
Investigation on the Efficacy of Two Food Supplements Containing a Fixed Combination of Selected Probiotics and β-Glucans or Elderberry Extract for the Immune System: Modulation on Cytokines Expression in Human THP-1 and PBMC

 ,
, 

Abstract
1. Introduction
2. Materials and Methods
2.1. Tested Samples and Probiotics Preparation
2.2. THP-1 and Peripheral Blood Mononuclear Cells (PBMC) Culture
2.3. Treatments
2.4. Cytokine Dosage
2.5. Statistical Analysis
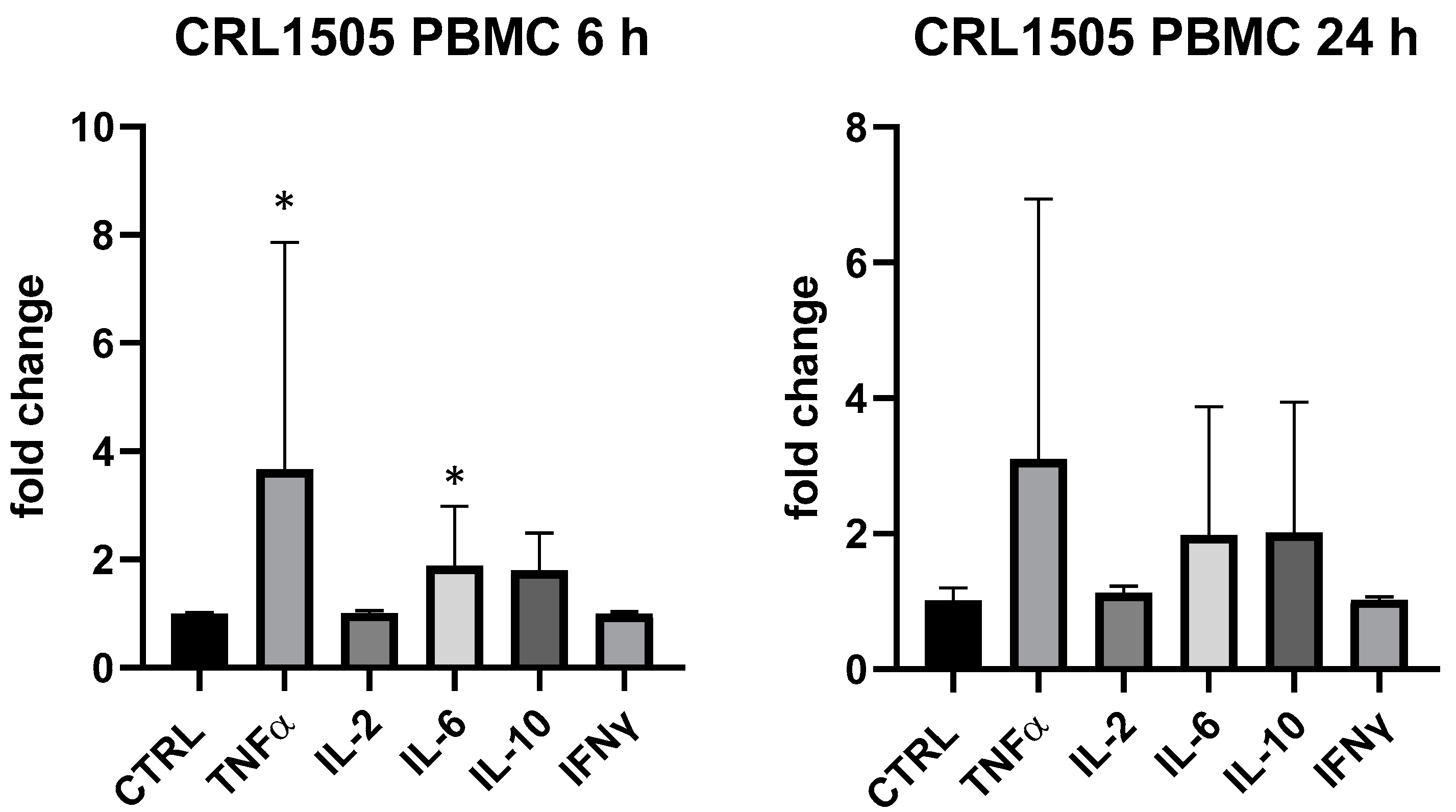
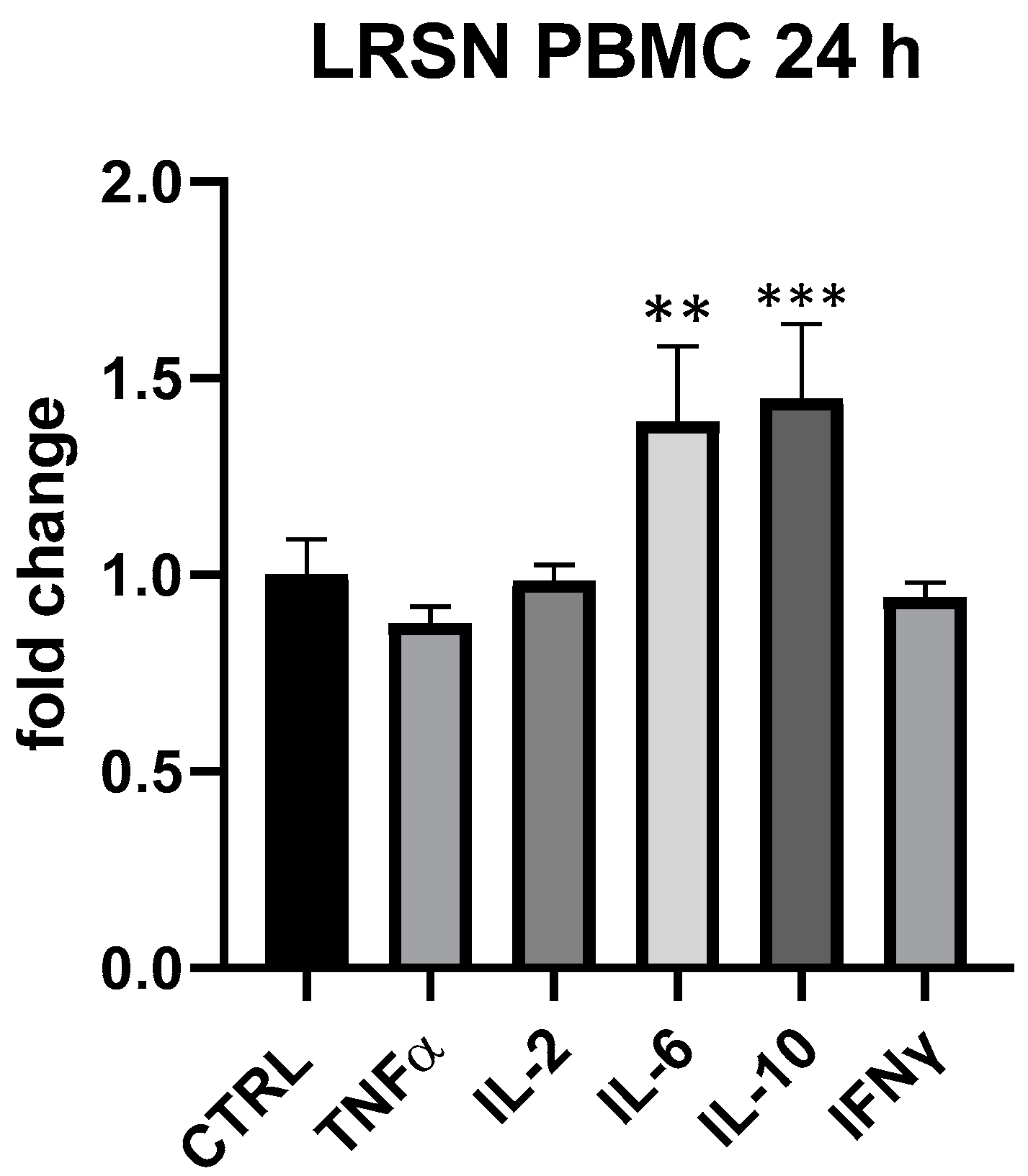
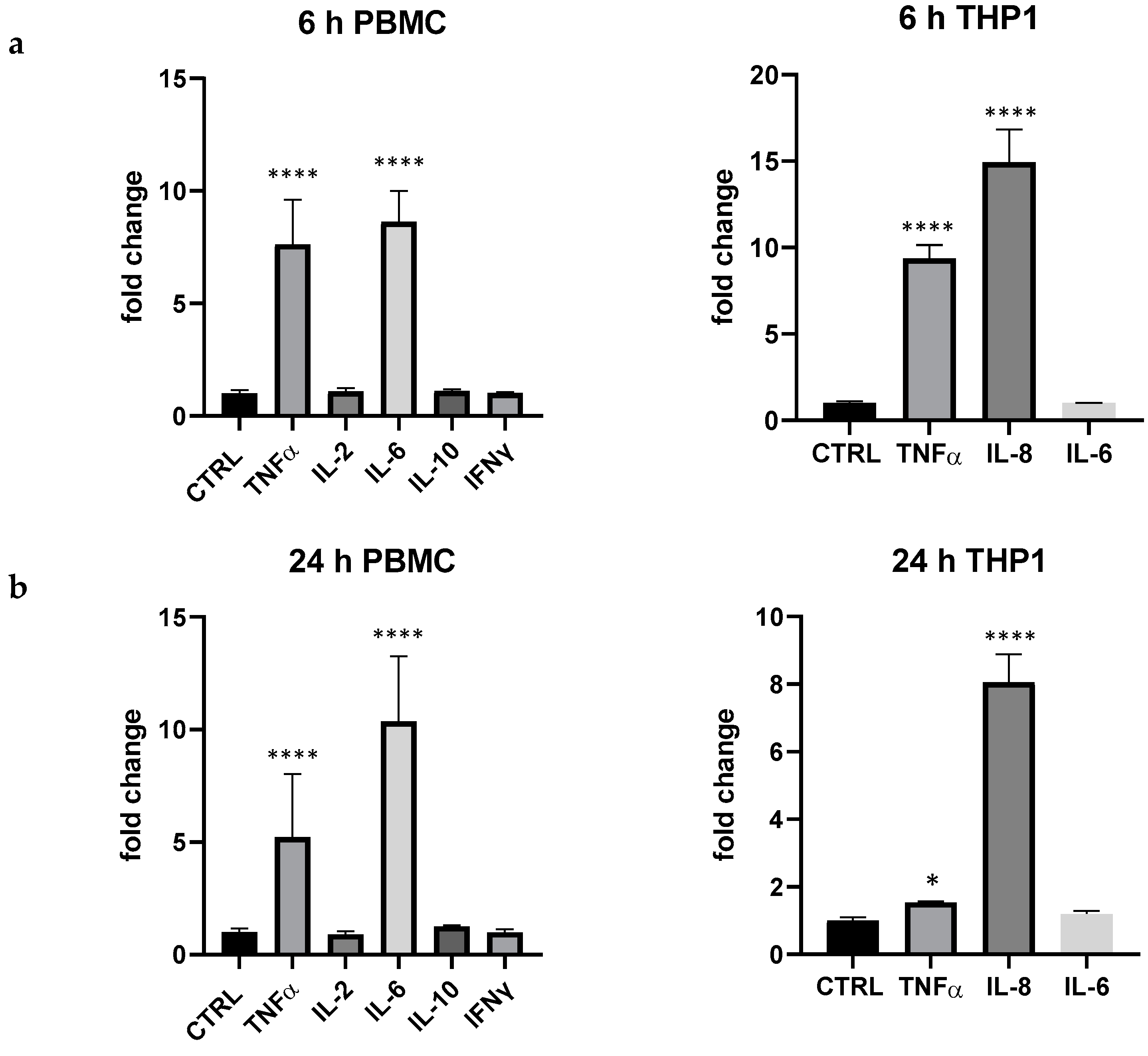
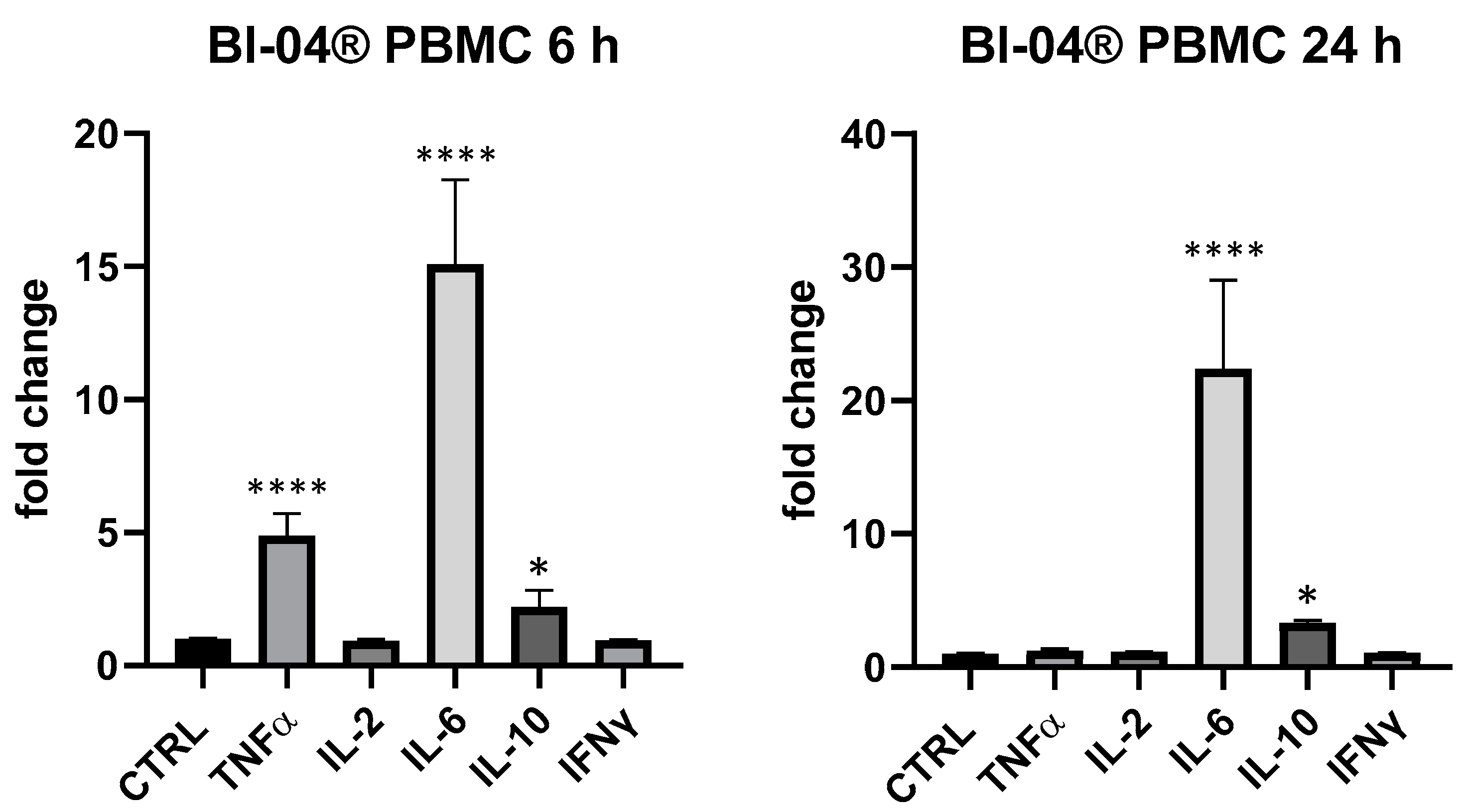
