
Impact of the Food Matrix on the Antioxidant and Hypoglycemic Effects of Betalains from Red Prickly Pear Juice After In Vitro Digestion

 ,
,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Raw Material
2.2. Betalain Extract Obtention from Red Prickly Pear Juice
2.2.1. Betalain Extraction
2.2.2. Purification of the Betalain Extract by Gel Permeation Chromatography (GPC)
2.3. Aqueous Model System Formulation
2.4. Antioxidant Potential
2.4.1. DPPH Assay
2.4.2. ABTS Assay
2.5. In Vitro Digestion
2.6. Enzymatic Inhibition Assay
2.6.1. α-Amylase Inhibition
- AbsControl is the maximum absorbance of starch without the enzyme (C -);
- AbsSample is the net absorbance of each sample;
- AbsSaBco is the absorbance of each sample without enzyme.
2.6.2. α-Glucosidase Inhibition
2.6.3. Statistical Analysis of the Data
3. Results and Discussion
3.1. Extraction of Betalains from Red Prickly Pear Juice
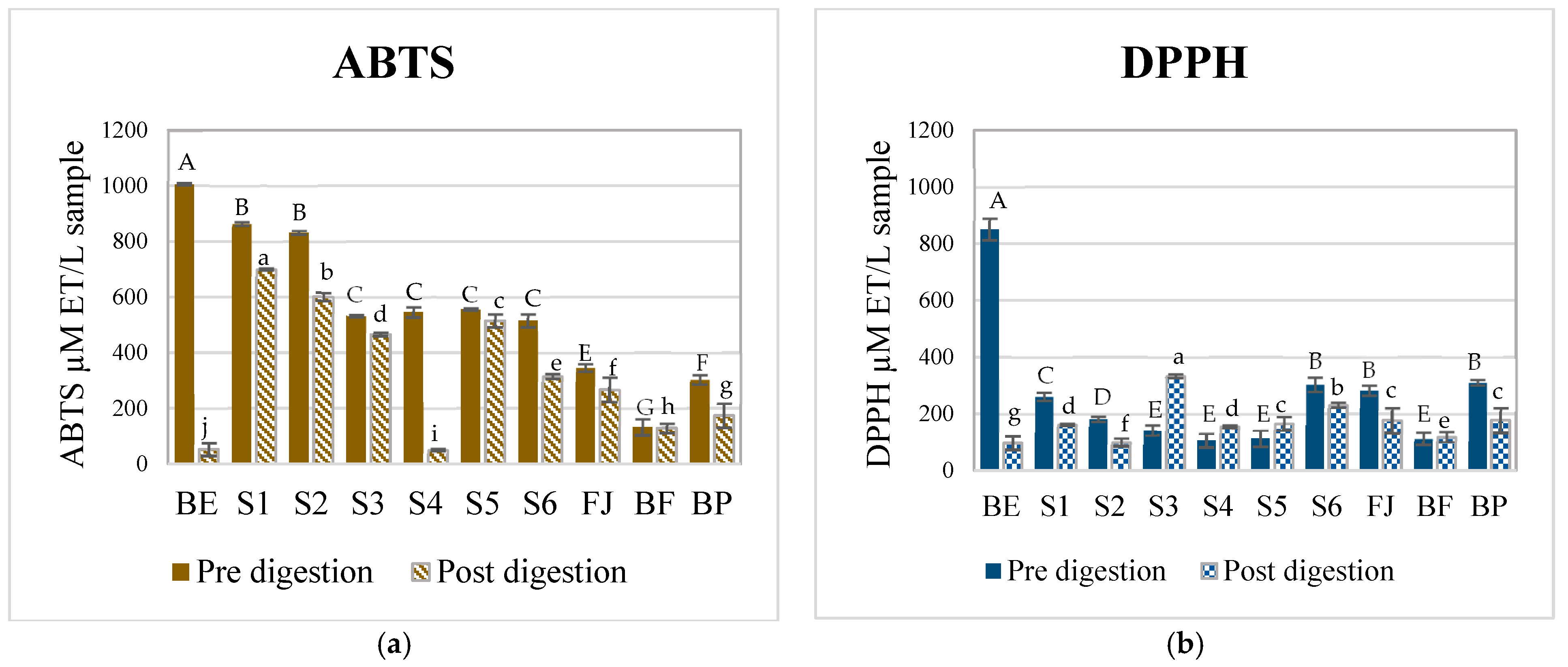
3.2. Antioxidant Potential of Betalains and AMSs
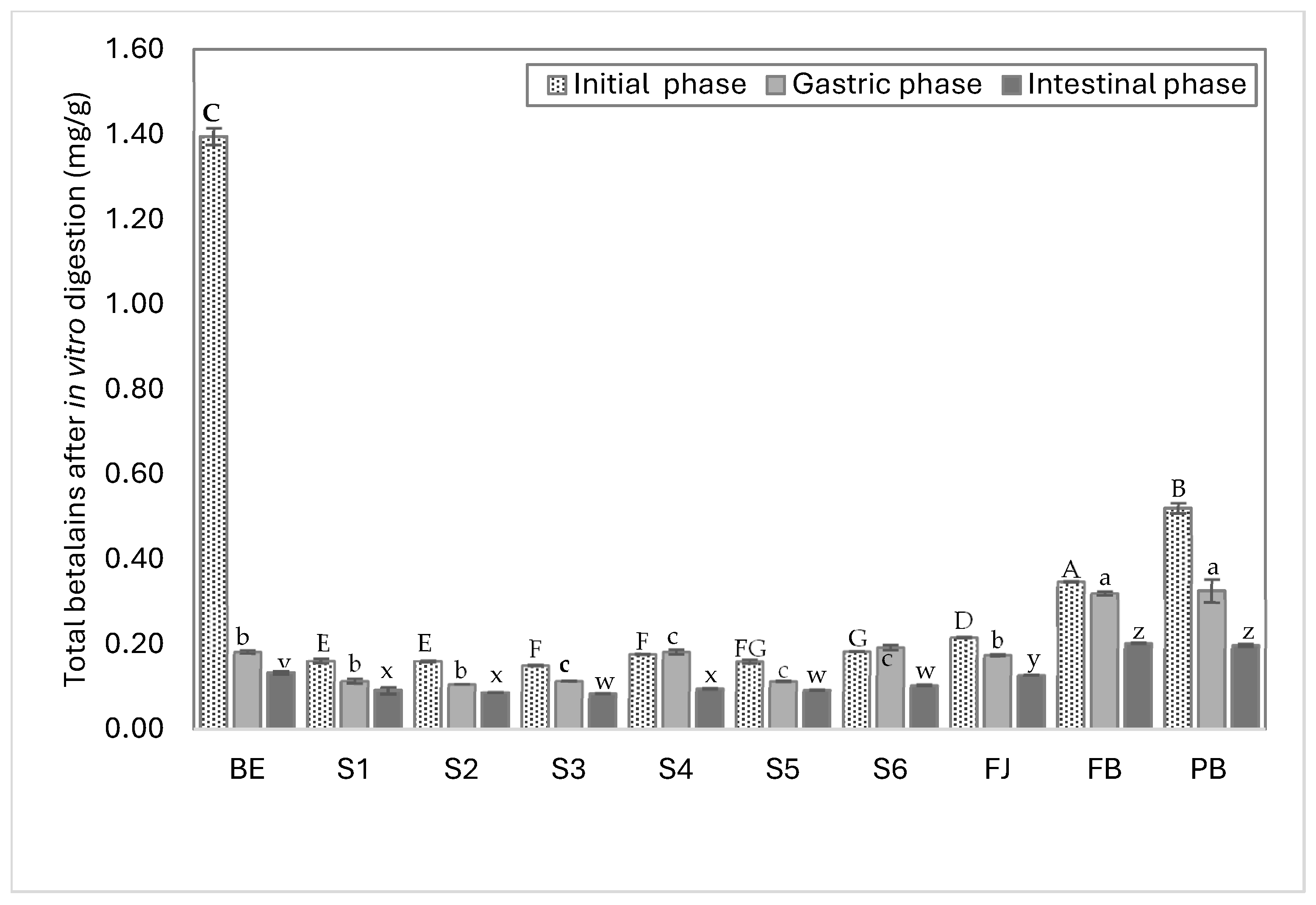
3.3. Betalain Bioaccessibility and Hypoglycemic Effect After In Vitro Digestion
3.3.1. In Vitro Digestion Bioaccessibility
3.3.2. Hypoglycemic Effect
α-Amylase Inhibition
α-Glucosidase Inhibition
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AMS | Aqueous model system |
| BE | Betalain extract |
| BF | Formulated beverage |
| BP | Pasteurized beverage |
| JF | Fresh red prickly pear juice |
| GPC | Gel permeation chromatography |
| G | Glucose |
| M | Mucilage |
| P | Pectin |
| CA | Citric acid |
Appendix A
Appendix A.1

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Initial Phase (mg/g) | Gastric Phase (mg/g) | Intestinal Phase (mg/g) | Bioaccessibility (%) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sample | Betacyanins | Betaxanthins | Total Betalains | Betacyanins | Betaxanthins | Total Betalains | Betacyanins | Betaxanthins | Total Betalains | Betacyanins | Betaxanthins | Total Betalains |
| BE | 0.7700 f ± 0.000 | 0.6252 g ± 0.020 | 1.3952 | 0.1001 d ± 0.002 | 0.0828 c ± 0.001 | 0.1829 | 0.0695 f ± 0.001 | 0.0635 f ± 0.001 | 0.133 | 5.0 a ± 0.001 | 4.6 a ± 0.001 | 9.53 a ± 0.001 |
| S1 | 0.0883 a ± 0.002 | 0.0733 b ± 0.002 | 0.1616 | 0.0601 b ± 0.003 | 0.0543 b ± 0.002 | 0.1144 | 0.0426 a ± 0.001 | 0.0489 d ± 0.007 | 0.0915 | 26.4 c ± 0.001 | 30.2 f ± 0.002 | 56.61 d ± 0.002 |
| S2 | 0.0891 a ± 0.001 | 0.0717 b ± 0.000 | 0.1608 | 0.0554 a ± 0.000 | 0.0514 a ± 0.000 | 0.1068 | 0.0424 a ± 0.000 | 0.0455 b ± 0.000 | 0.0879 | 26.3 c ± 0.000 | 28.3 d ± 0.001 | 54.62 c ± 0.001 |
| S3 | 0.0837 a ± 0.000 | 0.0676 a ± 0.000 | 0.1513 | 0.0613 b ± 0.000 | 0.0528 b ± 0.000 | 0.1141 | 0.0434 a ± 0.000 | 0.0417 a ± 0.000 | 0.0851 | 28.7 f ± 0.000 | 27.6 c ± 0.001 | 56.25 d ± 0.001 |
| S4 | 0.0983 b ± 0.000 | 0.0792 b ± 0.000 | 0.1775 | 0.0983 e ± 0.003 | 0.0846 d ± 0.002 | 0.1829 | 0.0480 c ± 0.000 | 0.0485 d ± 0.001 | 0.0965 | 27.0 d ± 0.000 | 27.3 c ± 0.001 | 54.36 c ± 0.001 |
