
Integrative Metabolomic and Transcriptomic Analysis Provides Novel Insights into the Effects of SO2 on the Postharvest Quality of ‘Munage’ Table Grapes



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Samples
2.2. Determination of Physicochemical Indices of Grapes
2.3. Transcriptome Sequencing and Analysis
2.4. Metabolite Extraction and Analysis
2.5. Association Analysis of the Metabolome and Transcriptome
2.6. qRT-PCR Analysis
2.7. Statistical Analysis
3. Results
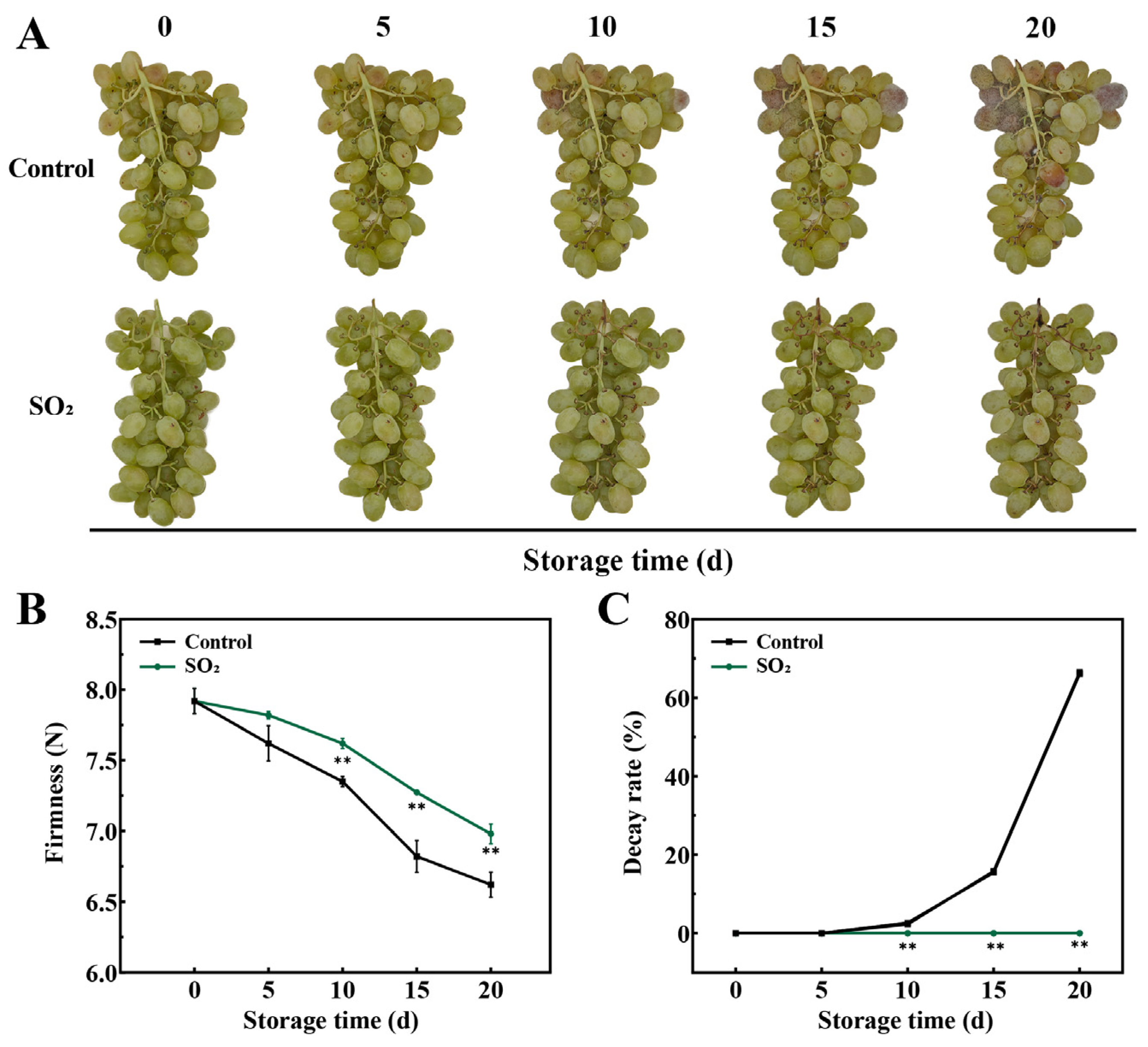
3.1. Effect of SO2 Treatment on the Quality Indices in Grapes
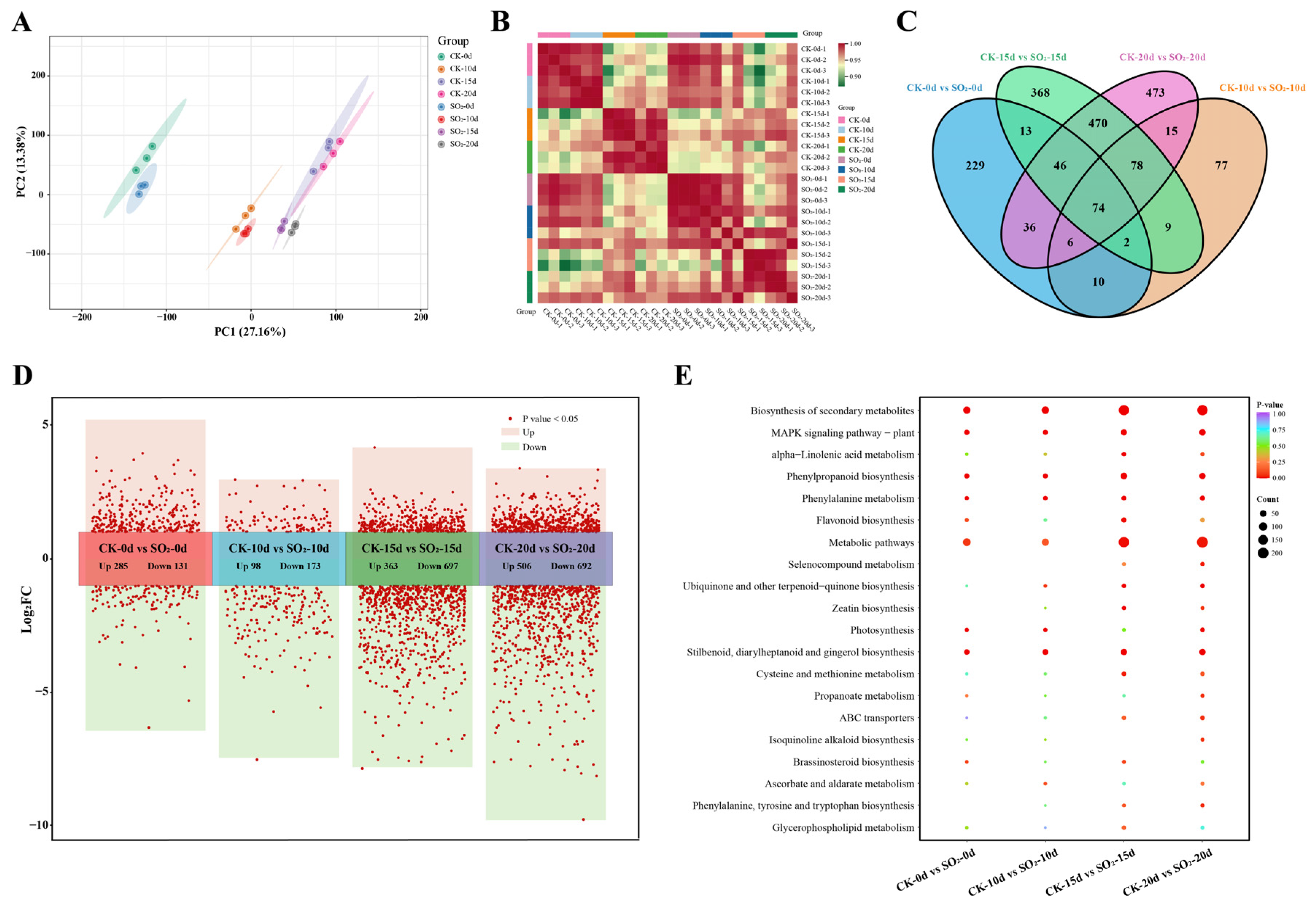
3.2. Transcriptomic Analysis
3.2.1. General Description of Transcriptomic Data
3.2.2. Analysis of DEGs in Grapes Under SO2 Treatment
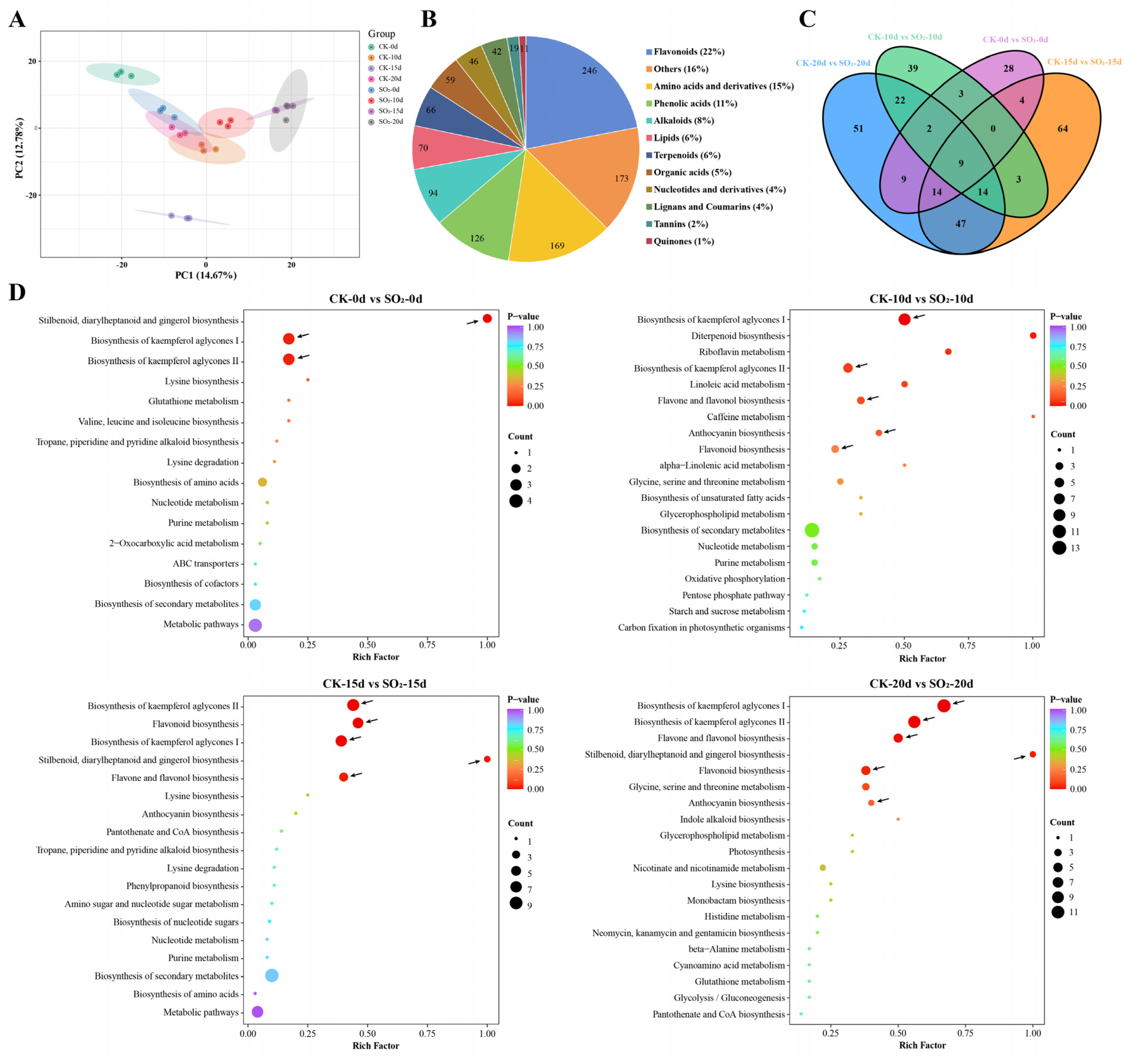
3.3. Metabolomic Analysis
3.3.1. Analysis of DAMs in Grape Under SO2 Treatment
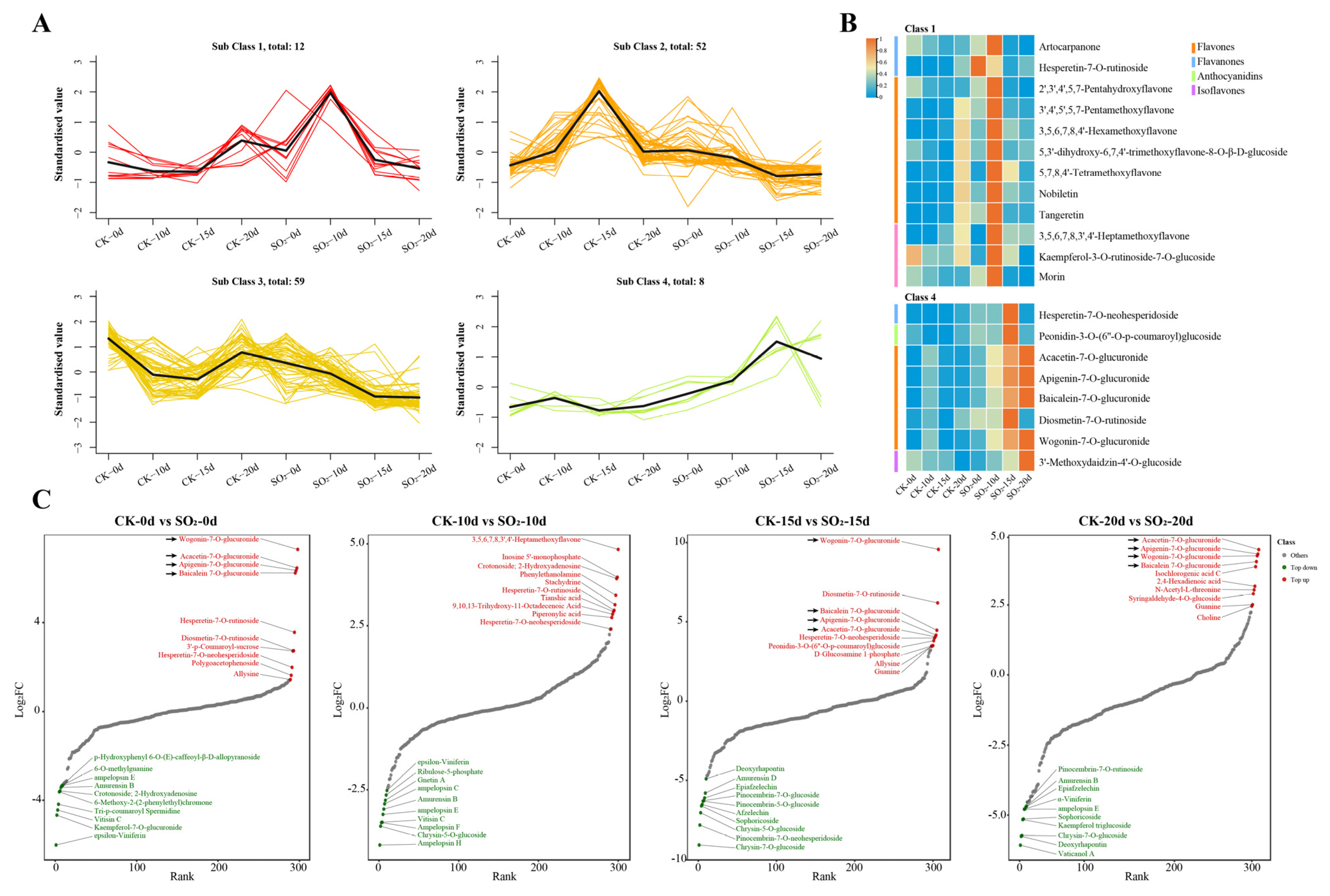
3.3.2. Analysis of Key Metabolites in Grape Under SO2 Treatment
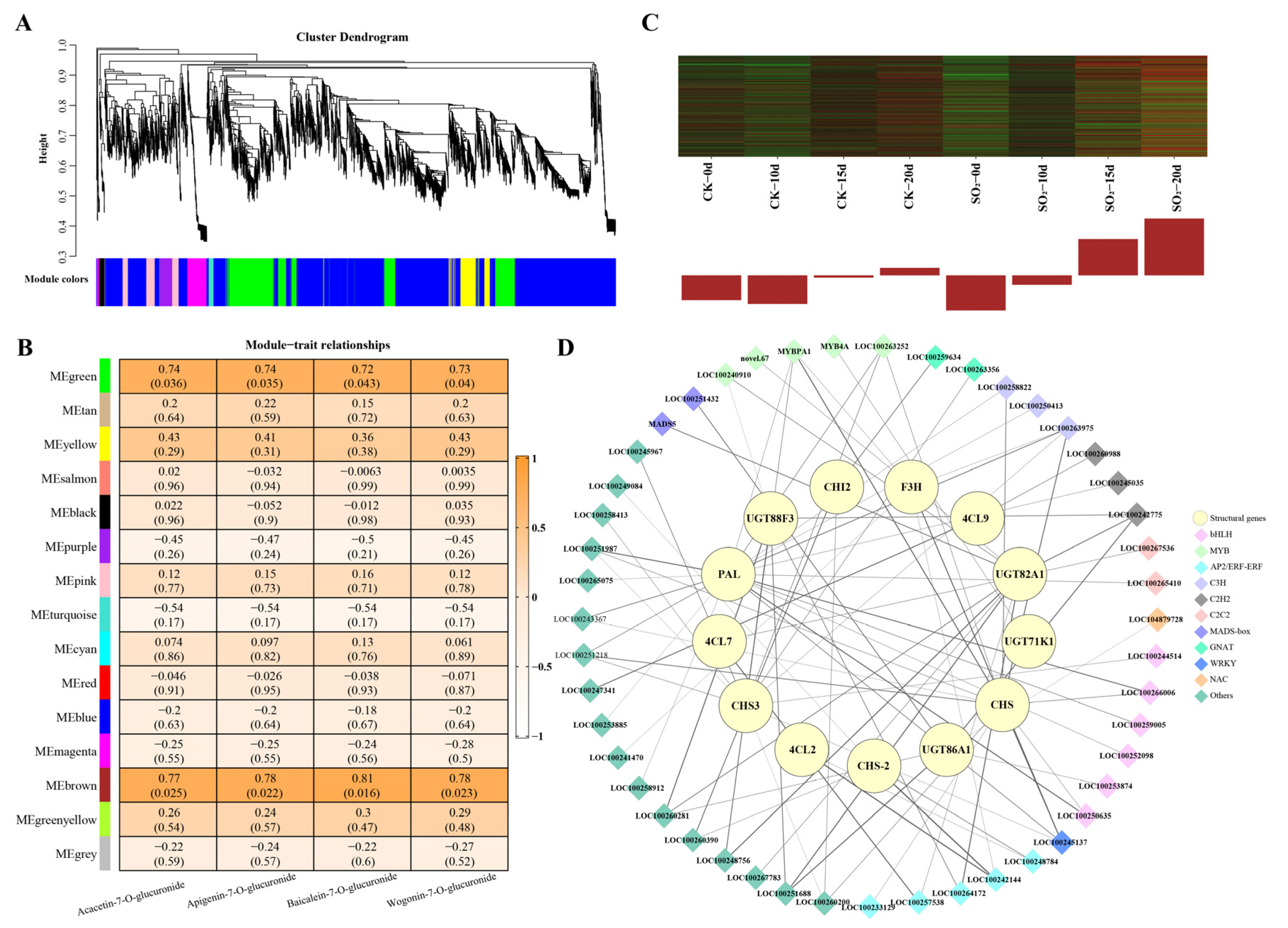
3.4. Co-Expression Network Analysis Associated with Key Flavonoids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ju, Y.L.; Wang, W.N.; Yue, X.F.; Xue, W.; Zhang, Y.L.; Fang, Y.L. Integrated metabolomic and transcriptomic analysis reveals the mechanism underlying the accumulation of anthocyanins and other flavonoids in the flesh and skin of teinturier grapes. Plant Physiol. Biochem. 2023, 197, 107667. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xing, S.J.; Yuan, Y.Y.; Zheng, Y.G.; Tian, Q.M.; Wu, B.; Wei, J. Sulfur dioxide fumigation promotes GABA accumulation and energy metabolism to delay quality deterioration in postharvest table grapes. Postharvest Biol. Technol. 2024, 215, 113022. [Google Scholar] [CrossRef]