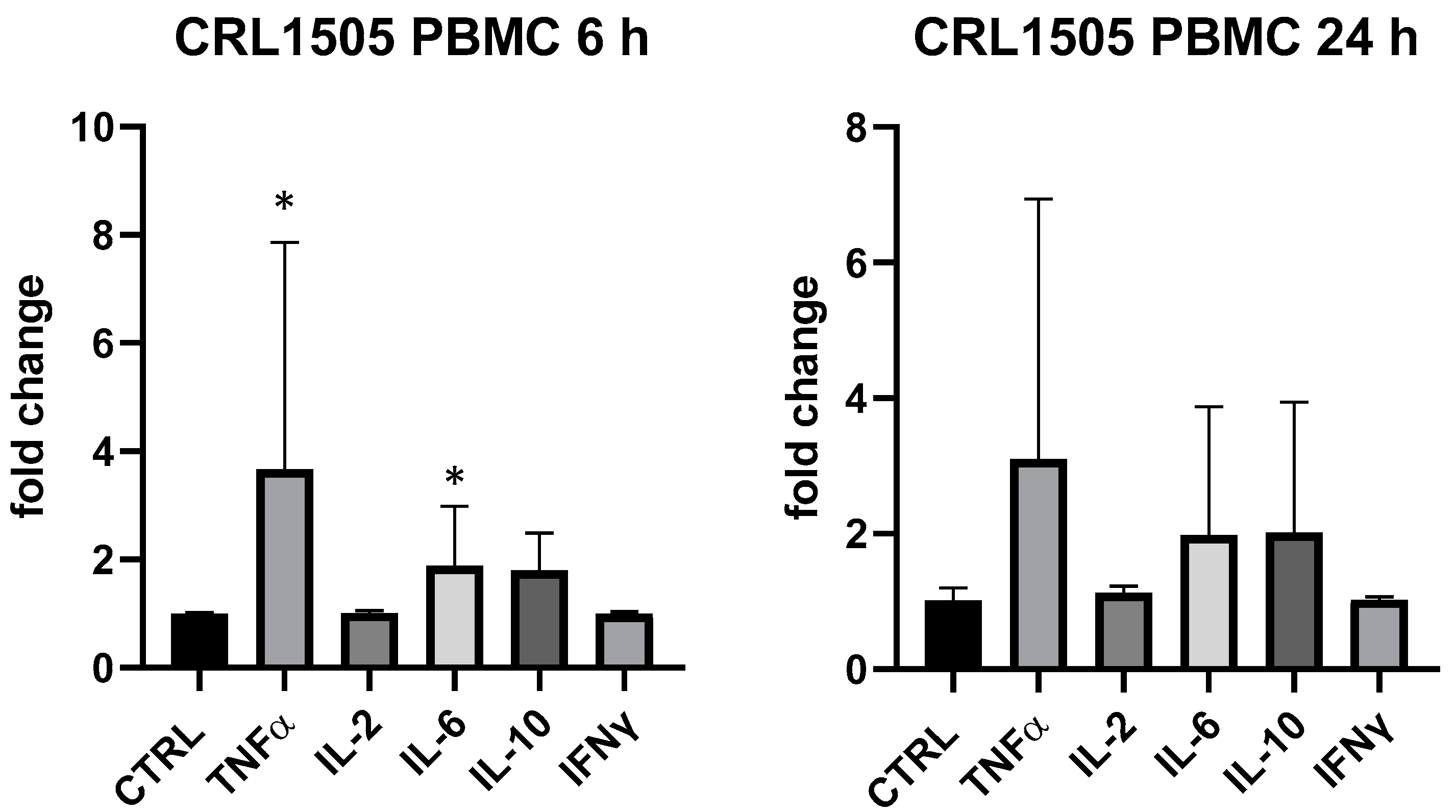
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Singh, M.R.; Yadav, K.; Chaurasiya, N.D.; Singh, D. Immune system and mechanism of immunomodulation. In Plants and Phytomolecules for Immunomodulation: Recent Trends and Advances; Springer Nature Singapore: Singapore, 2022; pp. 1–31. [Google Scholar]
- Antolín, C.S.; Valiante, M. Manuale CTO di Medicina e Chirurgia; Grupo CTO Editorial; Grupo CTO: Madrid, Spain, 2016. [Google Scholar]
- European Medicine Agency. Available online: www.ema.europa.eu (accessed on 27 December 2023).
- Trabattoni, D.; Clerici, M.; Centanni, S.; Mantero, M.; Garziano, M.; Blasi, F. Immunomodulatory effects of pidotimod in adults with community-acquired pneumonia undergoing standard antibiotic therapy. Pulm. Pharmacol. Ther. 2017, 44, 24–29. [Google Scholar] [CrossRef]
- DrugBank Database. Available online: www.drugbank.com (accessed on 27 December 2023).