| S5 | 0.0875 a ± 0.002 | 0.0728 b ± 0.001 | 0.1603 | 0.0603 b ± 0.001 | 0.0535 b ± 0.000 | 0.1138 | 0.0459 b ± 0.000 | 0.0471 c ± 0.001 | 0.093 | 28.7 f ± 0.000 | 29.4 e ± 0.001 | 58.03 e ± 0.001 |
| S6 | 0.1014 c ± 0.000 | 0.0825 c ± 0.000 | 0.1839 | 0.1004 d ± 0.003 | 0.0929 e ± 0.002 | 0.1933 | 0.0508 d ± 0.000 | 0.0534 e ± 0.001 | 0.1042 | 27.6 e ± 0.007 | 29.0 e ± 0.001 | 56.67 d ± 0.001 |
| FJ | 0.1024 c ± 0.001 | 0.1144 d ± 0.000 | 0.2168 | 0.0883 c ± 0.000 | 0.0871 d ± 0.001 | 0.1754 | 0.0585 e ± 0.000 | 0.0697 g ± 0.001 | 0.1282 | 27.0 d ± 0.000 | 32.1 h ± 0.001 | 59.13 f ± 0.001 |
| BF | 0.1884 d ± 0.001 | 0.1599 e ± 0.000 | 0.3483 | 0.1887 e ± 0.002 | 0.1321 f ± 0.001 | 0.3208 | 0.0929 g ± 0.001 | 0.1105 i ± 0.002 | 0.2034 | 26.7 c d ± 0.002 | 31.7 g ± 0.003 | 58.40 e ± 0.003 |
| BP | 0.2803 e ± 0.007 | 0.2402 f ± 0.005 | 0.5205 | 0.1730 e ± 0.016 | 0.1531 f ± 0.000 | 0.3261 | 0.0914 g ± 0.000 | 0.1076 h ± 0.002 | 0.199 | 17.6 b ± 0.007 | 20.7 b ± 0.003 | 38.23 b ± 0.004 |
Appendix A.2
| Pre-Digestion | Post-Digestion | |||||
|---|---|---|---|---|---|---|
| Samples | ABTS (µM ET/L Sample) | DPPH (µM ET/L Sample) | ABTS (µM ET/L Sample) | DPPH (µM ET/L Sample) | Δ ABTS | Δ DPPH |
| BE | 1005.69 a ± 3.47 | 850.79 a± 38.81 | 52.55 i ± 0.03 | 96.84 g ± 0.01 | 953.14 | 753.95 |
| S1 | 863.99 c ± 6.54 | 261.20 b ± 14.43 | 701.07 a ± 0.01 | 162.71 d ± 0.00 | 162.92 | 98.49 |
| S2 | 831.56 d ± 7.05 | 182.15 b ± 8.78 | 601.07 b ± 0.01 | 100.13 f ± 0.01 | 230.49 | 82.02 |
| S3 | 532.58 e ± 4.24 | 142.62 c ± 17.74 | 466.84 d ± 0.01 | 333.99 a ± 0.00 | 65.74 | −191.37 |
| S4 | 545.76 e ± 18.67 | 106.39 c ± 25.04 | 51.09 i ± 0.14 | 156.13 d ± 0.00 | 494.67 | −49.74 |
| S5 | 556.91 ± 4.24 e | 112.98 c± 29.19 | 515.15 c ± 0.0 | 166.01 c ± 0.01 | 41.76 | −53.03 |
| S6 | 515.35 b ± 23.88 | 304.02 c ± 25.04 | 316.06 e ± 0.01 | 231.88 b ± 0.00 | 199.29 | 72.14 |
| FJ | 346.62 b ± 13.55 | 283.15 d ± 17.74 | 267.53 f ± 0.01 | 177.20 c ± 0.02 | 79.09 | 105.95 |
| BF | 132.46 e ± 29.6 | 112.65 f ± 22.22 | 128.64 h ± 0.01 | 119.70 e ± 0.00 | 3.82 | −7.05 |
| BP | 303.59 e ± 16.79 | 310.65 e ± 10.00 | 173.76 g ± 0.00 | 177.20 c ± 0.02 | 129.83 | 133.45 |
References
- Kumari, S.; Kumar, D.; Mittal, M. An Ensemble Approach for Classification and Prediction of Diabetes Mellitus Using Soft Voting Classifier. Int. J. Cogn. Comput. Eng. 2021, 2, 40–46. [Google Scholar] [CrossRef]
- Ogunyemi, O.M.; Gyebi, G.A.; Saheed, A.; Paul, J.; Nwaneri-Chidozie, V.; Olorundare, O.; Adebayo, J.; Koketsu, M.; Aljarba, N.; Alkahtani, S.; et al. Inhibition Mechanism of Alpha-Amylase, a Diabetes Target, by a Steroidal Pregnane and Pregnane Glycosides Derived from Gongronema Latifolium Benth. Front. Mol. Biosci. 2022, 9, 866719. [Google Scholar] [CrossRef] [PubMed]
- Riaz, Z.; Ali, M.N.; Qureshi, Z.; Mohsin, M. In Vitro Investigation and Evaluation of Novel Drug Based on Polyherbal Extract against Type 2 Diabetes. J. Diabetes Res. 2020, 2020, 7357482. [Google Scholar] [CrossRef]