- Zhang, K.K.; Li, W.P.; Ju, Y.L.; Wang, X.H.; Sun, X.Y.; Fang, Y.L.; Chen, K.Q. Transcriptomic and metabolomic basis of short- and long-term post-harvest UV-C application in regulating grape berry quality development. Foods 2021, 10, 625. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.B.; Zhao, P.C.; Zhang, P.A.; Su, L.Y.; Jia, H.R.; Wei, X.K.; Fang, J.G.; Jia, H.F. Integrative transcriptomics and metabolomics data exploring the effect of chitosan on postharvest grape resistance to Botrytis cinerea. Postharvest Biol. Technol. 2020, 167, 111248. [Google Scholar] [CrossRef]
- Wu, Z.H.; Dong, C.H.; Wei, J.; Guo, L.M.; Meng, Y.N.; Wu, B.; Chen, J.L. A transcriptional study of the effects of nitric oxide on rachis browning in table grapes cv. Thompson seedless. Postharvest Biol. Technol. 2021, 175, 111471. [Google Scholar] [CrossRef]
- Ma, W.Y.; Xu, L.L.; Gao, S.W.; Lyu, X.; Cao, X.L.; Yao, Y.X. Melatonin alters the secondary metabolite profile of grape berry skin by promoting VvMYB14-mediated ethylene biosynthesis. Hortic. Res. 2021, 8, 43. [Google Scholar] [CrossRef]
- Saito, S.; Obenland, D.; Xiao, C.L. Influence of sulfur dioxide-emitting polyethylene packaging on blueberry decay and quality during extended storage. Postharvest Biol. Technol. 2020, 160, 111045. [Google Scholar] [CrossRef]
- Yang, Y.T.; Gong, Y.; Gao, Y.F.; Huang, J.Y.; Xu, M.; Xiong, B. Study on the preservation effect of propolis on sweet cherry. Earth Environ. Sci. 2020, 474, 032029. [Google Scholar] [CrossRef]
- Ali, Q.; Dogan, A.; Erkan, M. Sulfur dioxide generating pads containing different concentrations of sodium metabisulfite maintains postharvest quality of ‘hicaznar’ pomegranate. Sci. Hortic. 2024, 333, 113249. [Google Scholar] [CrossRef]
- Xue, M.Z.; Yi, H.L. Induction of disease resistance providing new insight into sulfur dioxide preservation in Vitis vinifera L. Sci. Hortic. 2017, 225, 567–573. [Google Scholar] [CrossRef]