- Biagi, M.; Pecorari, R.; Appendino, G.; Miraldi, E.; Magnano, A.R.; Governa, P.; Cettolin, G.; Giachetti, D. Herbal products in Italy: The thin line between phytotherapy, nutrition and parapharmaceuticals; a normative overview of the fastest growing market in Europe. Pharmaceuticals 2016, 9, 65. [Google Scholar] [CrossRef] [PubMed]
- Łagowska, K.; Bajerska, J. Probiotic Supplementation and Respiratory Infection and Immune Function in Athletes: Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Athl. Train. 2021, 56, 1213–1223. [Google Scholar] [CrossRef]
- Miller, L.E.; Lehtoranta, L.; Lehtinen, M.J. Short-term probiotic supplementation enhances cellular immune function in healthy elderly: Systematic review and meta-analysis of controlled studies. Nutr. Res. 2019, 64, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Vlasova, A.N.; Kandasamy, S.; Chattha, K.S.; Rajashekara, G.; Saif, L.J. Comparison of probiotic lactobacilli and bifidobacteria effects, immune responses and rotavirus vaccines and infection in different host species. Vet. Immunol. Immunopathol. 2016, 172, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Spigoni, V.; Aldigeri, R.; Antonini, M.; Micheli, M.M.; Fantuzzi, F.; Fratter, A.; Pellizzato, M.; Derlindati, E.; Zavaroni, I.; Bonadonna, R.C.; et al. Effects of a New Nutraceutical Formulation (Berberine, Red Yeast Rice and Chitosan) on Non-HDL Cholesterol Levels in Individuals with Dyslipidemia: Results from a Randomized, Double Blind, Placebo-Controlled Study. Int. J. Mol. Sci. 2017, 18, 1498. [Google Scholar] [CrossRef] [PubMed]
- Fratter, A.; Mason, V.; Pellizzato, M.; Valier, S.; Cicero, A.F.G.; Tedesco, E.; Meneghetti, E.; Benetti, F. Lipomatrix: A Novel Ascorbyl Palmitate-Based Lipid Matrix to Enhancing Enteric Absorption of Serenoa Repens Oil. Int. J. Mol. Sci. 2019, 20, 669. [Google Scholar] [CrossRef]
- Borgonetti, V.; Benatti, C.; Governa, P.; Isoldi, G.; Pellati, F.; Alboni, S.; Tascedda, F.; Montopoli, M.; Galeotti, N.; Manetti, F.; et al. Non-psychotropic Cannabis sativa L. phytocomplex modulates microglial inflammatory response through CB2 receptors-, endocannabinoids-, and NF-κB-mediated signaling. Phytother. Res. 2022, 36, 2246–2263. [Google Scholar] [CrossRef]
- Turner, R.B.; Woodfolk, J.A.; Borish, L.; Steinke, J.W.; Patrie, J.T.; Muehling, L.M.; Lahtinen, S.; Lehtinen, M.J. Effect of probiotic on innate inflammatory response and viral shedding in experimental rhinovirus infection—A randomised controlled trial. Benef. Microbes 2017, 8, 207–215. [Google Scholar] [CrossRef]
- Dentice Maidana, S.; Imamura, Y.; Elean, M.; Albarracín, L.; Nishiyama, K.; Suda, Y.; Kurata, S.; Jure, M.Á.; Kitazawa, H.; Villena, J. Oral Administration of Lacticaseibacillus rhamnosus CRL1505 Modulates Lung Innate Immune Response against Klebsiella pneumoniae ST25. Microorganisms 2023, 11, 1148. [Google Scholar] [CrossRef]
- Italian Ministry of Health: Ministerial Decree 2021/08/10. Attachment 1. Available online: http://www.trovanorme.salute.gov.it/norme/renderNormsanPdf?anno=2019&codLeg=70165&parte=2&serie= (accessed on 27 December 2023).
- Raso, F.; Bulferi, G.; De Togni, H.; Pauletto, L.; Carnevali, I. Pharmaceutical Formulations for Oral Use Containing Probiotic Strains for Modulation of the Immune Function. Patent No. 102021000015077 (Italy); Patent Pending No. WO2022259135A1; EP22734043.7, 21 June 2021. [Google Scholar]
- Governa, P.; Romagnoli, G.; Albanese, P.; Rossi, F.; Manetti, F.; Biagi, M. Effect of in vitro simulated digestion on the anti-Helicobacter Pylori activity of different Propolis extracts. J. Enzym. Inhib. Med. Chem. 2023, 38, 2183810. [Google Scholar] [CrossRef]
- Pressi, G.; Rigillo, G.; Governa, P.; Borgonetti, V.; Baini, G.; Rizzi, R.; Guarnerio, C.; Bertaiola, O.; Frigo, M.; Merlin, M.; et al. A Novel Perilla frutescens (L.) Britton Cell-Derived Phytocomplex Regulates Keratinocytes Inflammatory Cascade and Barrier Function and Preserves Vaginal Mucosal Integrity In Vivo. Pharmaceutics 2023, 15, 240. [Google Scholar] [CrossRef]
