

Nutritional Composition and Bioactive Profiles of Farmed and Wild Watermeal (Wolffia globosa)
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Proximate Composition Analysis
2.3. Measuring Chlorophyll Content
2.4. Determination of β-Glucans
2.5. Determination of Total Phenolic Content (TPC)
2.6. Determination of Total Flavonoid Content (TFC)
2.7. Determination of 2,2-Diphenyl-picryl-hydrazyl (DPPH) Radical Scavenging Ability
2.8. Determination of Ferric Reducing Antioxidant Power (FRAP)
2.9. Determination of Phenolic Acid and Flavonoids by HPLC
2.10. Extraction and Determination of Phytosterols
2.11. Determination of Amino Acid Composition
2.12. Determination of Fatty Acid Composition
2.12.1. Lipid Extraction
2.12.2. Fatty Acid Analysis
2.13. Data Analysis
3. Results and Discussion
3.1. Physical Appearance
3.2. Proximate Composition
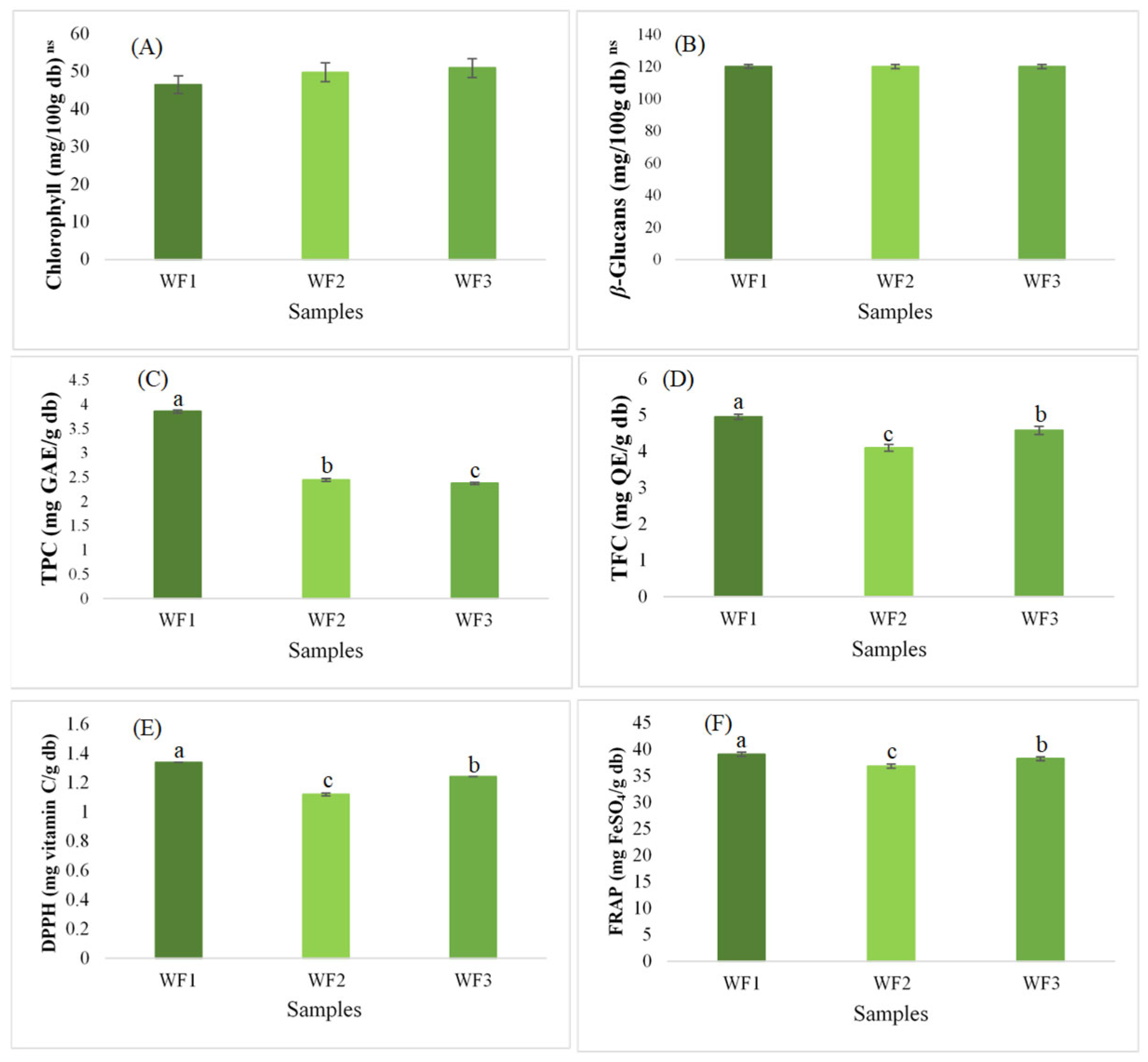
3.3. Chlorophyll, β-Glucans, and Bioactive Compounds
3.4. Phenolic Acids and Flavonoids Contents
3.5. Phytosterols Content
3.6. Amino Acid Compositions
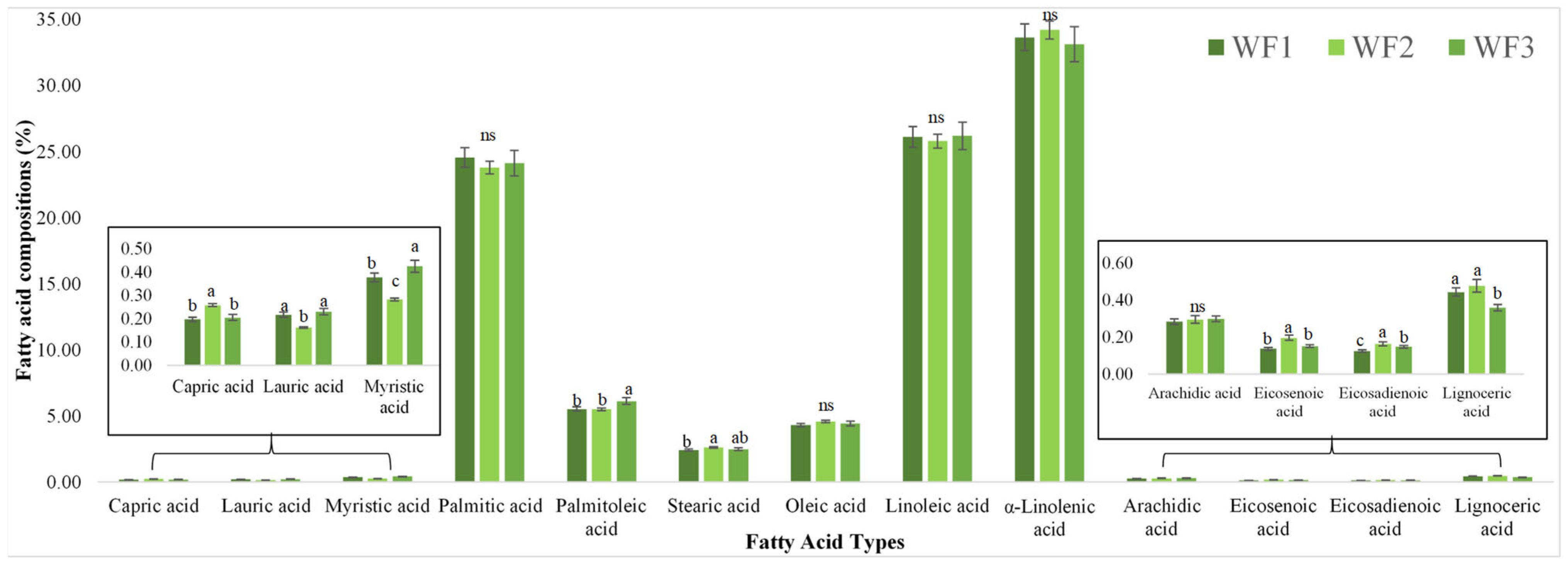
3.7. Fatty Acid Compositions
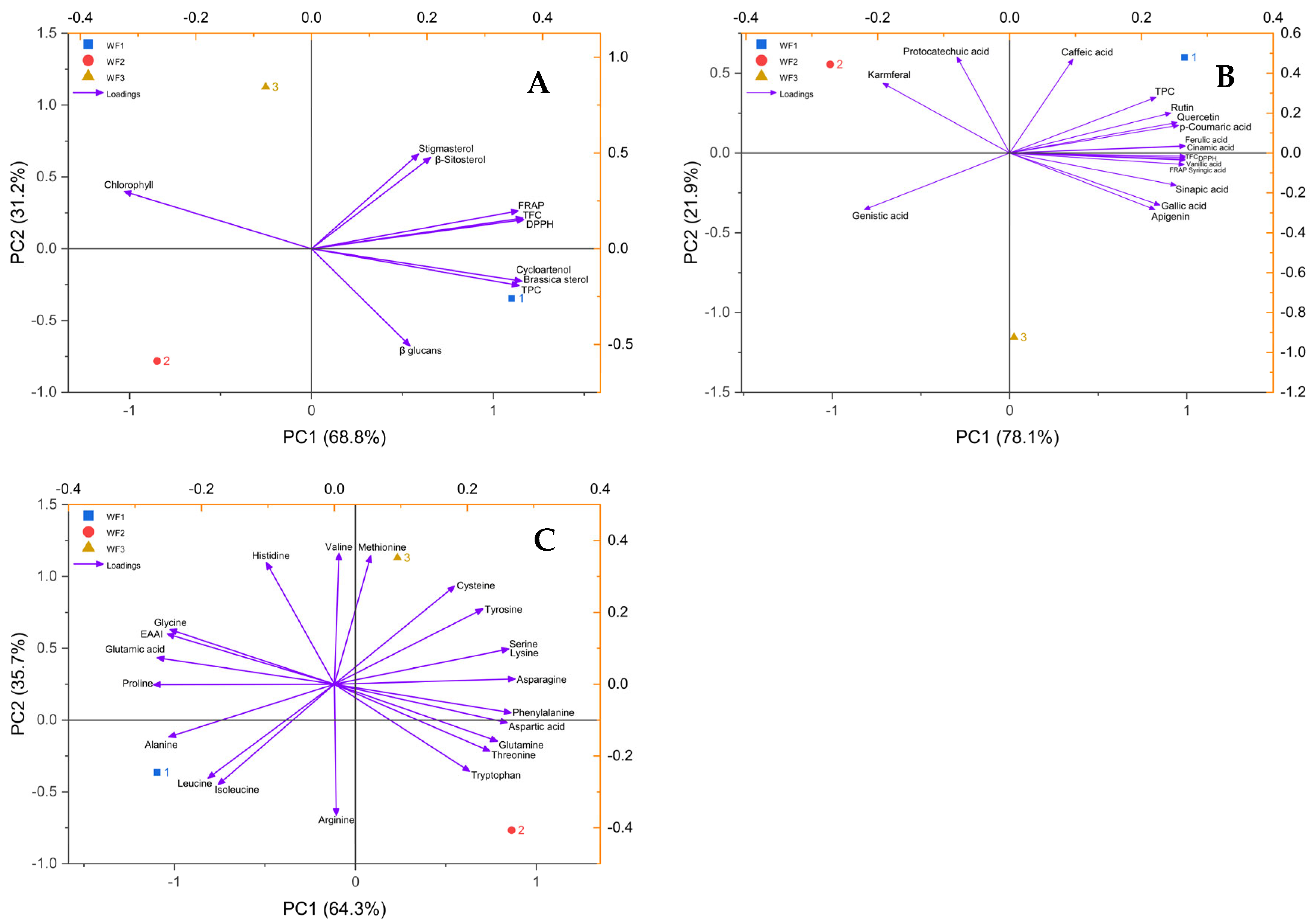
3.8. Principal Component Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boonarsa, P.; Bunyatratchata, A.; Chumroenphat, T.; Thammapat, P.; Chaikwang, T.; Siripan, T.; Li, H.; Siriamornpun, S. Nutritional Quality, Functional Properties, and Biological Characterization of Watermeal (Wolffia globosa). Horticulturae 2024, 10, 1171. [Google Scholar] [CrossRef]
- Siriwat, W.; Ungwiwatkul, S.; Unban, K.; Laokuldilok, T.; Klunklin, W.; Tangjaidee, P.; Potikanond, S.; Kaur, L.; Phongthai, S. Extraction, Enzymatic Modification, and Anti-Cancer Potential of an Alternative Plant-Based Protein from Wolffia globosa. Foods 2023, 12, 3815. [Google Scholar] [CrossRef]
- Romano, L.E.; van Loon, J.J.W.A.; Vincent-Bonnieu, S.; Aronne, G. Wolffia globosa, a Novel Crop Species for Protein Production in Space Agriculture. Sci. Rep. 2024, 14, 27979. [Google Scholar] [CrossRef]