- Zhang, J.; Xie, L.J.; Wang, H.J.; Zhou, S.H.; Zhu, Z.Q.; Xie, T.L.; Zhou, Y.M.; Li, W.; Pang, L.T.; Sun, J.; et al. Metabolome and transcriptome analyses provide insight into the effect of 1-MCP and SO2 preservatives on the synthesis and regulation of phenols in ‘Shine Muscat’ storage grapes. LWT 2024, 203, 116400. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, Z.H.; Yuan, Y.Y.; Zhang, J.X.; Wei, J.; Wu, B. Sulfur dioxide mitigates oxidative damage by modulating hydrogen peroxide homeostasis in postharvest table grapes. Postharvest Biol. Technol. 2022, 188, 111877. [Google Scholar] [CrossRef]
- Yan, D.M.; Yi, H.L. Transcriptome analysis provides insights into preservation mechanism of postharvest muscat hamburg grapes treated with SO2. Sci. Hortic. 2024, 331, 113108. [Google Scholar] [CrossRef]
- Li, Z.B.; Chen, S.Q.; Qi, M.; Yang, M.Y.; Yuan, H.M.; Xu, Y.Q.; Huang, J.; Li, D.; Zhou, W.; Yuan, Y.Y.; et al. Inhibition of postharvest rachis browning of table grapes by sulfur dioxide: Evidence from phenolic metabolism and sulfur assimilation. Postharvest Biol. Technol. 2023, 204, 112413. [Google Scholar] [CrossRef]
- Xue, M.Z.; Yi, H.L.; Wang, H. Identification of miRNAs involved in SO2 preservation in Vitis vinifera L. by deep sequencing. Environ. Exp. Bot. 2018, 153, 218–228. [Google Scholar] [CrossRef]
- Zhang, Z.; Wei, J.; Wang, M.; Zhang, J.; Wu, B. Induced sulfur metabolism by sulfur dioxide maintains postharvest quality of ‘thompson seedless’ grape through increasing sulfite content. J. Sci. Food Agric. 2022, 102, 1174–1184. [Google Scholar] [CrossRef]
- Zhang, Z.; Xu, J.; Chen, Y.; Wei, J.; Wu, B. Nitric oxide treatment maintains postharvest quality of table grapes by mitigation of oxidative damage. Postharvest Biol. Technol. 2019, 152, 9–18. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef]
