
Investigation of the Alternations in Lipid Oxidation and Lipase Activity in Air-Dried Hairtail (Trichiurus lepturus) during Chilled Storage
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Hairtail Samples and Treatments
2.3. Water Activity, Water Content, and NaCl Content Analysis
2.4. Lipid Oxidation Analysis
2.4.1. Peroxide Value (PV) Analysis
2.4.2. Para-Anisidine Value (p-AnV) Analysis
2.5. Non-Esterified Fatty Acid (NEFA) Content Analysis
2.6. Thiobarbituric Acid-Reactive Substance (TBARS) Value Analysis
2.7. Lipase Enzyme Activity Analysis
2.7.1. Crude Lipase Enzyme Extraction
2.7.2. Neutral Lipase Activity Analysis
2.7.3. Alkaline Lipase Activity Analysis
2.8. Lipoxygenase Activity Analysis
2.9. Data Analysis
3. Results and Discussion
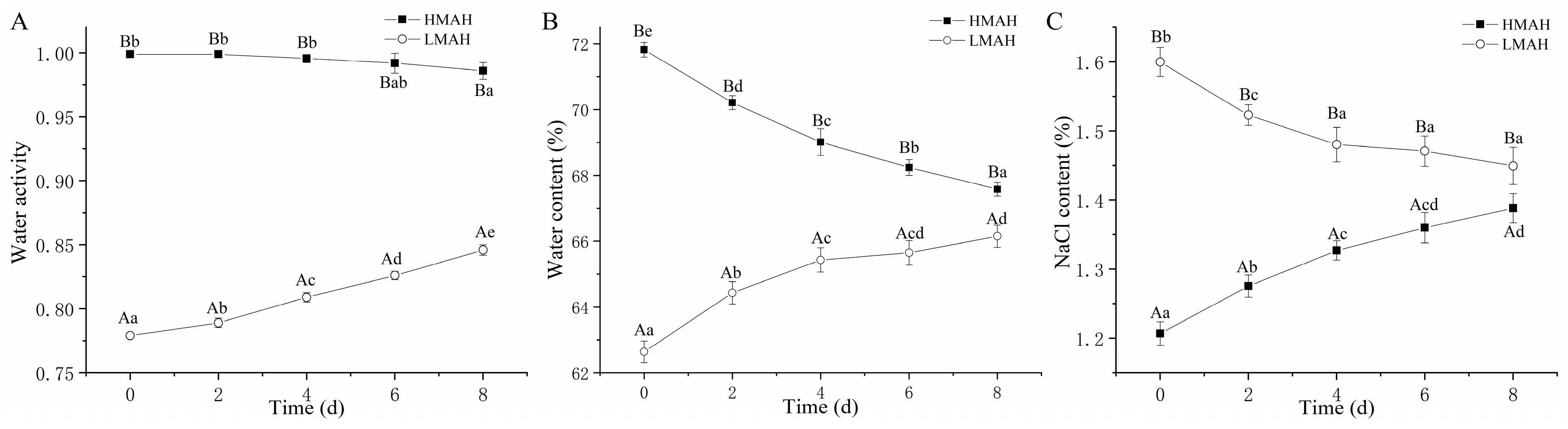
3.1. Water Activity, Water Content, and NaCl Content Analysis
3.2. Primary Oxidation Analysis
3.3. Secondary Oxidation Analysis
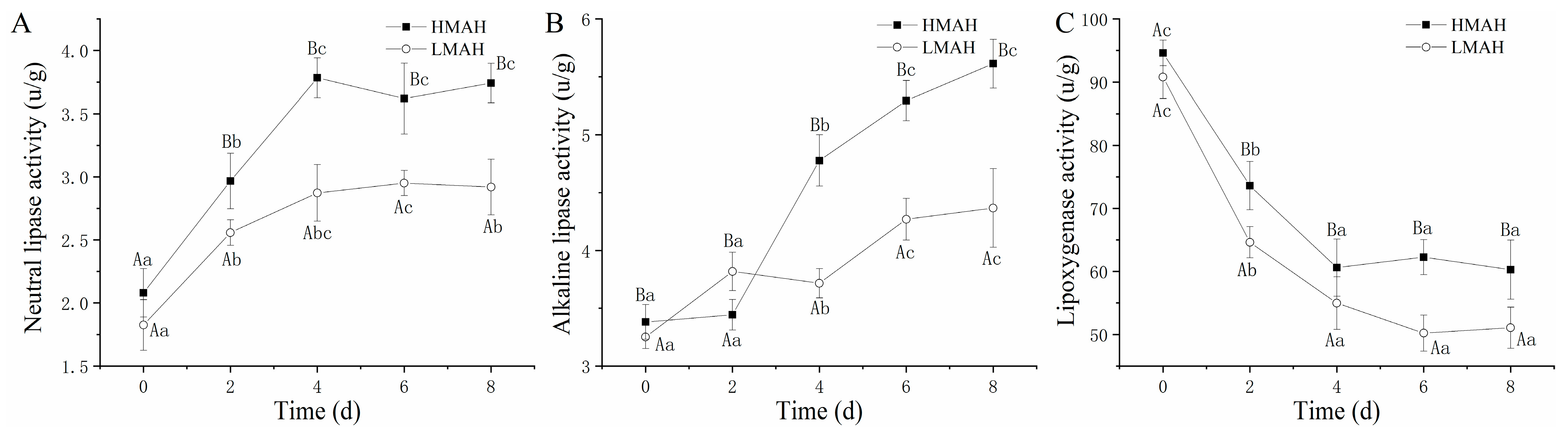
3.4. Neutral Lipase, Alkaline Lipase, and Lipoxygenase Activity Analysis
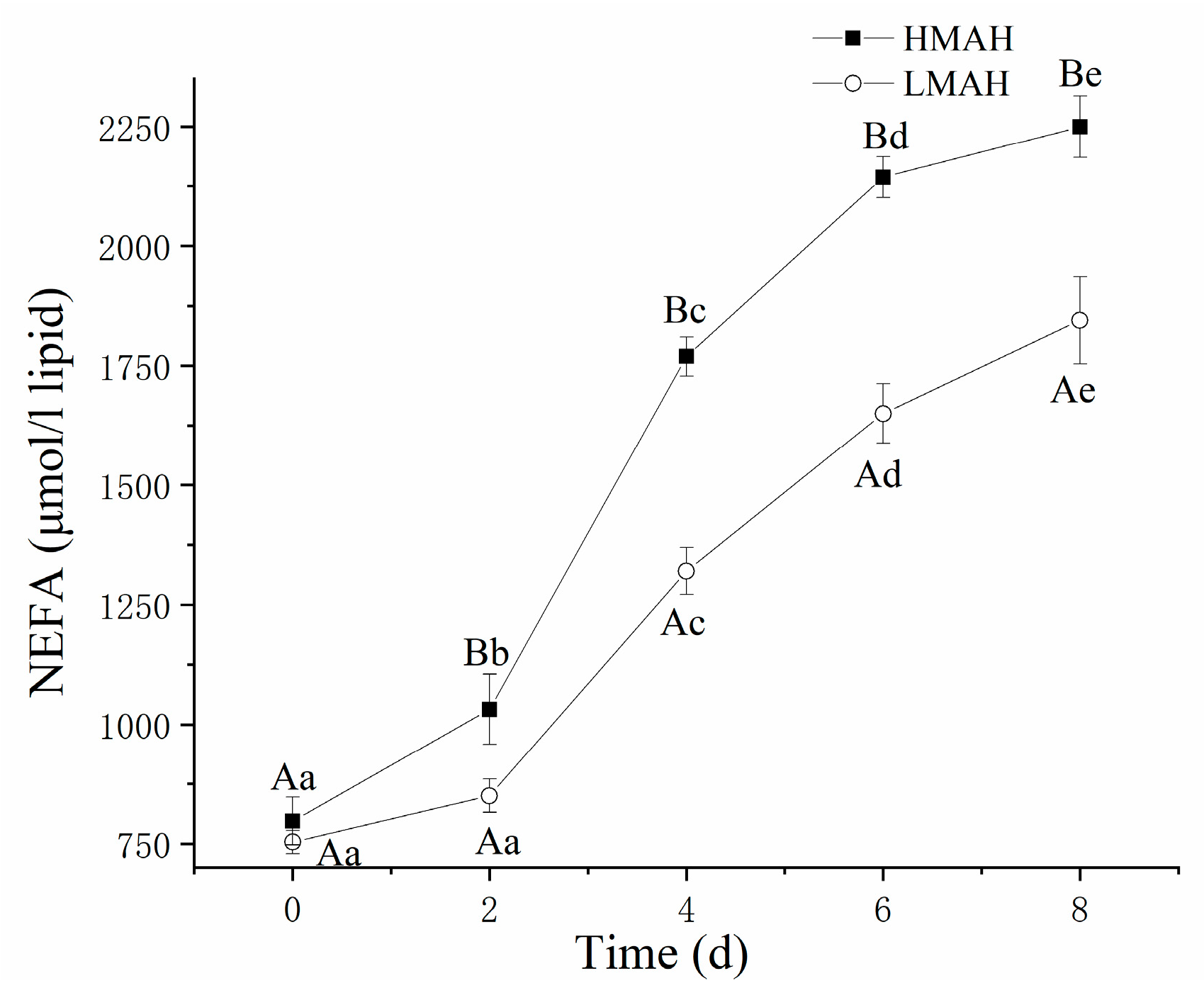
3.5. NEFA Content Analysis
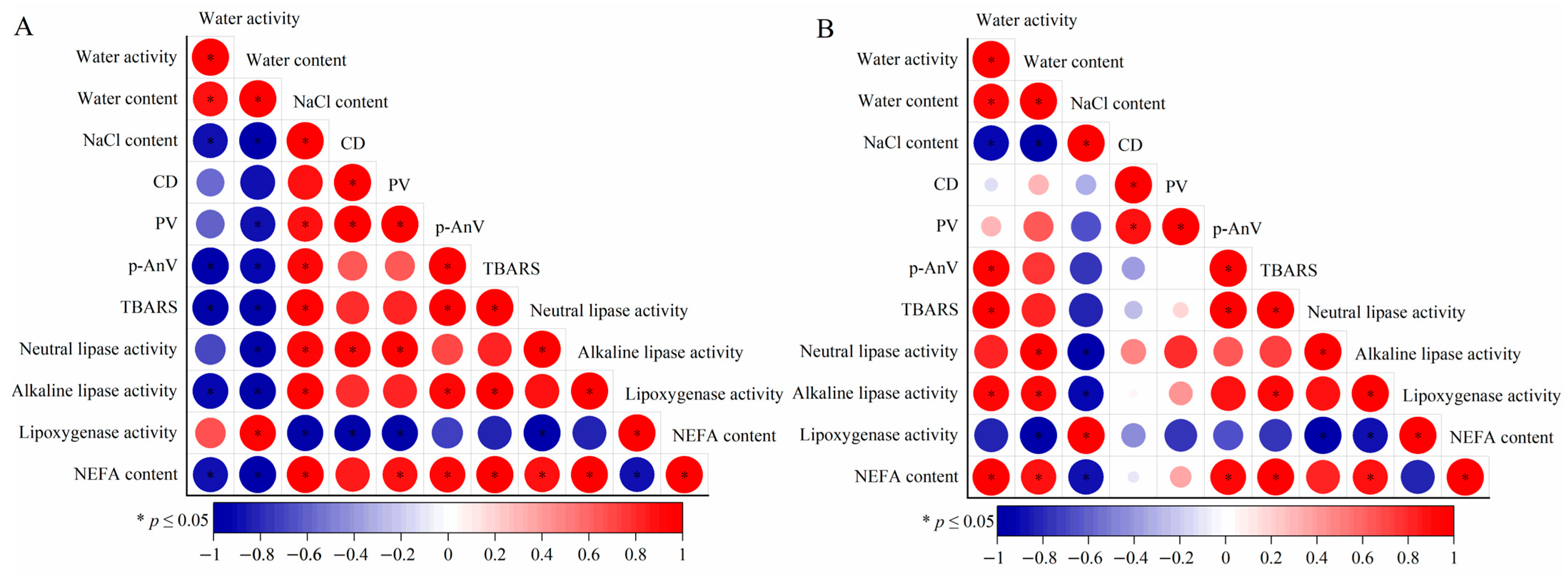
3.6. Correlation Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, X.Z.; Cui, L.F.; Jiang, K.Y.; Yuan, X.C.; Yu, X.J. 2022 China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2023. [Google Scholar]
- Wu, Y.; Cai, Q.; Li, L.; Wang, Y.; Yang, X. Comparison of the Changes in Fatty Acids and Triacylglycerols between Decapterus maruadsi and Trichiurus lepturus during Salt-Dried Process. J. Oleo Sci. 2019, 68, 769–779. [Google Scholar] [CrossRef]
- Liu, J.; Shao, Y.; Yuan, C.; Takaki, K.; Li, Y.; Ying, Y.; Hu, Y. Eugenol-Chitosan Nanoemulsion as an Edible Coating: Its Impact on Physicochemical, Microbiological and Sensorial Properties of Hairtail (Trichiurus haumela) during Storage at 4 °C. Int. J. Biol. Macromol. 2021, 183, 2199–2204. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Miao, W.; Zheng, B.; Deng, S.; Hatab, S. Assessment of the Effect of Cold Atmospheric Plasma (CAP) on the Hairtail (Trichiurus lepturus) Quality under Cold Storage Conditions. Foods 2022, 11, 3683. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Ding, Y.; Du, Q.; Wu, Y.; Lin, H.; Benjakul, S.; Zhang, B. Changes in the Lipid Profiles of Hairtail (Trichiurus lepturus) Muscle during Air-Drying via Liquid Chromatography-Mass Spectrometry Analysis. Food Chem. X 2023, 17, 100610. [Google Scholar] [CrossRef]
- Liao, Y.; Ding, Y.; Wu, Y.; Du, Q.; Xia, J.; Jia, J.; Lin, H.; Benjakul, S.; Zhang, B.; Hu, Y. Analysis of Volatile Compounds and Flavor Fingerprint in Hairtail (Trichiurus lepturus) during Air-Drying Using Headspace-Gas Chromatography-Ion Mobility Spectrometry (HS-GC-IMS). Front. Nutr. 2022, 9, 1088128. [Google Scholar] [CrossRef] [PubMed]
- Dai, W.; Yan, C.; Ding, Y.; Wang, W.; Gu, S.; Xu, Z.; Zhou, X.; Ding, Y. Effect of a Chitosan Coating Incorporating Epigallocatechin Gallate on the Quality and Shelf Life of Bighead Carp (Aristichthys nobilis) Fillets during Chilled Storage. Int. J. Biol. Macromol. 2022, 219, 1272–1283. [Google Scholar] [CrossRef]
- Duarte, A.M.; Silva, F.; Pinto, F.R.; Barroso, S.; Gil, M.M. Quality Assessment of Chilled and Frozen Fish—Mini Review. Foods 2020, 9, 1739. [Google Scholar] [CrossRef]
- Tsikas, D. Assessment of Lipid Peroxidation by Measuring Malondialdehyde (MDA) and Relatives in Biological Samples: Analytical and Biological Challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Mohdaly, A.A.A.; Mahmoud, A.A.; Ramadan, M.F.; Roby, M.H.H. Biochemical and Microbiological Characteristics of Some Mediterranean Salted Fish Products. Fis. Acc. Lincei 2021, 32, 343–355. [Google Scholar] [CrossRef]
- Grabner, G.F.; Xie, H.; Schweiger, M.; Zechner, R. Lipolysis: Cellular Mechanisms for Lipid Mobilization from Fat Stores. Nat. Metab. 2021, 3, 1445–1465. [Google Scholar] [CrossRef]