- Golovinskaia, O.; Wang, C.K. The Hypoglycemic Potential of Phenolics from Functional Foods and Their Mechanisms. Food Sci. Hum. Wellness 2023, 12, 986–1007. [Google Scholar] [CrossRef]
- Phung, O.J.; Scholle, J.M.; Talwar, M.; Coleman, C.I. Effect of Noninsulin Antidiabetic Drugs Added to Metformin Therapy on Glycemic Control, Weight Gain, and Hypoglycemia in Type 2 Diabetes. JAMA 2010, 303, 1410–1418. [Google Scholar] [CrossRef]
- Alkhatib, A.; Tsang, C.; Tiss, A.; Bahorun, T.; Arefanian, H.; Barake, R.; Khadir, A.; Tuomilehto, J. Functional Foods and Lifestyle Approaches for Diabetes Prevention and Management. Nutrients 2017, 9, 1310. [Google Scholar] [CrossRef]
- Demir, Y.; Durmaz, L.; Taslimi, P.; Gulçin, İ. Antidiabetic Properties of Dietary Phenolic Compounds: Inhibition Effects on α-Amylase, Aldose Reductase, and α-Glycosidase. Biotechnol. Appl. Biochem. 2019, 66, 781–786. [Google Scholar] [CrossRef]
- Mwakalukwa, R.; Amen, Y.; Nagata, M.; Shimizu, K. Postprandial Hyperglycemia Lowering Effect of the Isolated Compounds from Olive Mill Wastes—An Inhibitory Activity and Kinetics Studies on α-Glucosidase and α-Amylase Enzymes. ACS Omega 2020, 5, 20070–20079. [Google Scholar] [CrossRef]
- Venkatakrishnan, K.; Chiu, H.F.; Wang, C.K. Popular Functional Foods and Herbs for the Management of Type-2-Diabetes Mellitus: A Comprehensive Review with Special Reference to Clinical Trials and Its Proposed Mechanism. J. Funct. Foods 2019, 57, 425–438. [Google Scholar] [CrossRef]
- Madadi, E.; Mazloum-Ravasan, S.; Yu, J.S.; Ha, J.W.; Hamishehkar, H.; Kim, K.H. Therapeutic Application of Betalains: A Review. Plants 2020, 9, 1219. [Google Scholar] [CrossRef]
- Dhananjayan, I.; Kathiroli, S.; Subramani, S.; Veerasamy, V. Ameliorating Effect of Betanin, a Natural Chromoalkaloid by Modulating Hepatic Carbohydrate Metabolic Enzyme Activities and Glycogen Content in Streptozotocin—Nicotinamide Induced Experimental Rats. Biomed. Pharmacother. 2017, 88, 1069–1079. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-carmona, F. Biological Activities of Plant Pigments Betalains. Crit. Rev. Food Sci. Nutr. 2016, 56, 937–945. [Google Scholar] [CrossRef] [PubMed]
- Kaur, G.; Thawkar, B.; Dubey, S.; Jadhav, P. Pharmacological Potentials of Betalains. J. Complement. Integr. Med. 2018, 15, 20170063. [Google Scholar] [CrossRef] [PubMed]
- de Souza, R.L.A.; Santana, M.F.S.; de Macedo, E.M.S.; de Brito, E.S.; Correia, R.T.P. Physicochemical, Bioactive and Functional Evaluation of the Exotic Fruits Opuntia Ficus-Indica AND Pilosocereus Pachycladus Ritter from the Brazilian Caatinga. J. Food Sci. Technol. 2015, 52, 7329–7336. [Google Scholar] [CrossRef]
- Gómez-Maqueo, A.; García-Cayuela, T.; Fernández-López, R.; Welti-Chanes, J.; Cano, M.P. Inhibitory Potential of Prickly Pears and Their Isolated Bioactives against Digestive Enzymes Linked to Type 2 Diabetes and Inflammatory response. J. Sci. Food Agric. 2019, 99, 6380–6391. [Google Scholar] [CrossRef]
