
Interaction Effects of Tannic Acid and Gluten on Bread-Making and Its Starch Digestion



Abstract
1. Introduction
2. Materials and Methods
2.1. Experiment Materials
2.2. Fluorescent Quenching
2.3. Molecular Modeling
2.4. Bread-Making
2.5. Baking Performance
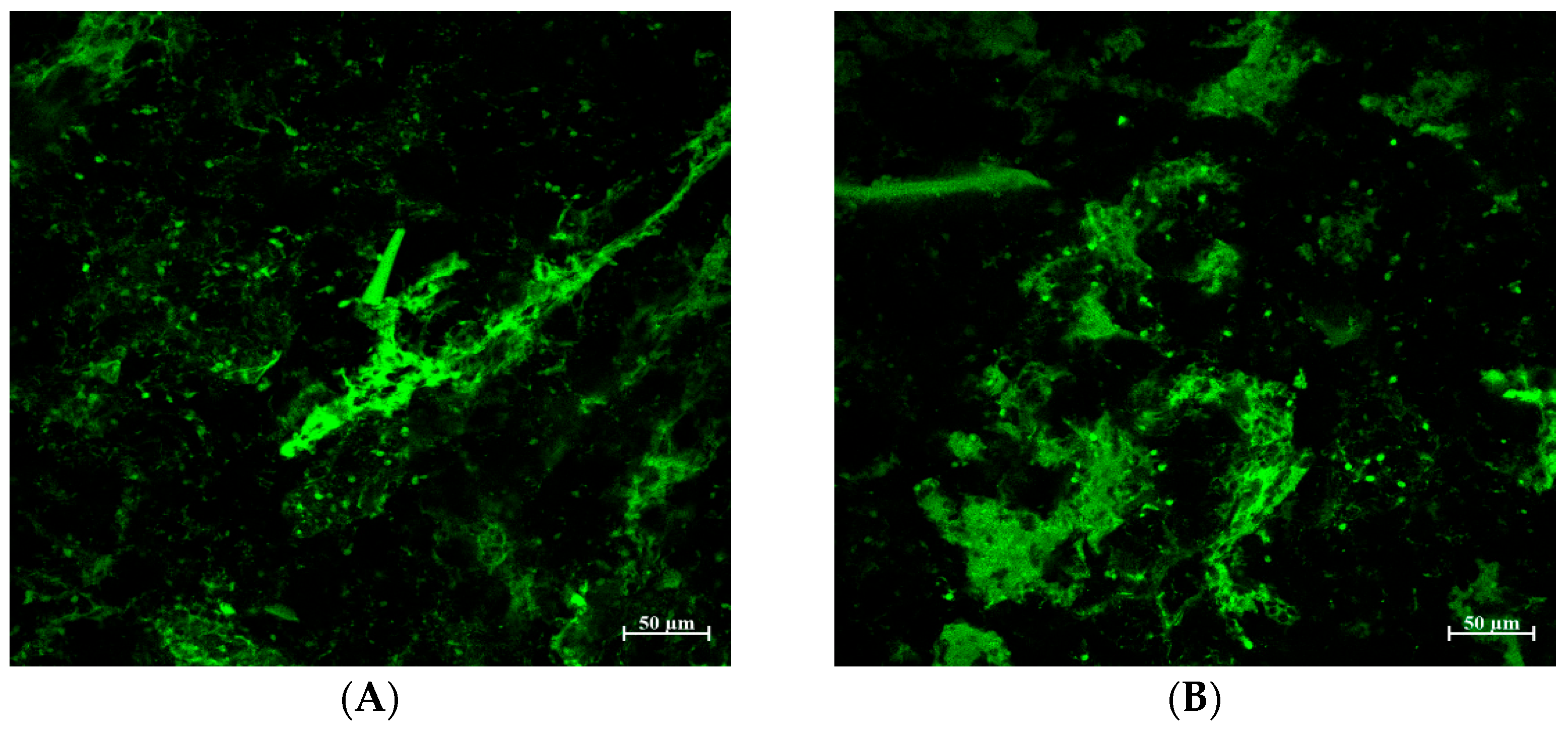
2.6. Confocal Laser Scanning Microscopy
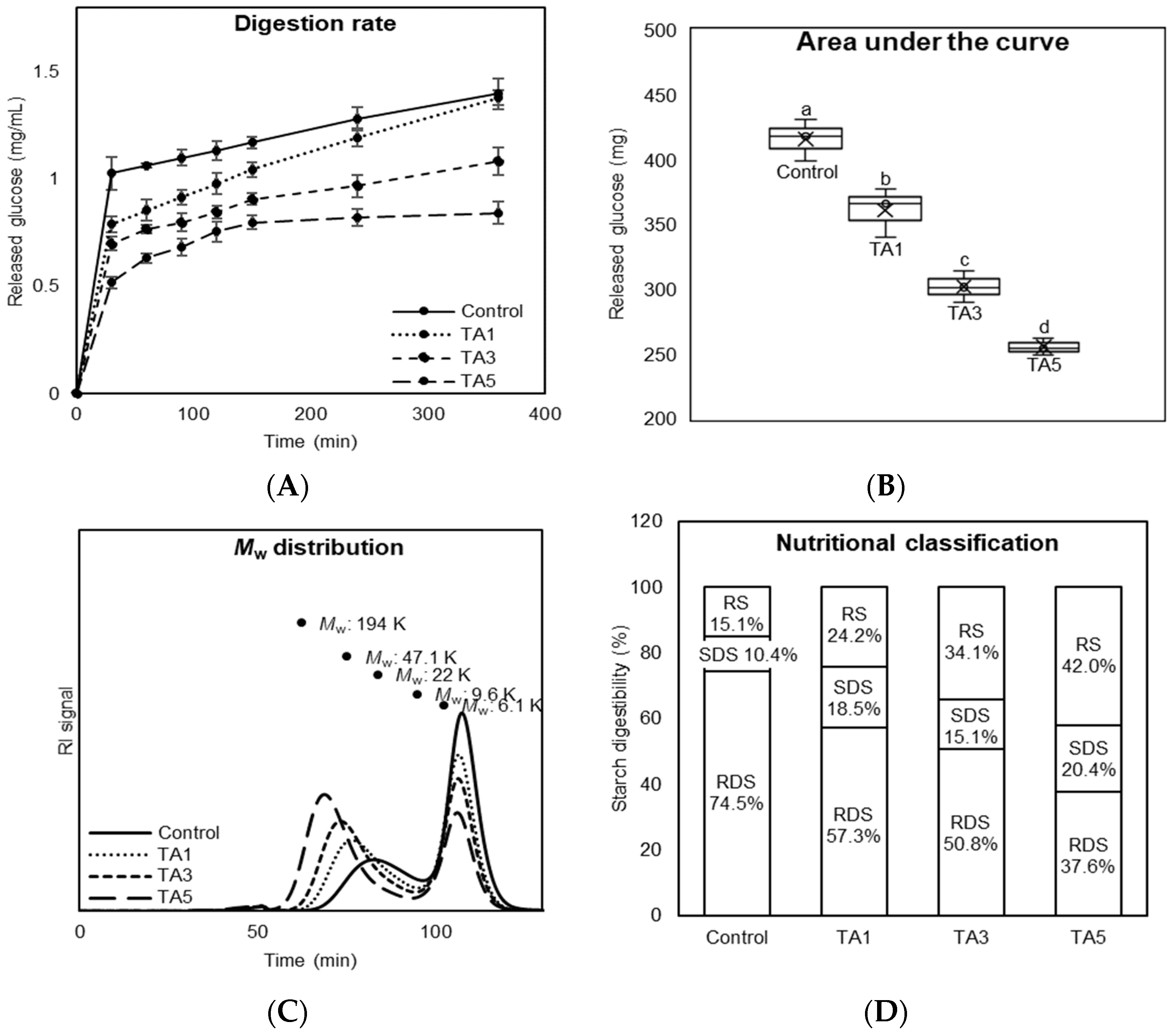
2.7. In Vitro Starch Digestibility
2.8. HPLC for Molecular Degradation of Carbohydrate During an In Vitro Digestion Process
2.9. Statistical Analysis
3. Results
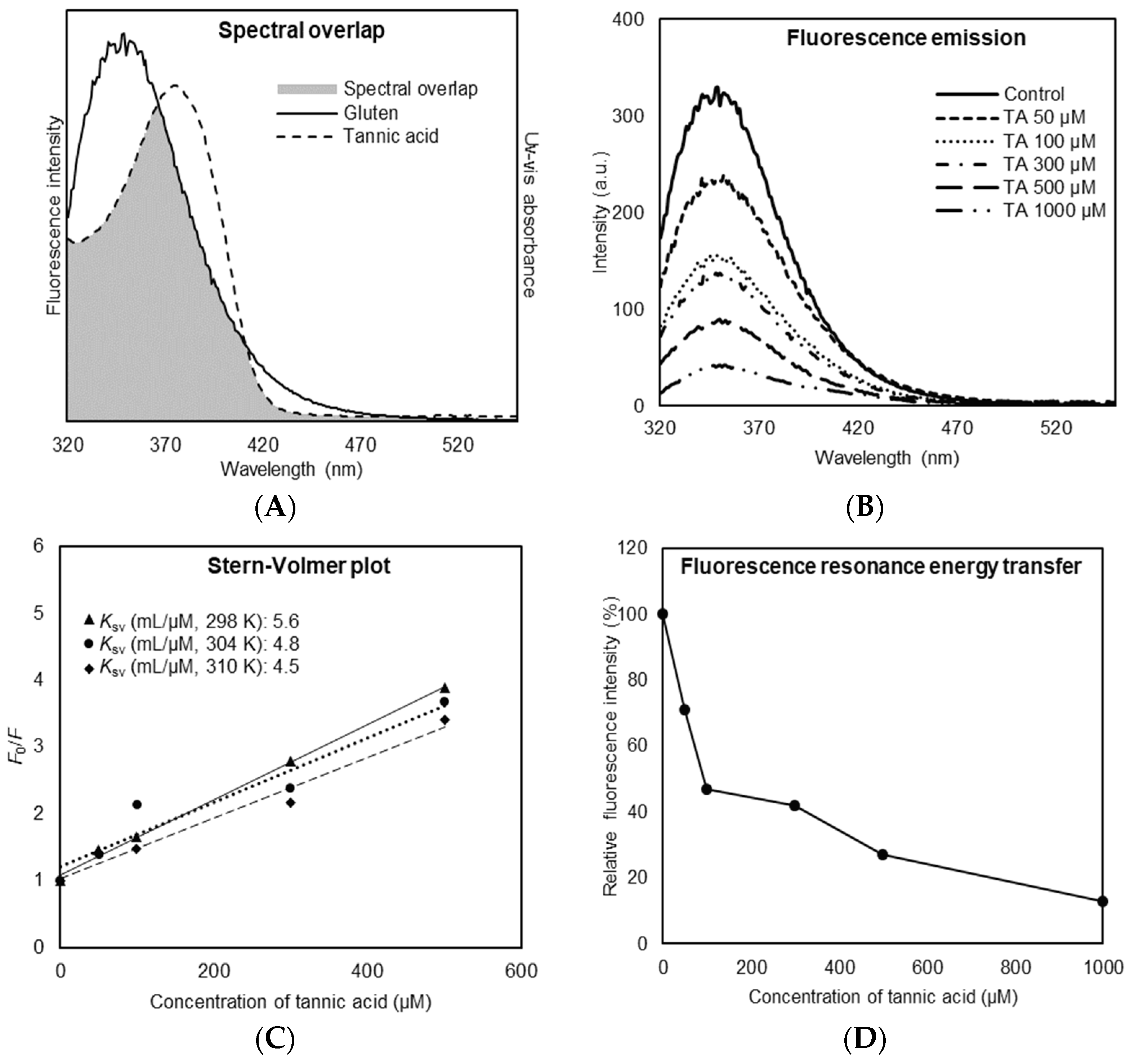
3.1. Binding Affinity of TA Toward Gluten Protein
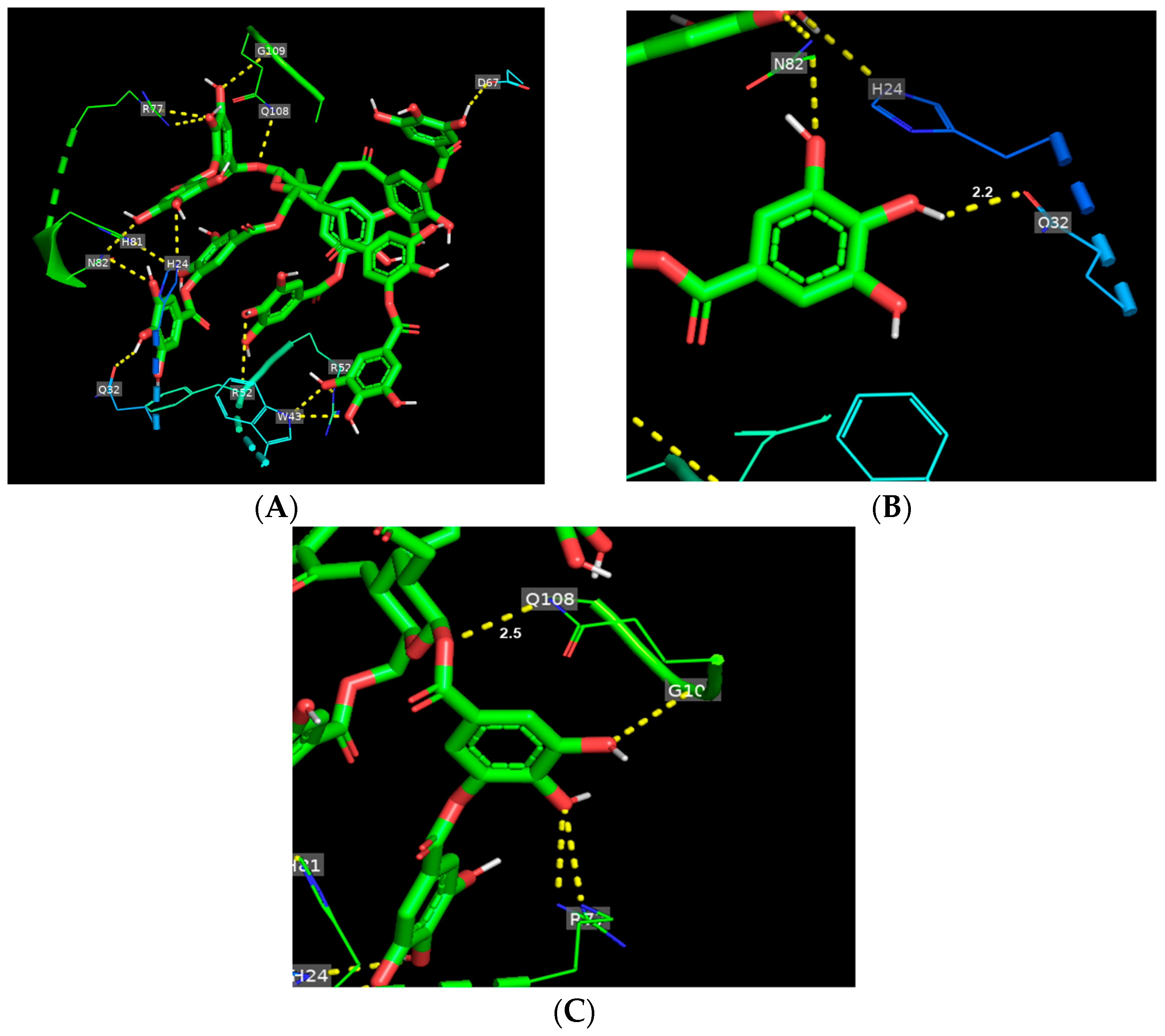
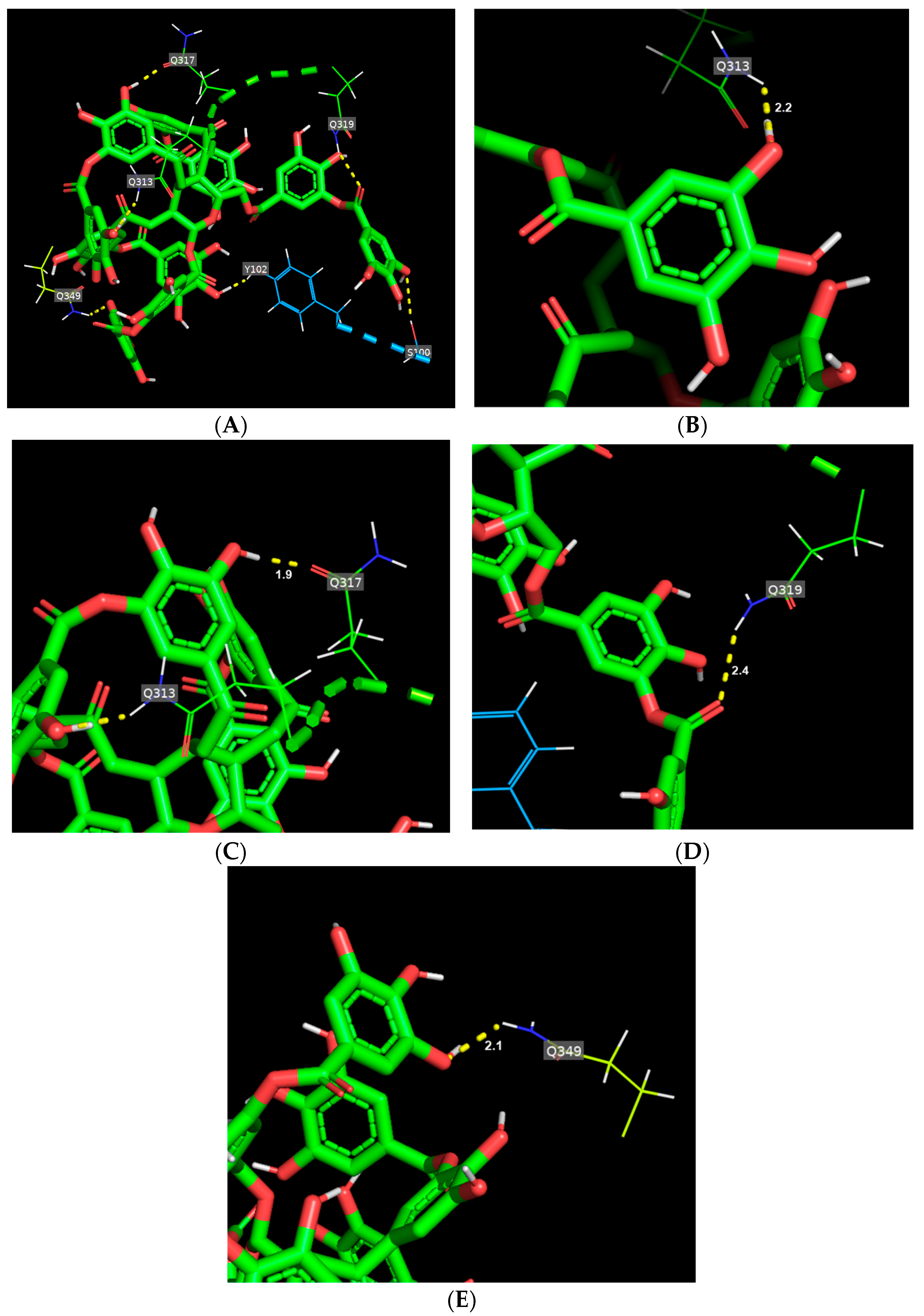
3.2. Binding Mode of TA Toward Gluten Protein
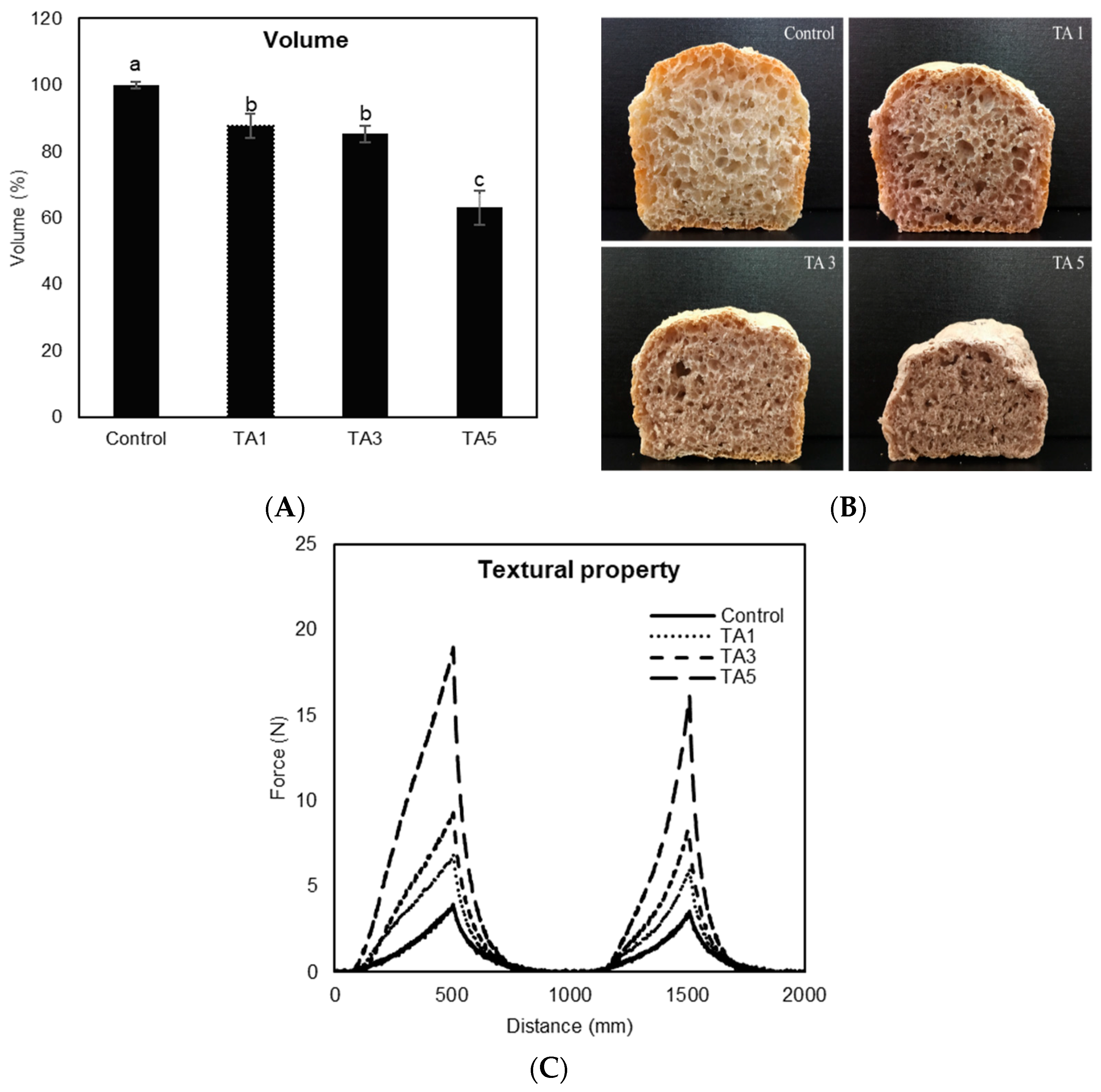
3.3. Baking Performance of TA-Treated Bread
3.4. Protein Structure in Bread Treated with TA
3.5. Starch Digestion Property of TA-Treated Bread
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Englyst, H.N.; Kingman, S.; Cummings, J. Classification and measurement of nutritionally important starch fractions. Eur. J. Clin. Nutr. 1992, 46, S33–S50. [Google Scholar] [PubMed]