- Yang, Z.C.; Lin, M.H.; Yang, X.Z.; Wu, D.; Chen, K.S. Comprehensive analysis of transcriptome and metabolome provides insights into the stress response mechanisms of apple fruit to postharvest impact damage. Food Chem. Mol. Sci. 2023, 7, 100176. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-Seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Mou, Z.L.; Wang, L.; Zeng, Z.X.; Su, X.G.; Ji, S.J.; Shan, W.; Kuang, J.F.; Lu, W.J.; Chen, Y.L.; Zhao, Y.T.; et al. Metabolomics integrated with transcriptomics unveil the regulatory pathways of modified atmosphere packaging–maintained leaf quality of chinese flowering cabbage. Food Chem. 2023, 405, 134910. [Google Scholar] [CrossRef]
- Song, M.Y.; Wang, L.; Zhang, Y.T.; Wang, Q.G.; Han, X.; Yang, Q.; Zhang, J.H.; Tong, Z.K. Temporospatial pattern of flavonoid metabolites and potential regulatory pathway of PbMYB211-coordinated kaempferol-3-O-rhamnoside biosynthesis in Phoebe bournei. Plant Physiol. Biochem. 2023, 202, 107913. [Google Scholar] [CrossRef] [PubMed]
- Guven, H.; Arici, A.; Simsek, O. Flavonoids in our foods: A short review. J. Basic Clin. Health Sci. 2019, 3, 96–106. [Google Scholar] [CrossRef]
- Han, S.A.; Xie, H.; Wang, M.; Zhang, J.G.; Xu, Y.H.; Zhu, X.H.; Caikasimu, A.; Zhou, X.W.; Mai, S.L.; Pan, M.Q.; et al. Transcriptome and metabolome reveal the effects of three canopy types on the flavonoids and phenolic acids in ‘Merlot’ (Vitis vinifera L.) berry pericarp. Food Res. Int. 2023, 163, 112196. [Google Scholar] [CrossRef]
- Zhao, Q.; Zhang, Y.; Wang, G.; Hill, L.; Weng, J.K.; Chen, X.Y.; Xue, H.X.; Martin, C. A specialized flavone biosynthetic pathway has evolved in the medicinal plant, Scutellaria baicalensis. Sci. Adv. 2016, 2, e1501780. [Google Scholar] [CrossRef]