- Yongyod, R.; Kamolrat, N. Effects of Different Nitrogen and Phosphorus Ratios on the Growth, Nutritional Value, and Nutrient Removal Efficiency of Wolffia globosa. Sci 2025, 7, 53. [Google Scholar] [CrossRef]
- Song, Y.; Hu, Z.; Liu, S.; Luo, S.; He, R.; Yang, X.; Li, S.; Yang, X.; An, Y.; Lu, Y. Utilization of Microalgae and Duckweed as Sustainable Protein Sources for Food and Feed: Nutritional Potential and Functional Applications. J. Agric. Food Chem. 2025, 73, 4466–4482. [Google Scholar] [CrossRef]
- Li, X.; Cao, Q.; Liu, G. Advances, Applications, Challenges and Prospects of Alternative Proteins. J. Food Compos. Anal. 2025, 137, 106900. [Google Scholar] [CrossRef]
- Dar, N.A.; Pandit, A.K.; Ganai, B.A. Factors Affecting the Distribution Patterns of Aquatic Macrophytes. Limnol. Rev. 2014, 14, 75–81. [Google Scholar] [CrossRef]
- Xu, J.; Shen, Y.; Zheng, Y.; Smith, G.; Sun, X.S.; Wang, D.; Zhao, Y.; Zhang, W.; Li, Y. Duckweed (Lemnaceae) for Potentially Nutritious Human Food: A Review. Food Rev. Int. 2023, 39, 3620–3634. [Google Scholar] [CrossRef]
- On-Nom, N.; Promdang, P.; Inthachat, W.; Kanoongon, P.; Sahasakul, Y.; Chupeerach, C.; Suttisansanee, U.; Temviriyanukul, P. Wolffia globosa-Based Nutritious Snack Formulation with High Protein and Dietary Fiber Contents. Foods 2023, 12, 2647. [Google Scholar] [CrossRef]
- Yadav, N.K.; Patel, A.B.; Priyadarshi, H.; Baidya, S. Salinity Stress-Induced Impacts on Biomass Production, Bioactive Compounds, Antioxidant Activities and Oxidative Stress in Watermeal (Wolffia globosa). Discov. Appl. Sci. 2025, 7, 106. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Nazir, F.; Maheshwari, C.; Chopra, P.; Chhillar, H.; Sreenivasulu, N. Mineral Nutrients in Plants under Changing Environments: A Road to Future Food and Nutrition Security. Plant Genome 2023, 16, e20362. [Google Scholar] [CrossRef] [PubMed]
- Meade, S.J.; Reid, E.A.; Gerrard, J.A. The Impact of Processing on the Nutritional Quality of Food Proteins. J. AOAC Int. 2005, 88, 904–922. [Google Scholar] [CrossRef]
- Seephua, N.; Liu, Y.; Li, H.; Bunyatratchata, A.; Phuseerit, O.; Siriamornpun, S. Nutritional Enhancement of Rice Noodles with Watermeal (Wolffia globosa). Foods 2025, 14, 1096. [Google Scholar] [CrossRef]