- Ota, A.; Ulrih, N.P. An Overview of Herbal Products and Secondary Metabolites Used for Management of Type Two Diabetes. Front. Pharmacol. 2017, 8, 436. [Google Scholar] [CrossRef]
- Čepo, D.V.; Radić, K.; Šalov, M.; Turčić, P.; Anić, D.; Komar, B. Food (Matrix) Effects on Bioaccessibility and Intestinal Permeability of Major Olive Antioxidants. Foods 2020, 9, 1831. [Google Scholar] [CrossRef]
- Santos, D.I.; Saraiva, J.M.A.; Vicente, A.A.; Moldão-martins, M. Methods for Determining Bioavailability and Bioaccessibility of Bioactive Compounds and Nutrients. In Innovative Thermal and Non-Thermal Processing, Bioaccessibility and Bioavailability of Nutrients and Bioactive Compounds; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 23–54. ISBN 9780128141748. [Google Scholar]
- Tesoriere, L.; Gentile, C.; Angileri, F.; Attanzio, A.; Tutone, M.; Allegra, M.; Livrea, M.A. Trans-Epithelial Transport of the Betalain Pigments Indicaxanthin and Betanin across Caco-2 Cell Monolayers and Influence of Food Matrix. Eur. J. Nutr. 2013, 52, 1077–1087. [Google Scholar] [CrossRef]
- Trych, U.; Buniowska-Olejnik, M.; Marszałek, K. Bioaccessibility of Betalains in Beetroot (Beta vulgaris L.) Juice under Different High-Pressure Techniques. Molecules 2022, 27, 7093. [Google Scholar] [CrossRef]
- Castro Enríquez, D.D. Encapsulación de Betalaínas En Una Matriz de Gluteninas Aplicadas Sobre Una Película de Ácido Poliláctico Modificada y Su Evaluación Como Un Sensor de PH En Alimentos. Ph.D. Thesis, Universidad de Sonora, Hermosillo, Mexico, 2020. [Google Scholar]
- Gómez-Maqueo, A.; Welti-Chanes, J.; Cano, M.P. Release Mechanisms of Bioactive Compounds in Fruits Submitted to High Hydrostatic Pressure: A Dynamic Microstructural Analysis Based on Prickly Pear Cells. Food Res. Int. 2020, 130, 108909. [Google Scholar] [CrossRef]
- Martínez-Rodríguez, P.; Guerrero-Rubio, M.A.; Henarejos-Escudero, P.; García-Carmona, F.; Gandía-Herrero, F. Health-Promoting Potential of Betalains in Vivo and Their Relevance as Functional Ingredients: A Review. Trends Food Sci. Technol. 2022, 122, 66–82. [Google Scholar] [CrossRef]
- Montiel-Sánchez, M.; García-Cayuela, T.; Gómez-Maqueo, A.; García, H.S.; Cano, M.P. In Vitro Gastrointestinal Stability, Bioaccessibility and Potential Biological Activities of Betalains and Phenolic Compounds in Cactus Berry Fruits (Myrtillocactus geometrizans). Food Chem. 2020, 342, 128087. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Pan, Y. Influence of Food Matrix and Food Processing on the Chemical Interaction and Bioaccessibility of Dietary Phytochemicals: A Review. Crit. Rev. Food Sci. Nutr. 2022, 62, 6421–6445. [Google Scholar] [CrossRef] [PubMed]