- Englyst, H.N.; Veenstra, J.; Hudson, G.J. Measurement of rapidly available glucose (RAG) in plant foods: A potential in vitro predictor of the glycemic response. Br. J. Nutr. 1996, 75, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhou, X.; Xiang, X.; Miao, M. Association of slowly digetible starch intake with reduction of postprandial glycemic response: An update meta-analysis. Foods 2023, 12, 89. [Google Scholar] [CrossRef]
- Sciarini, L.S.; Bustos, M.; Vignola, M.B.; Paesani, C.; Salinas, C.; Perez, G.T. A study on fibre addition to gluten free bread: Its effects on bread quality and in vitro digestibility. J. Food Sci. Technol. 2017, 54, 244–252. [Google Scholar] [CrossRef]
- Freitas, D.; Boué, F.; Benallaoua, M.; Airinei, G.; Benamouzig, R.; Le Feunteun, S. Lemon juice, but not tea, reduces the glycemic response to bread in healthy volunteers: A randomized crossover trial. Eur. J. Nutr. 2021, 60, 113–122. [Google Scholar] [CrossRef]
- Kerimi, A.; Nyambe-Silavwe, H.; Gauer, J.S.; Tomás-Barberán, F.A.; Williamson, G. Pomegranate juice, but not an extract, confers a lower glycemic response on a high–glycemic index food: Randomized, crossover, controlled trials in healthy subjects. Am. J. Clin. Nutr. 2017, 106, 1384–1393. [Google Scholar] [CrossRef]
