

Alleviation of Alcoholic Fatty Liver by Dendrobium officinale Flower Extracts due to Regulation of Gut Microbiota and Short-Chain Fatty Acids in Mice Exposed to Chronic Alcohol
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Dendrobium officinale Flower Extract
2.2. Animals and Experimental Design
2.3. Determination of Serum Biochemical Indicators
2.4. Hepatic Histopathological Evaluation by Hematoxylin–Eosin (H&E) Staining
2.5. Analysis of Hepatic Biochemical Indicators
2.6. Analysis of Gut Microbiota
2.7. Measurement of Short-Chain Fatty Acids in Feces
2.8. Statistical Analysis
3. Results and Discussion
3.1. Alleviation of Liver Injury and Lipid Disorder by DOFE
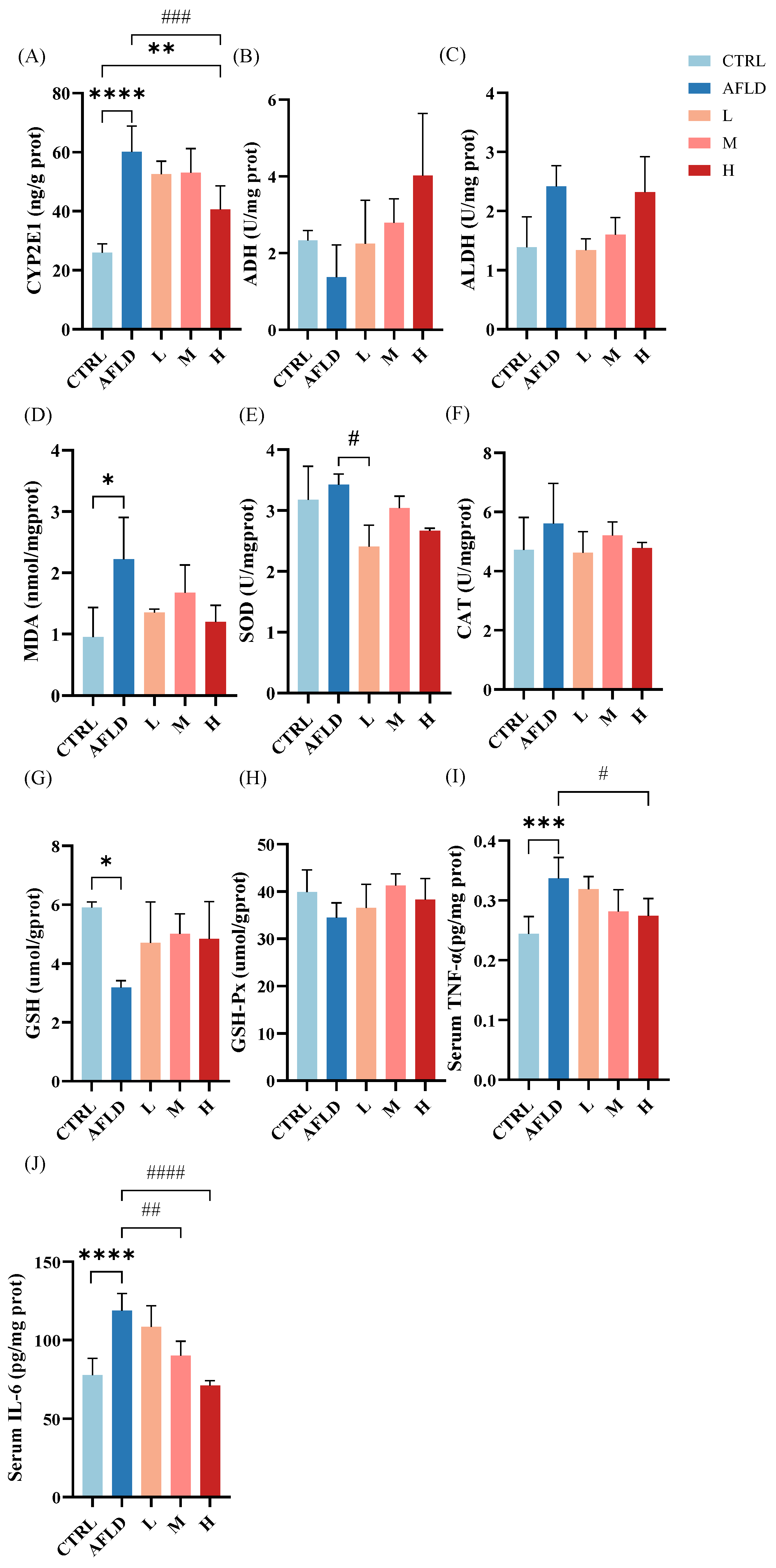
3.2. Effects of DOFE on Alcohol Metabolism, Hepatic Antioxidant Capacity, and Inflammatory Cytokine Levels
3.3. Effects of DOFE on the Diversity and Structure of the Gut Microbiota in Alcohol-Exposed Mice
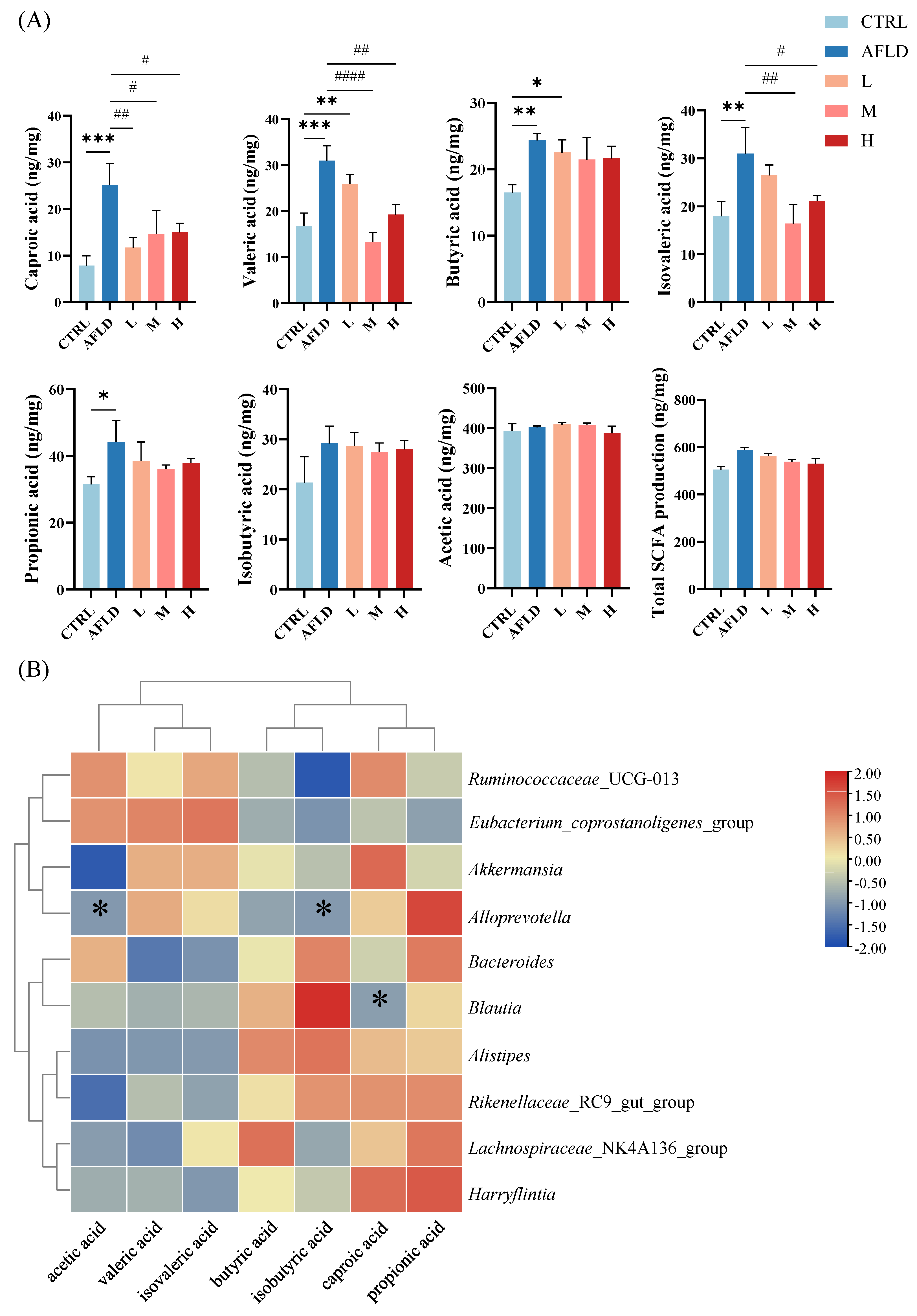
3.4. Effect of DOFE on SCFA
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lang, S.; Schnabl, B. Microbiota and fatty liver disease-the known, the unknown, and the future. Cell Host Microbe 2020, 28, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Fuster, D.; Samet, J.H. Alcohol use in patients with chronic liver disease. N. Engl. J. Med. 2018, 379, 1251–1261. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, T.S.; Liu, M.; Shah, V.H. The knowns and unknowns of treatment for alcoholic hepatitis. Lancet Gastroenterol. Hepatol. 2020, 5, 494–506. [Google Scholar] [CrossRef]