- Taesuk, N.; Wang, A.; Srikaew, M.; Chumroenphat, T.; Barile, D.; Siriamornpun, S.; Bunyatratchata, A. Phytochemical Profiling of Thai Plant-Based Milk Alternatives: Insights into Bioactive Compounds, Antioxidant Activities, Prebiotics, and Amino Acid Abundance. Food Chem. X 2025, 27, 102402. [Google Scholar] [CrossRef] [PubMed]
- Siriamornpun, S.; Tangkhawanit, E.; Kaewseejan, N. Reducing Retrogradation and Lipid Oxidation of Normal and Glutinous Rice Flours by Adding Mango Peel Powder. Food Chem. 2016, 201, 160–167. [Google Scholar] [CrossRef]
- Wanyo, P.; Kaewseejan, N.; Meeso, N.; Siriamornpun, S. Bioactive Compounds and Antioxidant Properties of Different Solvent Extracts Derived from Thai Rice By-Products. Appl. Biol. Chem. 2016, 59, 373–384. [Google Scholar] [CrossRef]
- Butsat, S.; Siriamornpun, S. Antioxidant Capacities and Phenolic Compounds of the Husk, Bran and Endosperm of Thai Rice. Food Chem. 2010, 119, 606–613. [Google Scholar] [CrossRef]
- Li, H.; Liu, Y.; Seephua, N.; Prakitchaiwattana, C.; Liu, R.-X.; Zheng, J.-S.; Siriamornpun, S. Fortification of Cricket and Silkworm Pupae Powders to Improve Nutritional Quality and Digestibility of Rice Noodles. Food Chem. X 2025, 26, 102279. [Google Scholar] [CrossRef]
- Chumroenphat, T.; Somboonwatthanakul, I.; Saensouk, S.; Siriamornpun, S. Changes in Curcuminoids and Chemical Components of Turmeric (Curcuma longa L.) under Freeze-Drying and Low-Temperature Drying Methods. Food Chem. 2021, 339, 128121. [Google Scholar] [CrossRef]
- Thammapat, P.; Meeso, N.; Siriamornpun, S. Effects of NaCl and Soaking Temperature on the Phenolic Compounds, α-Tocopherol, γ-Oryzanol and Fatty Acids of Glutinous Rice. Food Chem. 2015, 175, 218–224. [Google Scholar] [CrossRef]
- Yi, L.; Lakemond, C.M.M.; Sagis, L.M.C.; Eisner-Schadler, V.; Van Huis, A.; Van Boekel, M.A.J.S. Extraction and Characterisation of Protein Fractions from Five Insect Species. Food Chem. 2013, 141, 3341–3348. [Google Scholar] [CrossRef]