- Clara, C.A.; Angélica, R.G.; Hugo, O.C.V.; Fernando, D.D.L.S.; Lorena, Á.R.E.; Clara, P.Z.; Del Carmen, S.P.E.; Alberto, M.E.J. Chemical Characterization, Antioxidant Capacity, and Anti-Hyperglycemic Effect of Stenocereus Stellatus Fruits from the Arid Mixteca Baja Region of Mexico. Food Chem. 2020, 328, 127076. [Google Scholar] [CrossRef]
- Gonçalves, L.C.P.; Lopes, N.B.; Augusto, F.A.; Pioli, R.M.; Machado, C.O.; Freitas-Dörr, B.C.; Suffredini, H.B.; Bastos, E.L. Phenolic Betalain as Antioxidants: Meta Means More. Pure Appl. Chem. 2020, 92, 243–253. [Google Scholar] [CrossRef]
- Ayala Bendezú, T. Proyecto de Instalación de Una Planta de Procesamiento de Tuna En El Distrito de Chincho Provincia de Angaraes Departamento de Huancavelica; Universidad Nacional Mayor de San marcos: Lima, Peru, 2008. [Google Scholar]
- Virgen-Carrillo, C.A.; Valdés Miramontes, E.H.; Fonseca Hernández, D.; Luna-Vital, D.A.; Mojica, L. West Mexico Berries Modulate α-Amylase, α-Glucosidase and Pancreatic Lipase Using In Vitro and In Silico Approaches. Pharmaceuticals 2022, 15, 1081. [Google Scholar] [CrossRef]
- Domínguez-Murillo, A.C.; Urías-Silvas, J.E. Fermented Coconut Jelly as a Probiotic Vehicle, Physicochemical and Microbiology Characterisation during an in Vitro Digestion. Int. J. Food Sci. Technol. 2023, 58, 45–52. [Google Scholar] [CrossRef]
- Mojica, L.; Gonzalez, E.; Mejia, D.; Granados-silvestre, M.Á.; Menjivar, M. Evaluation of the Hypoglycemic Potential of a Black Bean Hydrolyzed Protein Isolate and Its Pure Peptides Using in Silico, in Vitro and in Vivo Approaches. J. Funct. Foods 2017, 31, 274–286. [Google Scholar] [CrossRef]
- Oluwagunwa, O.A.; Alashi, A.M.; Aluko, R.E. Inhibition of the in Vitro Activities of α-Amylase and Pancreatic Lipase by Aqueous Extracts of Amaranthus Viridis, Solanum Macrocarpon and Telfairia Occidentalis Leaves. Front. Nutr. 2021, 8, 772903. [Google Scholar] [CrossRef]
- Merck Enzymatic Assay of α-Amylase (EC 3.2.1.1). Available online: https://teams.microsoft.com/l/message/19:d1bab01c-0d84-4805-bf94-beeb43d40288_f0958689-fff2-4aab-a3d1-8529e18dc0c6@unq.gbl.spaces/1747275707127?context=%7B%22contextType%22%3A%22chat%22%7D (accessed on 6 May 2025).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Wickham, H.; Romain, F.; Lionel, H.; Kirill, M.; Davis, V. Dplyr: A Grammar of Data Manipulation 2023. Available online: https://dplyr.tidyverse.org/ (accessed on 6 May 2025).
- Graves, S.; Hans-Peter, P.; Sundar, D.-R. MultcompView: Visualizations of Paired Comparisons 2024. Available online: https://cloud.r-project.org/web/packages/multcompView/multcompView.pdf (accessed on 6 May 2025).
- Herbach, K.M.; Stintzing, F.C.; Carle, R. Betalain Stability and Degradation—Structural and Chromatic Aspects. J. Food Sci. 2006, 71, R41–R50. [Google Scholar] [CrossRef]
- Herbach, K.M.; Rohe, M.; Stintzing, F.C.; Carle, R. Structural and Chromatic Stability of Purple Pitaya (Hylocereus polyrhizus Britton & Rose) Betacyanins as Affected by the Juice Matrix and Selected Additives. Food Res. Int. 2006, 39, 667–677. [Google Scholar] [CrossRef]