- Warner, T.G.; Dambach, L.M.; Shin, J.H.; O’Brien, J.S. Separation and Characterization of the Acid Lipase and Neutral Esterases from Human Liver. Am. J. Hum. Genet. 1980, 32, 869–879. [Google Scholar] [PubMed]
- Santos, M.M.F.; Lima, D.A.S.; Madruga, M.S.; Silva, F.A.P. Lipid and Protein Oxidation of Emulsified Chicken Patties Prepared Using Abdominal Fat and Skin. Poult. Sci. 2020, 99, 1777–1787. [Google Scholar] [CrossRef] [PubMed]
- Bouchaâla, E.; BouAli, M.; Ali, Y.B.; Miled, N.; Gargouri, Y.; Fendri, A. Biochemical Characterization and Molecular Modeling of Pancreatic Lipase from a Cartilaginous Fish, the Common Stingray (Dasyatis pastinaca). Appl. Biochem. Biotechnol. 2015, 176, 151–169. [Google Scholar] [CrossRef] [PubMed]
- González-Félix, M.; Santana-Bejarano, E.; Perez Velazquez, M.; Villalba-Villalba, A. Partial Characterization, Quantification and Activity of Pancreatic Lipase in the Gastrointestinal Tract of Totoaba Macdonaldi. Arch. Biol. Sci. 2018, 70, 489–496. [Google Scholar] [CrossRef]
- Concha-Frías, B.; Gaxiola-Cortes, M.G.; De la Cruz-Alvarado, F.J.; Jimenez Martinez, L.D.; Peña-Marin, E.S.; Oliva-Arriagada, M.A.; Arias-Moscoso, J.L.; Alvarez-González, C.A. Intestinal Lipase Characterization in Common Snook (Centropomus undecimalis) Juveniles. Fishes 2022, 7, 107. [Google Scholar] [CrossRef]
- Feng, D.; Huang, Q.Y.; Liu, K.; Zhang, S.C.; Liu, Z.H. Comparative Studies of Zebrafish Danio rerio Lipoprotein Lipase (lpl) and Hepatic Lipase (lipc) Genes Belonging to the Lipase Gene Family: Evolution and Expression Pattern. J. Fish Biol. 2014, 85, 329–342. [Google Scholar] [CrossRef] [PubMed]
- Tian, Z.; Wei, M.; Xue, R.; Song, L.; Li, H.; Ji, H.; Sun, J. Lpla (Lipoprotein Lipase) Is a Marker of Early Adipogenesis Rather than Late Adipogenesis in Grass Carp (Ctenopharyngodon idellus). Fish Physiol. Biochem. 2023, 49, 1229–1239. [Google Scholar] [CrossRef]