- Budavári, N.; Pék, Z.; Helyes, L.; Takács, S.; Nemeskéri, E. An Overview on the Use of Artificial Lighting for Sustainable Lettuce and Microgreens Production in an Indoor Vertical Farming System. Horticulturae 2024, 10, 938. [Google Scholar] [CrossRef]
- Petersen, F.; Demann, J.; Restemeyer, D.; Olfs, H.-W.; Westendarp, H.; Appenroth, K.-J.; Ulbrich, A. Influence of Light Intensity and Spectrum on Duckweed Growth and Proteins in a Small-Scale, Re-Circulating Indoor Vertical Farm. Plants 2022, 11, 1010. [Google Scholar] [CrossRef]
- Hu, H.; Liu, H.; Zhang, H.; Zhu, J.; Yao, X.; Zhang, X.; Zheng, K. Assessment of Chlorophyll Content Based on Image Color Analysis, Comparison with SPAD-502. In Proceedings of the 2010 2nd International Conference on Information Engineering and Computer Science, Wuhan, China, 25–26 December 2010; IEEE: Piscataway, NJ, USA; pp. 1–3. [Google Scholar]
- Appenroth, K.-J.; Sree, K.S.; Böhm, V.; Hammann, S.; Vetter, W.; Leiterer, M.; Jahreis, G. Nutritional Value of Duckweeds (Lemnaceae) as Human Food. Food Chem. 2017, 217, 266–273. [Google Scholar] [CrossRef]
- Appenroth, K.-J.; Sree, K.S.; Bog, M.; Ecker, J.; Seeliger, C.; Böhm, V.; Lorkowski, S.; Sommer, K.; Vetter, W.; Tolzin-Banasch, K.; et al. Nutritional Value of the Duckweed Species of the Genus Wolffia (Lemnaceae) as Human Food. Front. Chem. 2018, 6, 483. [Google Scholar] [CrossRef]
- Ognjenović, M.; Seletković, I.; Marušić, M.; Jonard, M.; Rautio, P.; Timmermann, V.; Tadić, M.P.; Lanšćak, M.; Ugarković, D.; Potočić, N. The Effect of Environmental Factors on the Nutrition of European Beech (Fagus sylvatica L.) Varies with Defoliation. Plants 2022, 12, 168. [Google Scholar] [CrossRef]
- Czerpak, R.; Dobrzyn, P.; Krotke, A.; Kicinska, E. The Effect of Auxins and Salicylic Acid on Chlorophyll and Carotenoid Contents. Pol. J. Environ. Stud. 2002, 11, 231–235. [Google Scholar]