- Governa, P.; Marchi, M.; Cocetta, V.; De Leo, B.; Saunders, P.T.; Catanzaro, D.; Miraldi, E.; Montopoli, M.; Biagi, M. Effects of Boswellia Serrata Roxb. and Curcuma longa L. in an in vitro intestinal inflammation model using immune cells and Caco-2. Pharmaceuticals 2018, 11, 126. [Google Scholar] [CrossRef] [PubMed]
- Governa, P.; Cusi, M.G.; Borgonetti, V.; Sforcin, J.M.; Terrosi, C.; Baini, G.; Miraldi, E.; Biagi, M. Beyond the biological effect of a chemically characterized poplar propolis: Antibacterial and antiviral activity and comparison with flurbiprofen in cytokines release by LPS-stimulated human mononuclear cells. Biomedicines 2019, 7, 73. [Google Scholar] [CrossRef] [PubMed]
- Gertsch, J. Botanical Drugs, Synergy, and Network Pharmacology: Forth and Back to Intelligent Mixtures. Planta Medica 2011, 77, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Segura, M.; Vadeboncoeur, N.; Gottschalk, M. CD14-dependent and-independent cytokine and chemokine production by human THP-1 monocytes stimulated by Streptococcus suis capsular type 2. Clin. Exp. Immunol. 2002, 127, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Schindler, R.; Mancilla, J.; Endres, S.; Ghorbani, R.; Clark, S.C.; Dinarello, C.A. Correlations and interactions in the production of interleukin-6 (IL-6), IL-1, and tumor necrosis factor (TNF) in human blood mononuclear cells: IL-6 suppresses IL-1 and TNF. Blood 1990, 75, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Janský, L.; Reymanová, P.; Kopecký, J. Dynamics of cytokine production in human peripheral blood mononuclear cells stimulated by LPS or infected by Borrelia. Physiol. Res. 2003, 52, 593–598. [Google Scholar] [CrossRef] [PubMed]
- Nastasi, C.; Mannarino, L.; D’Incalci, M. DNA Damage Response and Immune Defense. Int. J. Mol. Sci. 2020, 21, 7504. [Google Scholar] [CrossRef] [PubMed]
- Al Mahmud, A.; Shafayet, A.S.; Karim, M.R.; Al-Mamun, M.R.; Akhter, S.; Sohel, M.; Hasan, M.; Bellah, S.F.; Amin, M.N. Clinically proven natural products, vitamins and mineral in boosting up immunity: A comprehensive review. Heliyon 2023, 9, e15292. [Google Scholar] [CrossRef]
- Yoo, J.Y.; Groer, M.; Dutra, S.V.O.; Sarkar, A.; McSkimming, D.I. Gut Microbiota and Immune System Interactions. Microorganisms 2020, 8, 1587. [Google Scholar] [CrossRef] [PubMed]
- Gasmi, A.; Shanaida, M.; Oleshchuk, O.; Semenova, Y.; Mujawdiya, P.K.; Ivankiv, Y.; Pokryshko, O.; Noor, S.; Piscopo, S.; Adamiv, S.; et al. Natural Ingredients to Improve Immunity. Pharmaceuticals 2023, 16, 528. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, A.L.; Murai, I.H.; Reis, B.Z.; Sales, L.P.; Santos, M.D.; Pinto, A.J.; Goessler, K.F.; Duran, C.S.C.; Silva, C.B.R.; Franco, A.S.; et al. Effect of a single high dose of vitamin D3 on cytokines, chemokines, and growth factor in patients with moderate to severe COVID-19. Am. J. Clin. Nutr. 2022, 115, 790–798. [Google Scholar] [CrossRef]
- Ramírez-Pérez, S.; Hernández-Palma, L.A.; Oregon-Romero, E.; Anaya-Macías, B.U.; García-Arellano, S.; González-Estevez, G.; Muñoz-Valle, J.F. Downregulation of inflammatory cytokine release from IL-1β and LPS-stimulated PBMC orchestrated by ST2825, a MyD88 dimerisation inhibitor. Molecules 2020, 25, 4322. [Google Scholar] [CrossRef] [PubMed]
- Mahashur, A.; Thomas, P.K.; Mehta, P.; Nivangune, K.; Muchhala, S.; Jain, R. Pidotimod: In-depth review of current evidence. Lung India 2019, 36, 422–433. [Google Scholar] [CrossRef]
- Di Renzo, M.; Pasqui, A.L.; Bruni, F.; Saletti, M.; Bova, G.; Chiarion, C.; Girardello, R.; Ferrì, P.; Auteri, A. The in vitro effect of Pidotimod on some immune functions in cancer patients. Immunopharmacol. Immunotoxicol. 1997, 19, 37–51. [Google Scholar] [CrossRef]