- Platzer, M.; Kiese, S.; Herfellner, T.; Schweiggert-Weisz, U.; Miesbauer, O.; Eisner, P. Common Trends and Differences in Antioxidant Activity Analysis of Phenolic Substances Using Single Electron Transfer Based Assays. Molecules 2021, 26, 1244. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. The Role of Phenolic Hydroxy Groups in the Free Radical Scavenging Activity of Betalains. J. Nat. Prod. 2009, 72, 1142–1146. [Google Scholar] [CrossRef] [PubMed]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Purification and Antiradical Properties of the Structural Unit of Betalains. J. Nat. Prod. 2012, 75, 1030–1036. [Google Scholar] [CrossRef]
- Gandía-Herrero, F.; Escribano, J.; García-Carmona, F. Structural Implications on Color, Fluorescence, and Antiradical Activity in Betalains. Planta 2010, 232, 449–460. [Google Scholar] [CrossRef]
- Guerrero-Rubio, M.A.; Escribano, J.; García-Carmona, F.; Gandía-Herrero, F. Light Emission in Betalains: From Fluorescent Flowers to Biotechnological Applications. Trends Plant Sci. 2020, 25, 159–175. [Google Scholar] [CrossRef]
- Kanner, J.; Harel, S.; Granit, R. Betalains—A New Class of Dietary Cationized Antioxidants. J. Agric. Food Chem. 2001, 49, 5178–5185. [Google Scholar] [CrossRef]
- Slimen, I.B.; Najar, T.; Abderrabba, M. Chemical and Antioxidant Properties of Betalains. J. Agric. Food Chem. 2017, 65, 675–689. [Google Scholar] [CrossRef]
- Gómez-Maqueo, A.; Antunes-Ricardo, M.; Welti-Chanes, J.; Cano, M.P. Digestive Stability and Bioaccessibility of Antioxidants in Prickly Pear Fruits from the Canary Islands: Healthy Foods and Ingredients. Antioxidants 2020, 9, 164. [Google Scholar] [CrossRef]
- Gómez-Maqueo, A.; Steurer, D.; Welti-Chanes, J.; Cano, M.P. Bioaccessibility of Antioxidants in Prickly Pear Fruits Treated with High Hydrostatic Pressure: An Application for Healthier Foods. Molecules 2021, 26, 5252. [Google Scholar] [CrossRef]
- Igual, M.; Fernandes, Â.; Dias, M.I.; Pinela, J.; García-Segovia, P.; Martínez-Monzó, J.; Barros, L. The In Vitro Simulated Gastrointestinal Digestion Affects the Bioaccessibility and Bioactivity of Beta vulgaris Constituents. Foods 2023, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- López-Palestina, C.U.; Aguirre-Mancilla, C.L.; Raya-Pérez, J.C.; Ramírez-Pimentel, J.G.; Gutiérrez-Tlahque, J.; Hernández-Fuentes, A.D. The Effect of an Edible Coating with Tomato Oily Extract on the Physicochemical and Antioxidant Properties of Garambullo (Myrtillocactus geometrizans) Fruits. Agronomy 2018, 8, 248. [Google Scholar] [CrossRef]
- Sadowska-Bartosz, I.; Bartosz, G. Biological Properties and Applications of Betalains. Molecules 2021, 26, 2520. [Google Scholar] [CrossRef]
- Tesoriere, L.; Fazzari, M.; Angileri, F.; Gentile, C.; Livrea, M.A. In Vitro Digestion of Betalainic Foods. Stability and Bioaccessibility of Betaxanthins and Betacyanins and Antioxidative Potential of Food Digesta. J. Agric. Food Chem. 2008, 56, 10487–10492. [Google Scholar] [CrossRef]