- Brennan, C.S.; Cleary, L.J. The Potential Use of Cereal (1→3,1→4)-β-d-Glucans as Functional Food Ingredients. J. Cereal Sci. 2005, 42, 1–13. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant Phenolics: Extraction, Analysis and Their Antioxidant and Anticancer Properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An Overview. J. Nutr. Sci. 2016, 5, e47. [Google Scholar] [CrossRef]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized Methods for the Determination of Antioxidant Capacity and Phenolics in Foods and Dietary Supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Shahidi, F.; Ambigaipalan, P. Phenolics and Polyphenolics in Foods, Beverages and Spices: Antioxidant Activity and Health Effects—A Review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Pruthvi, G.; Apoorva, M.; Anuthilakesh, T.; Bhargavi, K.; Nithish, G.S.; Achar, R. Food Polyphenols. In Polyphenols; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2023; pp. 1–20. ISBN 978-1-394-18886-4. [Google Scholar]
- Salee, N.; Naruenartwongsakul, S.; Chaiyana, W.; Yawootti, A.; Suthapakti, K.; Simapaisarn, P.; Chaisan, W.; Utama-ang, N. Enhancing Catechins, Antioxidant and Sirtuin 1 Enzyme Stimulation Activities in Green Tea Extract through Pulse Electric Field-Assisted Water Extraction: Optimization by Response Surface Methodology Approach. Heliyon 2024, 10, e36479. [Google Scholar] [CrossRef]
- Bian, Y.; Zhang, Y.; Ruan, L.-Y.; Feng, X.-S. Phytosterols in Plant-Derived Foods: Recent Updates in Extraction and Analysis Methods. Crit. Rev. Anal. Chem. 2024, 1–19. [Google Scholar] [CrossRef]
- Zio, S.; Tarnagda, B.; Tapsoba, F.; Zongo, C.; Savadogo, A. Health Interest of Cholesterol and Phytosterols and Their Contribution to One Health Approach: Review. Heliyon 2024, 10, e40132. [Google Scholar] [CrossRef]
- Kotowska, U.; Piotrowska, A.; Isidorova, A.; Bajguz, A.; Isidorov, V. Gas Chromatographic-Mass Spectrometric Investigation of the Chemical Composition of the Aquatic Plant Wolffia arrhiza (Lemnaceae). Oceanol. Hydrobiol. Stud. 2013, 42, 181–187. [Google Scholar] [CrossRef]
- Piironen, V.; Lindsay, D.G.; Miettinen, T.A.; Toivo, J.; Lampi, A.-M. Plant Sterols: Biosynthesis, Biological Function and Their Importance to Human Nutrition. J. Sci. Food Agric. 2000, 80, 939–966. [Google Scholar] [CrossRef]