- Yang, J.; Liang, R.; Mao, Y.; Dong, P.; Zhu, L.; Luo, X.; Zhang, Y.; Yang, X. Potential Inhibitory Effect of Carbon Dioxide on the Spoilage Behaviors of Pseudomonas Fragi in High-Oxygen Packaged Beef during Refrigerated Storage. Food Microbiol. 2023, 112, 104229. [Google Scholar] [CrossRef]
- Coupland, J.N.; McClements, D.J. Lipid oxidation in food emulsions. Trends Food Sci. Technol. 1996, 7, 83–91. [Google Scholar] [CrossRef]
- Ahmed, M.; Pickova, J.; Ahmad, T.; Liaquat, M.; Farid, A.; Jahangir, M. Oxidation of Lipids in Foods. Sarhad J. Agric. 2016, 32, 230–238. [Google Scholar] [CrossRef]
- Hideg, É. Detection of Free Radicals and Reactive Oxygen Species. In Photosynthesis Research Protocols; Humana Press: Totowa, NJ, USA, 2004; Volume 274, pp. 249–260. ISBN 978-1-59259-799-4. [Google Scholar]
- Leães, Y.S.V.; Lorenzo, J.M.; Cichoski, A.J.; Wagner, R.; Santos, E.M.; Reyes, J.F.; Campagnol, P.C.B. Lipid Oxidation (Primary and Secondary Products). In Methods to Assess the Quality of Meat Products; Lorenzo, J.M., Domínguez, R., Pateiro, M., Munekata, P.E.S., Eds.; Methods and Protocols in Food Science; Springer: New York, NY, USA, 2022; pp. 115–132. ISBN 978-1-07-162002-1. [Google Scholar]
- Jin, G.; He, L.; Zhang, J.; Yu, X.; Wang, J.; Huang, F. Effects of Temperature and NaCl Percentage on Lipid Oxidation in Pork Muscle and Exploration of the Controlling Method Using Response Surface Methodology (RSM). Food Chem. 2012, 131, 817–825. [Google Scholar] [CrossRef]
- Vu, T.P.; He, L.; McClements, D.J.; Decker, E.A. Effects of Water Activity, Sugars, and Proteins on Lipid Oxidative Stability of Low Moisture Model Crackers. Food Res. Int. 2020, 130, 108844. [Google Scholar] [CrossRef] [PubMed]
