciniphila Ameliorates Dextran Sulfate Sodium-Induced Ulcerative Colitis in Mice. Front. Microbiol. 2019, 10, 2259. [Google Scholar] [CrossRef]
- Liu, Q.; Lu, W.; Tian, F.; Zhao, J.; Zhang, H.; Hong, K.; Yu, L. Akkermansia muciniphila Exerts Strain-Specific Effects on DSS-Induced Ulcerative Colitis in Mice. Front. Cell. Infect. Microbiol. 2021, 11, 698914. [Google Scholar] [CrossRef]
- Xu, D.; Zhuang, L.; Gao, S.; Ma, H.; Cheng, J.; Liu, J.; Liu, D.; Fu, S.; Hu, G. Orally Administered Ginkgolide C Attenuates DSS-Induced Colitis by Maintaining Gut Barrier Integrity, Inhibiting Inflammatory Responses, and Regulating Gut microbiota. J. Agric. Food Chem. 2022, 46, 14718–14731. [Google Scholar] [CrossRef]
- Liu, C.; Hua, H.; Zhu, H.; Cheng, Y.; Guo, Y.; Yao, W.; Qian, H. Aloe Polysaccharides Ameliorate Acute Colitis in Mice Via Nrf2/HO-1 Signaling Pathway and Short-Chain Fatty Acids Metabolism. Int. J. Biol. Macromol. 2021, 185, 804–812. [Google Scholar] [CrossRef]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; De Los Reyes-gavilán, C.G.; Salazar, N. Intestinal Short Chain Fatty Acids and Their Link with Diet and Human Health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef]
- Kim, C.H.; Park, J.; Kim, M. Gut Microbiota-Derived Short-Chain Fatty Acids, T Cells, and Inflammation. Immune Netw. 2014, 14, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The Microbial Metabolites, Short-Chain Fatty Acids, Regulate Colonic Treg Cell Homeostasis. Science 2013, 341, 569–573. [Google Scholar] [CrossRef]
- Solanki, S.; Lee, J.-H.; Shah, Y. Amino Acid Sensing Pathways in Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2022, 28 (Suppl. S1), S23–S24. [Google Scholar] [CrossRef]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzmanr, I.R.; Lin, J. Butyrate Enhances the Intestinal Barrier by Facilitating Tight Junction Assembly Via Activation of AMP-activated Protein Kinase in Caco-2 Cell Monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef]
- Kelly, C.J.; Zheng, L.; Campbell, E.L.; Saeedi, B.; Scholz, C.C.; Bayless , A.J.; Wilson, K.E.; Glover, L.E.; Kominsky, D.J.; Magnuson, A.; et al. Crosstalk Between Microbiota-Derived Short-Chain Fatty Acids and Intestinal Epithelial HIF Augments Tissue Barrier Function. Biol. Invasions 2015, 17 (Suppl. S1), 662–671. [Google Scholar] [CrossRef]
- Chen, T.; Shen, M.; Yu, Q.; Chen, Y.; Wen, H.; Lu, H.; Chen, S.; Xie, J. Purple Red Rice Anthocyanins Alleviate Intestinal Damage in Cyclophosphamide-Induced Mice Associated with Modulation of Intestinal Barrier Function and Gut Microbiota. Food Chem. 2022, 397, 133768. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stool | Bleeding | Weight Loss (%) | Score |
|---|---|---|---|
| Normal | Normal | Unchanged | 0 |
| - | - | 1–5 | 1 |
| Loose | Occult blood positive | 5–10 | 2 |
| - | - | 10–15 | 3 |
| Diarrhea | Overt bleeding | >15 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Lin, H.; Xiong, Y.; Zhang, Z.; Zeng, L.; Liu, Z. Fu Brick Tea Protects the Intestinal Barrier and Ameliorates Colitis in Mice by Regulating Gut Microbiota. Foods 2025, 14, 1122. https://doi.org/10.3390/foods14071122
Zhang Y, Lin H, Xiong Y, Zhang Z, Zeng L, Liu Z. Fu Brick Tea Protects the Intestinal Barrier and Ameliorates Colitis in Mice by Regulating Gut Microbiota. Foods. 2025; 14(7):1122. https://doi.org/10.3390/foods14071122
Chicago/Turabian StyleZhang, Yangbo, Haiyan Lin, Yifan Xiong, Zhixu Zhang, Li Zeng, and Zhonghua Liu. 2025. "Fu Brick Tea Protects the Intestinal Barrier and Ameliorates Colitis in Mice by Regulating Gut Microbiota" Foods 14, no. 7: 1122. https://doi.org/10.3390/foods14071122
APA StyleZhang, Y., Lin, H., Xiong, Y., Zhang, Z., Zeng, L., & Liu, Z. (2025). Fu Brick Tea Protects the Intestinal Barrier and Ameliorates Colitis in Mice by Regulating Gut Microbiota. Foods, 14(7), 1122. https://doi.org/10.3390/foods14071122