- Ferreira, R.M.; Amaral, R.A.; Silva, A.M.S.; Cardoso, S.M.; Saraiva, J.A. Effect of High-Pressure and Thermal Pasteurization on Microbial and Physico-Chemical Properties of Opuntia Ficus-Indica Juices. Beverages 2022, 8, 84. [Google Scholar] [CrossRef]
- Glitz, C.; Dyekjær, J.D.; Vaitkus, D.; Babaei, M.; Welner, D.H.; Borodina, I. Screening of Plant UDP-Glycosyltransferases for Betanin Production in Yeast. Appl. Biochem. Biotechnol. 2025, 197, 2356–2382. [Google Scholar] [CrossRef]
- Khan, M.I. Plant Betalains: Safety, Antioxidant Activity, Clinical Efficacy, and Bioavailability. Compr. Rev. Food Sci. Food Saf. 2016, 15, 316–330. [Google Scholar] [CrossRef]
- Rodriguez, S.A.; Baumgartner, M.T. Betanidin pKa Prediction Using DFT Methods. ACS Omega 2020, 5, 13751–13759. [Google Scholar] [CrossRef]
- Sawicki, T.; Martinez-villaluenga, C.; Frias, J.; Wiczkowski, W.; Peñas, E.; Baczek, N.; Zielinski, H. The Effect of Processing and in Vitro Digestion on the Betalain Profile and ACE Inhibition Activity of Red Beetroot Products. J. Funct. Foods 2019, 55, 229–237. [Google Scholar] [CrossRef]
- Payan, F. Structural Basis for the Inhibition of Mammalian and Insect α-Amylases by Plant Protein Inhibitors. Biochim. Et Biophys. Acta Proteins Proteom. 2004, 1696, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Askarzadeh, M.; Azizian, H.; Adib, M.; Mohammadi-Khanaposhtani, M.; Mojtabavi, S.; Faramarzi, M.A.; Sajjadi-Jazi, S.M.; Larijani, B.; Hamedifar, H.; Mahdavi, M. Design, Synthesis, in Vitro α-Glucosidase Inhibition, Docking, and Molecular Dynamics of New Phthalimide-Benzenesulfonamide Hybrids for Targeting Type 2 Diabetes. Sci. Rep. 2022, 12, 10569. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Mauldin, R.; Byers, L. Multiple Sugar Binding Sites in A-Glucosidase. Biochim. Et Biophys. Acta (BBA)-Proteins Proteom. 2003, 1645, 22–29. [Google Scholar] [CrossRef]
- Martinez, R.M.; Melo, C.P.B.; Pinto, I.C.; Mendes-Pierotti, S.; Vignoli, J.A.; Verri, W.A.; Casagrande, R. Betalains: A Narrative Review on Pharmacological Mechanisms Supporting the Nutraceutical Potential Towards Health Benefits. Foods 2024, 13, 3909. [Google Scholar] [CrossRef] [PubMed]
- Allegra, M.; Tutone, M.; Tesoriere, L.; Almerico, A.M.; Culletta, G.; Livrea, M.A.; Attanzio, A. Indicaxanthin, a Multi-Target Natural Compound from Opuntia Ficus-Indica Fruit: From Its Poly-Pharmacological Effects to Biochemical Mechanisms and Molecular Modelling Studies. Eur. J. Med. Chem. 2019, 179, 753–764. [Google Scholar] [CrossRef]
- Clifford, T.; Constantinou, C.M.; Keane, K.M.; West, D.J.; Howatson, G.; Stevenson, E.J. The Plasma Bioavailability of Nitrate and Betanin from Beta vulgaris rubra in Humans. Eur. J. Nutr. 2017, 56, 1245–1254. [Google Scholar] [CrossRef]
- Slámová, K.; Kapešová, J.; Valentová, K. “Sweet Flavonoids”: Glycosidase-Catalyzed Modifications. Int. J. Mol. Sci. 2018, 19, 2126. [Google Scholar] [CrossRef]