- Cho, B.; Charoensri, K.; Doh, H.; Park, H. jin Preparation of Colorimetric Sensor Array System to Evaluate the Effects of Alginate Edible Coating on Boiled-Dried Anchovy. Foods 2023, 12, 638. [Google Scholar] [CrossRef] [PubMed]
- ISO 18787:2017; Determination of Water Activity. International Standards Foodstuffs. ISO: Geneva, Switzerland, 2017.
- ISO 1442:1997; Determination of Moisture Content. International Standards Meat and Meat Products. ISO: Geneva, Switzerland, 1997.
- ISO 1841-1:1996; Determination of Chloride Content, Part 1: Volhard Method. International Standards Meat and Meat Products. ISO: Geneva, Switzerland, 1996.
- Pawliszyn, J.; Mondello, L.; Dugo, P. Comprehensive Sampling and Sample Preparation: Analytical Techniques for Scientists; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- GB 5009.227; Determination of Peroxide Value in Food. National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016. (In Chinese)
- GB/T 24304; Animal and Vegetable Fats and Oils Determination of Anisidine Value (ISO 6885: 2006, IDT). National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2009. (In Chinese)
- Nan, X.; Wu, Q.; Nan, S.; Zeng, X.; Dai, Y.; Kang, L. Effect of Oil Oxidation on Acrylamide Formation in Oil-Rich Model Systems without the Participation of Reducing Sugars. J. Food Prot. 2020, 83, 342–349. [Google Scholar] [CrossRef]
- GB 5009.181; Determination of Malondialdehyde in Food. National Health and Family Planning Commission of the People’s Republic of China: Beijing, China, 2016. (In Chinese)
- Hernández, P.; Navarro, J.-L.; Toldrá, F. Lipid Composition and Lipolytic Enzyme Activities in Porcine Skeletal Muscles with Different Oxidative Pattern. Meat Sci. 1998, 49, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Motilva, M.-J.; Toldrá, F.; Flores, J. Assay of Lipase and Esterase Activities in Fresh Pork Meat and Dry-Cured Ham. Z Leb. Unters Forch 1992, 195, 446–450. [Google Scholar] [CrossRef]