- Zhao, Q.; Yang, J.; Cui, M.Y.; Liu, J.; Fang, Y.M.; Yan, M.X.; Qiu, W.Q.; Shang, H.W.; Xu, Z.C.; Yidiresi, R.; et al. The reference genome sequence of Scutellaria baicalensis provides insights into the evolution of wogonin biosynthesis. Mol. Plant. 2019, 12, 935–950. [Google Scholar] [CrossRef]
- Shojaeifard, Z.; Hemmateenejad, B.; Jassbi, A.R. Chemometrics-based LC-UV-ESIMS analyses of 50 Salvia species for detecting their antioxidant constituents. J. Pharm. Biomed. Anal. 2021, 193, 113745. [Google Scholar] [CrossRef]
- Donnez, D.; Jeandet, P.; Clément, C.; Courot, E. Bioproduction of resveratrol and stilbene derivatives by plant cells and microorganisms. Trends Biotechnol. 2009, 27, 706–713. [Google Scholar] [CrossRef]
- Rivière, C.; Pawlus, A.D.; Mérillon, J. Natural stilbenoids: Distribution in the plant kingdom and chemotaxonomic interest in vitaceae. Nat. Prod. Rep. 2012, 29, 1317–1333. [Google Scholar] [CrossRef]
- Xu, R.R.; Wang, L.M.; Li, K.X.; Cao, J.K.; Zhao, Z.L. Integrative transcriptomic and metabolomic alterations unravel the effect of melatonin on mitigating postharvest chilling injury upon plum (cv. Friar) fruit. Postharvest Biol. Technol. 2022, 186, 111819. [Google Scholar] [CrossRef]
- Wang, Y.Q.; Zhang, Y.X.; Guo, Y.Y.; Ji, N.N.; Chen, Y.; Sun, Y.P.; Wang, Z.L.; Guan, L.X.; Guo, P.C. Integrated transcriptomic and metabolomic analysis reveals the effects and potential mechanism of hydrogen peroxide on pigment metabolism in postharvest broccoli. J. Food Sci. 2024, 89, 6189–6202. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Horvath, S. A general framework for weighted gene co-expression network analysis. Stat. Appl. Genet. Mol. Biol. 2005, 4, Article17. [Google Scholar] [CrossRef] [PubMed]