- Liang, D.M.; Liu, J.H.; Wu, H.; Wang, B.B.; Zhu, H.J.; Qiao, J.J. Glycosyltransferases: Mechanisms and Applications in Natural Product Development. Chem. Soc. Rev. 2015, 44, 8350–8374. [Google Scholar] [CrossRef]
- Assefa, S.T.; Yang, E.Y.; Chae, S.Y.; Song, M.; Lee, J.; Cho, M.C.; Jang, S. Alpha Glucosidase Inhibitory Activities of Plants with Focus on Common Vegetables. Plants 2020, 9, 2. [Google Scholar] [CrossRef]
- Üst, Ö.; Yalçin, E.; Çavuşoğlu, K.; Özkan, B. LC–MS/MS, GC–MS and Molecular Docking Analysis for Phytochemical Fingerprint and Bioactivity of Beta vulgaris L. Sci. Rep. 2024, 14, 7491. [Google Scholar] [CrossRef]



| AMS | Components | Type | Complexity | |
|---|---|---|---|---|
| BE | Betalain extract (1.7 mg/g) | extract | -- | |
| S1 | BE (0.30 mg/mL) | water | mono-component | + |
| S2 | BE (0.30 mg/mL) | glucose (13.7%) | bi-component | ++ |
| S3 | BE (0.30 mg/mL) | citric acid (0.03%) | bi-component | ++ |
| S4 | BE (0.30 mg/mL) | pectin (0.9%) | bi-component | ++ |
| S5 | BE (0.30 mg/mL) | mucilage (0.2%) | bi-component | ++ |
| S6 | G+M+P+CA | multi-component | +++ | |
| FJ | 0.30 mg/mL betalains | fresh red prickly pear fruit juice | complex | ++++ |
| BF | 0.30 mg/mL betalains | formulated beverage | complex | ++++ |
| BP | 0.30 mg/mg betalains | formulated beverage, pasteurized | complex | ++++ |
| Sample | Betacyanins (mg Eb/g) λmax | Betaxanthins (mg EI/g) λmax | Total Betalains (mg Eb+EI/g) | Total Betalains in the Sample |
|---|---|---|---|---|
| Betalain extract (150 g fresh juice) | 1.206 ± 0.003 | 1.018 ± 0.002 | 2.224 ± 0.005 | 33.36 ± 0.005 |
| Fresh juice (100 g) | 0.144 ± 0.002 | 0.120± 0.002 | 0.264 ± 0.005 | 26.4 ± 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yvonne, R.-M.; Socorro Josefina, V.-R.; Luis Alfonso, M.; Judith Esmeralda, U.-S. Impact of the Food Matrix on the Antioxidant and Hypoglycemic Effects of Betalains from Red Prickly Pear Juice After In Vitro Digestion. Foods 2025, 14, 1757. https://doi.org/10.3390/foods14101757
Yvonne R-M, Socorro Josefina V-R, Luis Alfonso M, Judith Esmeralda U-S. Impact of the Food Matrix on the Antioxidant and Hypoglycemic Effects of Betalains from Red Prickly Pear Juice After In Vitro Digestion. Foods. 2025; 14(10):1757. https://doi.org/10.3390/foods14101757
Chicago/Turabian StyleYvonne, Roman-Maldonado, Villanueva-Rodríguez Socorro Josefina, Mojica Luis Alfonso, and Urías-Silvas Judith Esmeralda. 2025. "Impact of the Food Matrix on the Antioxidant and Hypoglycemic Effects of Betalains from Red Prickly Pear Juice After In Vitro Digestion" Foods 14, no. 10: 1757. https://doi.org/10.3390/foods14101757
APA StyleYvonne, R.-M., Socorro Josefina, V.-R., Luis Alfonso, M., & Judith Esmeralda, U.-S. (2025). Impact of the Food Matrix on the Antioxidant and Hypoglycemic Effects of Betalains from Red Prickly Pear Juice After In Vitro Digestion. Foods, 14(10), 1757. https://doi.org/10.3390/foods14101757