- Mazziotta, C.; Tognon, M.; Martini, F.; Torreggiani, E.; Rotondo, J.C. Probiotics Mechanism of Action on Immune Cells and Beneficial Effects on Human Health. Cells 2023, 12, 184. [Google Scholar] [CrossRef]
- Plaza-Diaz, J.; Ruiz-Ojeda, F.J.; Gil-Campos, M.; Gil, A. Mechanisms of Action of Probiotics. Adv. Nutr. 2019, 10 (Suppl. 1), S49–S66. [Google Scholar] [CrossRef]
- López, P.; Gueimonde, M.; Margolles, A.; Suárez, A. Distinct Bifidobacterium strains drive different immune responses in vitro. Int. J. Food Microbiol. 2010, 138, 157–165. [Google Scholar] [CrossRef]
- Fong, F.L.Y.; Kirjavainen, P.V.; El-Nezami, H. Immunomodulation of Lactobacillus rhamnosus GG (LGG)-derived soluble factors on antigen-presenting cells of healthy blood donors. Sci. Rep. 2016, 6, 22845. [Google Scholar] [CrossRef]
- Villena, J.; Chiba, E.; Vizoso-Pinto, M.G.; Tomosada, Y.; Takahashi, T.; Ishizuka, T.; Aso, H.; Salva, S.; Alvarez, S.; Kitazawa, H. Immunobiotic Lactobacillus rhamnosus strains differentially modulate antiviral immune response in porcine intestinal epithelial and antigen presenting cells. BMC Microbiol. 2014, 14, 126. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, E.; Brzezińska-Błaszczyk, E.; Rasmus, P.; Żelechowska, P. Fungal β-glucans and mannan stimulate peripheral blood mononuclear cells to cytokine production in Syk-dependent manner. Immunobiology 2020, 225, 151985. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, F.; Weng, D.; Song, L.; Li, C.; Tang, W.; Yu, Y.; Dai, W.; Chen, J. T(reg) cells may regulate interlukin-17 production by modulating TH1 responses in 1,3-β-glucan-induced lung inflammation in mice. J. Immunotoxicol. 2013, 10, 253–261. [Google Scholar] [CrossRef] [PubMed]
- EMA Assessment Report on Sambucus nigra, L. Flower. Available online: https://www.ema.europa.eu/en/documents/herbal-report/final-assessment-report-sambucus-nigra-l-flos-revision-1_en.pdf (accessed on 27 December 2023).
- Stich, L.; Plattner, S.; McDougall, G.; Austin, C.; Steinkasserer, A. Polysaccharides from European Black Elderberry Extract Enhance Dendritic Cell Mediated T Cell Immune Responses. Int. J. Mol. Sci. 2022, 23, 3949. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulation 1, Adults (BLBG) | Formulation 2, Children (LRSN) |
|---|---|
|
|
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cappellucci, G.; Baini, G.; Miraldi, E.; Pauletto, L.; De Togni, H.; Raso, F.; Biagi, M. Investigation on the Efficacy of Two Food Supplements Containing a Fixed Combination of Selected Probiotics and β-Glucans or Elderberry Extract for the Immune System: Modulation on Cytokines Expression in Human THP-1 and PBMC. Foods 2024, 13, 458. https://doi.org/10.3390/foods13030458
Cappellucci G, Baini G, Miraldi E, Pauletto L, De Togni H, Raso F, Biagi M. Investigation on the Efficacy of Two Food Supplements Containing a Fixed Combination of Selected Probiotics and β-Glucans or Elderberry Extract for the Immune System: Modulation on Cytokines Expression in Human THP-1 and PBMC. Foods. 2024; 13(3):458. https://doi.org/10.3390/foods13030458
Chicago/Turabian StyleCappellucci, Giorgio, Giulia Baini, Elisabetta Miraldi, Lara Pauletto, Heide De Togni, Floriana Raso, and Marco Biagi. 2024. "Investigation on the Efficacy of Two Food Supplements Containing a Fixed Combination of Selected Probiotics and β-Glucans or Elderberry Extract for the Immune System: Modulation on Cytokines Expression in Human THP-1 and PBMC" Foods 13, no. 3: 458. https://doi.org/10.3390/foods13030458
APA StyleCappellucci, G., Baini, G., Miraldi, E., Pauletto, L., De Togni, H., Raso, F., & Biagi, M. (2024). Investigation on the Efficacy of Two Food Supplements Containing a Fixed Combination of Selected Probiotics and β-Glucans or Elderberry Extract for the Immune System: Modulation on Cytokines Expression in Human THP-1 and PBMC. Foods, 13(3), 458. https://doi.org/10.3390/foods13030458