- WHO/FAO/UNU. Protein and Amino Acid Requirements in Human Nutrition: Report of a Joint FAO/WHO/UNU Expert Consultation. Available online: https://iris.who.int/handle/10665/43411 (accessed on 10 May 2024).
- Nyangena, D.N.; Mutungi, C.; Imathiu, S.; Kinyuru, J.; Affognon, H.; Ekesi, S.; Nakimbugwe, D.; Fiaboe, K.K.M. Effects of Traditional Processing Techniques on the Nutritional and Microbiological Quality of Four Edible Insect Species Used for Food and Feed in East Africa. Foods 2020, 9, 574. [Google Scholar] [CrossRef]
- Calder, P.C. Functional Roles of Fatty Acids and Their Effects on Human Health. J. Parenter. Enter. Nutr. 2015, 39, 18S–32S. [Google Scholar] [CrossRef] [PubMed]
- Arbex, A.K.; Bizarro, V.R.; Santos, J.C.S.; Araújo, L.M.M.; De Jesus, A.L.C.; Fernandes, M.S.A.; Salles, M.M.; Rocha, D.R.T.W.; Marcadenti, A. The Impact of the Essential Fatty Acids (EFA) in Human Health. Open J. Endocr. Metab. Dis. 2015, 5, 98–104. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient Solution | Concentration (ppm) |
|---|---|
| Nitrogen (N) | 268.59 |
| NH4+ | 30.75 |
| NO3 | 237.84 |
| Phosphorus (P) | 30.04 |
| Potassium (K) | 290.00 |
| Calcium (Ca) | 113.56 |
| Magnesium (Mg) | 30.00 |
| Sulfur (S) | 40.00 |
| Iron (Fe) | 2.40 |
| Boron (B) | 0.51 |
| Manganese (Mn) | 1.63 |
| Zinc (Zn) | 0.44 |
| Copper (Cu) | 0.125 |
| Proximate Composition (g/100 g DW) | WF1 | WF2 | WF3 |
|---|---|---|---|
| Moisture | 4.25 ± 0.24 a | 3.73 ± 0.7 c | 4.06 ± 0.67 b |
| Protein | 22.74 ± 0.66 a | 20.55 ± 0.43 b | 20.85 ± 0.58 b |
| Lipid | 3.23 ± 0.32 b | 4.08 ± 0.24 a | 3.43 ± 0.14 b |
| Ash | 7.84 ± 0.83 a | 6.83 ± 0.35 b | 6.76 ± 0.50 b |
| Fiber ns | 16.53 ± 0.72 | 16.09 ± 0.12 | 15.76 ± 0.09 |
| Carbohydrate | 50.73 ± 1.03 b | 52.44 ± 0.26 a | 51.11 ± 0.62 ab |
| Parameter (µg/g DW) | WF1 | WF2 | WF3 |
|---|---|---|---|
| Phenolic acid content | |||
| Gallic acid ns | 173.04 ± 4.24 | 169.86 ± 3.02 | 173.20 ± 1.22 |
| Protocatechuic acid | 170.90 ± 0.25 a | 172.49 ± 2.67 a | 166.94 ± 1.79 b |
| p-Hydroxybenzoic acid | ND | ND | ND |