- Bicas, J.L.; de Quadros, C.P.; Néri-Numa, I.A.; Pastore, G.M. Integrated Process for Co-Production of Alkaline Lipase and R-(+)-α-Terpineol by Fusarium Oxysporum. Food Chem. 2010, 120, 452–456. [Google Scholar] [CrossRef]
- Gata, J.L.; Pinto, M.C.; Macías, P. Lipoxygenase Activity in Pig Muscle: Purification and Partial Characterization. J. Agric. Food Chem. 1996, 44, 2573–2577. [Google Scholar] [CrossRef]
- Bian, H.; Ma, J.; Geng, Z.; Liu, T.; Sun, C.; Wang, D.; Zhang, M.; Xu, W. Changes of Hydroxyl-Linoleic Acids during Chinese-Style Sausage Processing and Their Relationships with Lipids Oxidation. Food Chem. 2019, 296, 63–68. [Google Scholar] [CrossRef]
- Yan, H.; Jiao, L.; Fang, C.; Benjakul, S.; Zhang, B. Chemical and LC-MS-Based Lipidomics Analyses Revealed Changes in Lipid Profiles in Hairtail (Trichiurus haumela) Muscle during Chilled Storage. Food Res. Int. 2022, 159, 111600. [Google Scholar] [CrossRef]
- Zhang, B.; Yan, H.; Su, L.; Chen, X. Kappa-Carrageenan Oligosaccharides Retard the Progression of Protein and Lipid Oxidation in Mackerel (Scomber japonicus) Fillets during Frozen Storage. RSC Adv. 2020, 10, 20827–20836. [Google Scholar] [CrossRef] [PubMed]
- Adawiyah, D.; Soekarto, S.; Hariyadi, P. Fat Hydrolysis in a Food Model System: Effect of Water Activity and Glass Transition. Int. Food Res. J. 2012, 19, 737–741. [Google Scholar]
- Yuan, Y.; Cai, W.; Wang, M.; Liu, Y.; Fu, J.; Chen, Y. Effects of Sous Vide Cooking Combined with Ultrasound Pretreatment on Physicochemical Properties and Microbial Communities of Russian Sturgeon Meat (Acipenser gueldenstaedti). J. Sci. Food Agric. 2023, 103, 2166–2174. [Google Scholar] [CrossRef]
- Cao, Q.; Du, H.; Huang, Y.; Hu, Y.; You, J.; Liu, R.; Xiong, S.; Manyande, A. The Inhibitory Effect of Chlorogenic Acid on Lipid Oxidation of Grass Carp (Ctenopharyngodon idellus) during Chilled Storage. Food Bioprocess Technol. 2019, 12, 2050–2061. [Google Scholar] [CrossRef]