- Saito, K.; Yonekura-Sakakibara, K.; Nakabayashi, R.; Higashi, Y.; Yamazaki, M.; Tohge, T.; Fernie, A.R. The flavonoid biosynthetic pathway in arabidopsis: Structural and genetic diversity. Plant Physiol. Biochem. 2013, 72, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Andersen, J.R.; Zein, I.; Wenzel, G.; Darnhofer, B.; Eder, J.; Ouzunova, M.; Lübberstedt, T. Characterization of phenylpropanoid pathway genes within european maize (Zea mays L.) inbreds. BMC Plant Biol. 2008, 8, 2. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Hidalgo-Shrestha, C.; Bonawitz, N.D.; Franke, R.B.; Chapple, C. Spatio-temporal control of phenylpropanoid biosynthesis by inducible complementation of a cinnamate 4-hydroxylase mutant. J. Exp. Bot. 2021, 72, 3061–3073. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The origin and evolution of plant flavonoid metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef]
- Cheng, L.; Han, M.; Yang, L.M.; Yang, L.; Sun, Z.; Zhang, T. Changes in the physiological characteristics and baicalin biosynthesis metabolism of Scutellaria baicalensis Georgi under drought stress. Ind. Crop. Prod. 2018, 122, 473–482. [Google Scholar] [CrossRef]
- Jiang, Y.F.Y.; Ji, X.Y.; Duan, L.X.; Ye, P.; Yang, J.F.; Zhan, R.T.; Chen, W.W.; Ma, D.M. Gene mining and identification of a flavone synthase II involved in flavones biosynthesis by transcriptomic analysis and targeted flavonoid profiling in Chrysanthemum indicum L. Ind. Crop. Prod. 2019, 134, 244–256. [Google Scholar] [CrossRef]
- Wu, J.; Wang, X.C.; Liu, Y.; Du, H.; Shu, Q.Y.; Su, S.; Wang, L.J.; Li, S.S.; Wang, L.S. Flavone synthases from Lonicera japonica and L. macranthoides reveal differential flavone accumulation. Sci. Rep. 2016, 6, 19245. [Google Scholar] [CrossRef]
- Zhang, S.; Yang, J.; Li, H.Q.; Chiang, V.L.; Fu, Y.J. Cooperative regulation of flavonoid and lignin biosynthesis in plants. CRC Crit. Rev. Plant Sci. 2021, 40, 109–126. [Google Scholar] [CrossRef]