| Vanillic acid | 86.20 ± 2.07 a | 78.90 ± 1.25 b | 83.14 ± 2.08 a |
| Caffeic acid ns | 13.69 ± 1.89 | 13.28 ± 1.27 | 12.62 ± 0.44 |
| Syringic acid ns | 10.71 ± 1.95 | 9.61 ± 0.49 | 10.24 ± 0.46 |
| Vanillin | ND | ND | ND |
| p-Coumaric acid | 59.80 ± 2.16 a | 55.36 ± 0.92 b | 56.57 ± 0.92 b |
| Ferulic acid | 22.63 ± 2.52 ab | 18.02 ± 1.88 b | 20.15 ± 1.82 ab |
| Sinapic acid ns | 137.05 ± 2.34 | 129.26 ± 3.74 | 135.60 ± 6.42 |
| Cinamic acid ns | 61.38 ± 3.40 | 57.04 ± 1.35 | 59.02 ± 3.26 |
| Genistic acid | 20.72 ± 0.35 b | 26.29 ± 0.17 a | 26.64 ± 0.17 a |
| Total | 756.12 ± 2.22 a | 646.81 ± 1.68 c | 684.52 ± 2.16 b |
| Flavonoid content | |||
| Rutin | 89.73 ± 1.62 a | 81.30 ± 0.10 b | 82.53 ± 5.30 b |
| Catechin | ND | ND | ND |
| Quercetin ns | 195.36 ± 18.95 | 176.77 ± 5.63 | 181.30 ± 5.39 |
| Apigenin ns | 179.20 ± 26.31 | 172.55 ± 9.15 | 179.98 ± 8.74 |
| Karmferal ns | 39.50 ± 3.90 | 42.46 ± 1.41 | 38.40 ± 3.45 |
| Total | 503.79 ± 14.19 | 473.08 ± 15.23 | 482.21 ± 5.72 |
| Phytosterols (µg/100 g DW) | WF1 | WF2 | WF3 |
|---|---|---|---|
| Campesterol | 122.70 ± 4.70 b | ND | 212.45 ± 1.21 a |
| Stigmasterol | 144.45 ± 1.20 b | ND | 212.90 ± 1.45 a |
| β-Sitosterol | 487.41 ± 3.03 b | 101.96 ± 1.06 c | 625.08 ± 6.30 a |
| Cycloartenol | 43.74 ± 0.60 | ND | ND |
| Brassica sterol | 29.88 ± 0.17 | ND | ND |
| Amino Acids (AAs, g/100 g DW) | WF1 | WF2 | WF3 | WHO/FAO/UNU for Adults [41] |
|---|---|---|---|---|
| Essential amino acids (EAAs) | ||||
| Arginine | 1.95 ± 0.10 b | 2.13 ± 0.05 a | 1.32 ± 0.03 c | - |
| Histidine | 0.34 ± 0.01 b | 0.24 ± 0.01 c | 0.42 ± 0.02 a | 1.5 |
| Isoleucine | 1.32 ± 0.02 a | 1.13 ± 0.03 b | 0.92 ± 0.02 c | 3.0 |
| Leucine | 1.24 ± 0.03 a | 1.04 ± 0.04 b | 0.87 ± 0.02 c | 5.9 |
| Lysine | 0.29 ± 0.02 b | 0.47 ± 0.01 a | 0.46 ± 0.02 a | 4.5 |
| Methionine | 0.14 ± 0.01 b | 0.14 ± 0.01 b | 0.18 ± 0.01 a | 2.2 |
| Phenylalanine | 1.01 ± 0.01 c | 1.36 ± 0.03 a | 1.18 ± 0.01 b | 3.8 |
| Threonine | 0.12 ± 0.01 b | 0.22 ± 0.02 a | 0.14 ± 0.01 b | 2.3 |
| Tryptophan | 0.45 ± 0.01 b | 0.58 ± 0.01 a | 0.45 ± 0.01 b | 0.6 |