- Javed, S.; Azeem, F.; Hussain, S.; Rasul, I.; Siddique, M.H.; Riaz, M.; Afzal, M.; Kouser, A.; Nadeem, H. Bacterial Lipases: A Review on Purification and Characterization. Prog. Biophys. Mol. Biol. 2018, 132, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Aro, T.L.; Larmo, P.S.; Bäckman, C.H.; Kallio, H.P.; Tahvonen, R.L. Fatty Acids and Fat-Soluble Vitamins in Salted Herring (Clupea harengus) Products. J. Agric. Food Chem. 2005, 53, 1482–1488. [Google Scholar] [CrossRef] [PubMed]
- Guillén, M.D.; Cabo, N. Fourier Transform Infrared Spectra Data versus Peroxide and Anisidine Values to Determine Oxidative Stability of Edible Oils. Food Chem. 2002, 77, 503–510. [Google Scholar] [CrossRef]
- Verger, R. ‘Interfacial Activation’ of Lipases: Facts and Artifacts. Trends Biotechnol. 1997, 15, 32–38. [Google Scholar] [CrossRef]
- Jittrepotch, N.; Ushio, H.; Ohshima, T. Effects of EDTA and a Combined Use of Nitrite and Ascorbate on Lipid Oxidation in Cooked Japanese Sardine (Sardinops melanostictus) during Refrigerated Storage. Food Chem. 2006, 99, 70–82. [Google Scholar] [CrossRef]
- Fredrikson, G.; Tornqvist, H.; Belfrage, P. Hormone-Sensitive Lipase and Monoacylglycerol Lipase Are Both Required for Complete Degradation of Adipocyte Triacylglycerol. Biochim. Biophys. Acta 1986, 876, 288–293. [Google Scholar] [CrossRef]
- Sae-leaw, T.; Benjakul, S. Lipase from Liver of Seabass (Lates calcarifer): Characteristics and the Use for Defatting of Fish Skin. Food Chem. 2018, 240, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Jahan, F.; Mahajan, R.V.; Saxena, R.K. Efficient Regioselective Acylation of Quercetin Using Rhizopus Oryzae Lipase and Its Potential as Antioxidant. Bioresour. Technol. 2016, 218, 1246–1248. [Google Scholar] [CrossRef] [PubMed]