- Deluc, L.; Barrieu, F.; Marchive, C.; Lauvergeat, V.; Decendit, A.; Richard, T.; Carde, J.P.; Mérillon, J.M.; Hamdi, S. Characterization of a grapevine R2R3-MYB transcription factor that regulates the phenylpropanoid pathway. Plant Physiol. 2006, 140, 499–511. [Google Scholar] [CrossRef] [PubMed]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verriès, C.; Cheynier, V.; Romieu, C. Ectopic expression of VvMybPA2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol. 2009, 149, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Song, W.; Lin, S.Q.; Yin, Q.; Liu, T.H.; Gan, L.Z.; Qi, J.J.; Yang, Y.Y.; Wei, W.; Shan, W.; Kuang, J.F.; et al. A multi-omics approach reveals low temperature inhibition of flavones and flavonols accumulation in postharvest bananas via downregulation of MabHLH363. Postharvest Biol. Technol. 2024, 218, 113152. [Google Scholar] [CrossRef]
- Xu, W.J.; Dubos, C.; Lepiniec, L. Transcriptional control of flavonoid biosynthesis by MYB–bHLH–WDR complexes. Trends Plant Sci. 2015, 20, 176–185. [Google Scholar] [CrossRef]
- Wan, H.L.; Liu, Y.H.; Wang, T.T.; Jiang, P.; Wen, W.W.; Nie, J.Y. Combined transcriptomic and metabolomic analyses identifies CsERF003, a citrus ERF transcription factor, as flavonoid activator. Plant Sci. 2023, 334, 111762. [Google Scholar] [CrossRef]
- Zhuo, M.G.; Wang, T.Y.; Huang, X.M.; Hu, G.B.; Zhou, B.Y.; Wang, H.C.; Abbas, F. ERF transcription factors govern anthocyanin biosynthesis in litchi pericarp by modulating the expression of anthocyanin biosynthesis genes. Sci. Hortic. 2024, 337, 113464. [Google Scholar] [CrossRef]
- Yue, L.Q.; Kang, Y.Y.; Zhong, M.; Kang, D.J.; Zhao, P.Y.; Chai, X.R.; Yang, X. Melatonin delays postharvest senescence through suppressing the inhibition of BrERF2/BrERF109 on flavonoid biosynthesis in flowering chinese cabbage. Int. J. Mol. Sci. 2023, 24, 2933. [Google Scholar] [CrossRef]