| Valine | 1.15 ± 0.04 b | 1.03 ± 0.07 c | 1.67 ± 0.11 a | 3.9 |
| ƩEAAs | 8.26 ± 0.13 a | 8.25 ± 0.18 a | 7.59 ± 0.13 b | - |
| EAAI ns | 0.21 ± 0.01 | 0.18 ± 0.01 | 0.20 ± 0.01 | - |
| Non-Essential amino acids (NEAAs) | ||||
| Alanine | 2.05 ± 010 a | 1.17 ± 0.03 b | 1.07 ± 0.05 c | |
| Asparagine | 0.36 ± 0.02 c | 0.99 ± 0.08 a | 0.81 ± 0.05 b | |
| Aspartic acid | 0.51 ± 0.02 c | 0.75 ± 0.04 a | 0.61 ± 0.01 b | |
| Cysteine | 0.03 ± 0.00 c | 0.05 ± 0.00 b | 0.07 ± 0.00 a | |
| Glutamine | 0.62 ± 0.03 c | 1.08 ± 0.01 a | 0.75 ± 0.03 b | |
| Glutamic acid | 0.77 ± 0.03 a | 0.49 ± 0.03 c | 0.63 ± 0.02 b | |
| Glycine | 0.27 ± 0.01 a | 0.17 ± 0.02 c | 0.24 ± 0.01 b | |
| Proline | 1.00 ± 0.03 a | 0.03 ± 0.00 c | 0.34 ± 0.02 b | |
| Serine | 0.19 ± 0.01 b | 0.37 ± 0.02 a | 0.36 ± 0.01 a | |
| Tyrosine | 1.66 ± 0.03 a | 1.45 ± 0.02 b | 1.57 ± 0.07 a | |
| ƩNEAAs | 7.51 ± 0.21 a | 6.83 ± 0.11 b | 6.42 ± 0.15 c | |
| ƩAAs | 15.77 ± 0.23 a | 15.08 ± 0.21 b | 14.01 ± 0.07 c | |
| ƩEAAs/ƩAAs (%) | 52.37 | 54.70 | 54.16 | |
| ƩNEAAs/ƩAAs (%) | 47.63 | 45.29 | 45.84 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seephua, N.; Boonarsa, P.; Li, H.; Thammapat, P.; Siriamornpun, S. Nutritional Composition and Bioactive Profiles of Farmed and Wild Watermeal (Wolffia globosa). Foods 2025, 14, 1832. https://doi.org/10.3390/foods14101832
Seephua N, Boonarsa P, Li H, Thammapat P, Siriamornpun S. Nutritional Composition and Bioactive Profiles of Farmed and Wild Watermeal (Wolffia globosa). Foods. 2025; 14(10):1832. https://doi.org/10.3390/foods14101832
Chicago/Turabian StyleSeephua, Nidthaya, Parinya Boonarsa, Hua Li, Pornpisanu Thammapat, and Sirithon Siriamornpun. 2025. "Nutritional Composition and Bioactive Profiles of Farmed and Wild Watermeal (Wolffia globosa)" Foods 14, no. 10: 1832. https://doi.org/10.3390/foods14101832
APA StyleSeephua, N., Boonarsa, P., Li, H., Thammapat, P., & Siriamornpun, S. (2025). Nutritional Composition and Bioactive Profiles of Farmed and Wild Watermeal (Wolffia globosa). Foods, 14(10), 1832. https://doi.org/10.3390/foods14101832