- Wieser, H. Chemistry of gluten proteins. Food Microbiol. 2007, 24, 115–119. [Google Scholar] [CrossRef]
- Zou, W.; Sissons, M.; Gidley, M.J.; Gilbert, R.G.; Warren, F.J. Combined techniques for characterising pasta structure reveals how the gluten network slows enzymic digestion rate. Food Chem. 2015, 188, 559–568. [Google Scholar] [CrossRef]
- Bordenave, N.; Hamaker, B.R.; Ferruzzi, M.G. Nature and consequences of non-covalent interactions between flavonoids and macronutrients in foods. Food Funct. 2014, 5, 18–34. [Google Scholar] [CrossRef]
- Hager, A.-S.; Vallons, K.J.; Arendt, E.K. Influence of gallic acid and tannic acid on the mechanical and barrier properties of wheat gluten films. J. Agric. Food Chem. 2012, 60, 6157–6163. [Google Scholar] [CrossRef]
- Girard, A.L.; Awika, J.M. Effects of edible plant polyphenols on gluten protein functionality and potential applications of polyphenol–gluten interactions. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2164–2199. [Google Scholar] [CrossRef] [PubMed]
- Girard, A.L.; Bean, S.R.; Tilley, M.; Adrianos, S.L.; Awika, J.M. Interaction mechanisms of condensed tannins (proanthocyanidins) with wheat gluten proteins. Food Chem. 2018, 245, 1154–1162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Cheng, L.; Jiang, L.; Wang, Y.; Yang, G.; He, G. Effects of tannic acid on gluten protein structure, dough properties and bread quality of Chinese wheat. J. Sci. Food Agric. 2010, 90, 2462–2468. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, H. Förster resonance energy transfer–A spectroscopic nanoruler: Principle and applications. J. Photochem. Photobiol. C Photochem. Rev. 2011, 12, 20–30. [Google Scholar] [CrossRef]
- Yang, J.; Yan, R.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput.-Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef]
- Eftink, M.R.; Ghiron, C.A. Fluorescence quenching studies with proteins. Anal. Biochem. 1981, 114, 199–227. [Google Scholar] [CrossRef]
- Kan, L.; Capuano, E.; Fogliano, V.; Oliviero, T.; Verkerk, R. Tea polyphenols as a strategy to control starch digestion in bread: The effects of polyphenol type and gluten. Food Funct. 2020, 11, 5933–5943. [Google Scholar] [CrossRef]
- Wang, J.; Wei, J.; Su, S.; Qiu, J. Novel fluorescence resonance energy transfer optical sensors for vitamin B 12 detection using thermally reduced carbon dots. New J. Chem. 2015, 39, 501–507. [Google Scholar] [CrossRef]