- Gardner, H.W. Lipid Enzymes: Lipases, Lipoxygenases, and “Hydroperoxidases”. In Autoxidation in Food and Biological Systems; Simic, M.G., Karel, M., Eds.; Springer: Boston, MA, USA, 1980; pp. 447–504. ISBN 978-1-4757-9351-2. [Google Scholar]
- Veldink, G.A.; Vliegenthart, J.F.G.; Boldingh, J. Plant Lipoxygenases. Prog. Chem. Fats Other Lipids 1977, 15, 131–166. [Google Scholar] [CrossRef]
- Frega, N.; Mozzon, M.; Lercker, G. Effects of Free Fatty Acids on Oxidative Stability of Vegetable Oil. J. Am. Oil Chem. Soc. 1999, 76, 325–329. [Google Scholar] [CrossRef]
- Vázquez, M.; Fidalgo, L.G.; Saraiva, J.A.; Aubourg, S.P. Preservative Effect of a Previous High-Pressure Treatment on the Chemical Changes Related to Quality Loss in Frozen Hake (Merluccius merluccius). Food Bioprocess Technol. 2018, 11, 293–304. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhan, Y.; Li, J.; Li, T.; Xie, K.; Tu, C.; Liu, Z.; Pang, J.; Zhang, B. Investigation of the Alternations in Lipid Oxidation and Lipase Activity in Air-Dried Hairtail (Trichiurus lepturus) during Chilled Storage. Foods 2024, 13, 229. https://doi.org/10.3390/foods13020229
Zhan Y, Li J, Li T, Xie K, Tu C, Liu Z, Pang J, Zhang B. Investigation of the Alternations in Lipid Oxidation and Lipase Activity in Air-Dried Hairtail (Trichiurus lepturus) during Chilled Storage. Foods. 2024; 13(2):229. https://doi.org/10.3390/foods13020229
Chicago/Turabian StyleZhan, Yuexiang, Jiagen Li, Taiyu Li, Kai Xie, Chuanhai Tu, Zhiyu Liu, Jie Pang, and Bin Zhang. 2024. "Investigation of the Alternations in Lipid Oxidation and Lipase Activity in Air-Dried Hairtail (Trichiurus lepturus) during Chilled Storage" Foods 13, no. 2: 229. https://doi.org/10.3390/foods13020229
APA StyleZhan, Y., Li, J., Li, T., Xie, K., Tu, C., Liu, Z., Pang, J., & Zhang, B. (2024). Investigation of the Alternations in Lipid Oxidation and Lipase Activity in Air-Dried Hairtail (Trichiurus lepturus) during Chilled Storage. Foods, 13(2), 229. https://doi.org/10.3390/foods13020229