- Liu, Y.; Guan, C.N.; Chen, Y.Y.; Shi, Y.L.; Long, O.; Lin, H.; Zhang, K.X.; Zhou, M.L. Evolutionary analysis of MADS-box genes in buckwheat species and functional study of FdMADS28 in flavonoid metabolism. Plant Physiol. Biochem. 2024, 210, 108637. [Google Scholar] [CrossRef]
- Wang, N.; Liu, W.J.; Zhang, T.L.; Jiang, S.H.; Xu, H.F.; Wang, Y.C.; Zhang, Z.Y.; Wang, C.Z.; Chen, X.S. Transcriptomic analysis of red-fleshed apples reveals the novel role of MdWRKY11 in flavonoid and anthocyanin biosynthesis. J. Agric. Food Chem. 2018, 66, 7076–7086. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mou, Z.; Yuan, Y.; Wei, W.; Zhao, Y.; Wu, B.; Chen, J. Integrative Metabolomic and Transcriptomic Analysis Provides Novel Insights into the Effects of SO2 on the Postharvest Quality of ‘Munage’ Table Grapes. Foods 2024, 13, 3494. https://doi.org/10.3390/foods13213494
Mou Z, Yuan Y, Wei W, Zhao Y, Wu B, Chen J. Integrative Metabolomic and Transcriptomic Analysis Provides Novel Insights into the Effects of SO2 on the Postharvest Quality of ‘Munage’ Table Grapes. Foods. 2024; 13(21):3494. https://doi.org/10.3390/foods13213494
Chicago/Turabian StyleMou, Zhenliang, Yuyao Yuan, Wei Wei, Yating Zhao, Bin Wu, and Jianye Chen. 2024. "Integrative Metabolomic and Transcriptomic Analysis Provides Novel Insights into the Effects of SO2 on the Postharvest Quality of ‘Munage’ Table Grapes" Foods 13, no. 21: 3494. https://doi.org/10.3390/foods13213494
APA StyleMou, Z., Yuan, Y., Wei, W., Zhao, Y., Wu, B., & Chen, J. (2024). Integrative Metabolomic and Transcriptomic Analysis Provides Novel Insights into the Effects of SO2 on the Postharvest Quality of ‘Munage’ Table Grapes. Foods, 13(21), 3494. https://doi.org/10.3390/foods13213494