- Khan, S.N.; Islam, B.; Yennamalli, R.; Sultan, A.; Subbarao, N.; Khan, A.U. Interaction of mitoxantrone with human serum albumin: Spectroscopic and molecular modeling studies. Eur. J. Pharm. Sci. 2008, 35, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Mateus, N.; De Freitas, V. Interaction of different polyphenols with bovine serum albumin (BSA) and human salivary α-amylase (HSA) by fluorescence quenching. J. Agric. Food Chem. 2007, 55, 6726–6735. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Zhang, G.; Pan, J.; Wang, Y. α-Glucosidase inhibition by luteolin: Kinetics, interaction and molecular docking. Int. J. Biol. Macromol. 2014, 64, 213–223. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Ma, Y.; Wang, L.; Zhang, Y.; Zhou, J. Multispectroscopic studies on the interaction of maltol, a food additive, with bovine serum albumin. Food Chem. 2012, 133, 264–270. [Google Scholar] [CrossRef]
- Lagrain, B.; Thewissen, B.G.; Brijs, K.; Delcour, J.A. Mechanism of gliadin–glutenin cross-linking during hydrothermal treatment. Food Chem. 2008, 107, 753–760. [Google Scholar] [CrossRef]
- Wang, Y.; Gan, J.; Zhou, Y.; Cheng, Y.; Nirasawa, S. Improving solubility and emulsifying property of wheat gluten by deamidation with four different acids: Effect of replacement of folded conformation by extended structure. Food Hydrocoll. 2017, 72, 105–114. [Google Scholar] [CrossRef]
- Hoseney, R.C. Principles of Cereal Science and Technology; American Association of Cereal Chemists (AACC): St. Paul, MN, USA, 1994. [Google Scholar]
- Sim, S.; Aziah, A.N.; Cheng, L. Quality and functionality of Chinese steamed bread and dough added with selected non-starch polysaccharides. J. Food Sci. Technol. 2015, 52, 303–310. [Google Scholar] [CrossRef]
- Lim, J.; Ferruzzi, M.G.; Hamaker, B.R. Structural requirements of flavonoids for the selective inhibition of α-amylase versus α-glucosidase. Food Chem. 2022, 370, 130981. [Google Scholar] [CrossRef]
- Yuan, W.; Fan, W.; Mu, Y.; Meng, D.; Yan, Z.; Li, Y.; Lv, Z. Baking intervention for the interaction behaviours between bamboo (Phyllostachys heterocycla) leaf flavonoids and gliadin. Ind. Crop. Prod. 2021, 164, 113385. [Google Scholar] [CrossRef]
- Lim, J.; Zhang, X.; Ferruzzi, M.G.; Hamaker, B.R. Starch digested product analysis by HPAEC reveals structural specificity of flavonoids in the inhibition of mammalian α-amylase and α-glucosidases. Food Chem. 2019, 288, 413–421. [Google Scholar] [CrossRef]
- Gellynck, X.; Kühne, B.; Van Bockstaele, F.; Van de Walle, D.; Dewettinck, K. Consumer perception of bread quality. Appetite 2009, 53, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Li, Y.; Sun, F.; Li, X.; Wang, P.; Sun, J.; Zeng, J.; Wang, C.; Hu, W.; Chang, J. Tannins improve dough mixing properties through affecting physicochemical and structural properties of wheat gluten proteins. Food Res. Int. 2015, 69, 64–71. [Google Scholar] [CrossRef]
- Ozturk, O.K.; Kaasgaard, S.G.; Palmén, L.G.; Vidal, B., Jr.; Hamaker, B.R. Protein matrix retains most starch granules within corn fiber from corn wet-milling process. Ind. Crop. Prod. 2021, 165, 113429. [Google Scholar] [CrossRef]
- Baggio, L.L.; Drucker, D.J. Biology of incretins: GLP-1 and GIP. Gastroenterology 2007, 132, 2131–2157. [Google Scholar] [CrossRef]
- Lim, J.; Ferruzzi, M.G.; Hamaker, B.R. Dietary starch is weight reducing when distally digested in the small intestine. Carbohydr. Polym. 2021, 273, 118599. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Color | Control | TA1 | TA3 | TA5 | |
|---|---|---|---|---|---|
| Crust | L | 66.48 b | 67.01 b | 67.11 b | 75.35 a |
| a | 9.63 a | 9.01 a,b | 8.77 b | 4.94 c | |
| b | 28.00 a | 27.04 a | 25.69 b | 16.47 c | |
| DE A | 1.26 c | 2.54 b | 15.28 a | ||
| Crumb | L | 67.11 a | 56.74 c | 60.65 b | 61.60 b |
| a | 0.64 b | 7.50 a | 7.76 a | 7.81 a | |
| b | 19.87 a | 14.06 b | 14.18 b | 13.99 b | |
| DE A | 10.53 b | 11.16 b | 19.90 a | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nam, S.; Ozturk, O.K.; Lim, J. Interaction Effects of Tannic Acid and Gluten on Bread-Making and Its Starch Digestion. Foods 2025, 14, 233. https://doi.org/10.3390/foods14020233
Nam S, Ozturk OK, Lim J. Interaction Effects of Tannic Acid and Gluten on Bread-Making and Its Starch Digestion. Foods. 2025; 14(2):233. https://doi.org/10.3390/foods14020233
Chicago/Turabian StyleNam, Seonghyeon, Oguz K. Ozturk, and Jongbin Lim. 2025. "Interaction Effects of Tannic Acid and Gluten on Bread-Making and Its Starch Digestion" Foods 14, no. 2: 233. https://doi.org/10.3390/foods14020233
APA StyleNam, S., Ozturk, O. K., & Lim, J. (2025). Interaction Effects of Tannic Acid and Gluten on Bread-Making and Its Starch Digestion. Foods, 14(2), 233. https://doi.org/10.3390/foods14020233