- Louvet, A.; Mathurin, P. Alcoholic liver disease: Mechanisms of injury and targeted treatment. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Correnti, J.; Lin, C.; Brettschneider, J.; Kuriakose, A.; Jeon, S.; Scorletti, E.; Oranu, A.; McIver-Jenkins, D.; Kaneza, I.; Buyco, D.; et al. Liver-specific ceramide reduction alleviates steatosis and insulin resistance in alcohol-fed mice. J. Lipid Res. 2020, 61, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Chen, J.; Ji, X.; Qin, Q.; Yang, H.; Liu, D.; Li, D.; Sun, M. Alcoholic fatty liver disease inhibited the co-expression of fmo5 and pparα to activate the nf-κb signaling pathway, thereby reducing liver injury via inducing gut microbiota disturbance. J. Exp. Clin. Cancer Res. 2021, 40, 18. [Google Scholar] [CrossRef] [PubMed]
- Lackner, C.; Spindelboeck, W.; Haybaeck, J.; Douschan, P.; Rainer, F.; Terracciano, L.; Haas, J.; Berghold, A.; Bataller, R.; Stauber, R.E. Histological parameters and alcohol abstinence determine long-term prognosis in patients with alcoholic liver disease. J. Hepatol. 2016, 66, 610–618. [Google Scholar] [CrossRef]
- Louvet, A.; Labreuche, J.; Artru, F.; Bouthors, A.; Rolland, B.; Saffers, P.; Lollivier, J.; Lemaître, E.; Dharancy, S.; Lassailly, G.; et al. Main drivers of outcome differ between short term and long term in severe alcoholic hepatitis: A prospective study. Hepatology 2017, 66, 1464–1473. [Google Scholar] [CrossRef]
- Singal, A.K.; Bataller, R.; Ahn, J.; Kamath, P.S.; Shah, V.H. ACG Clinical Guideline: Alcoholic Liver Disease. Am. J. Gastroenterol. 2018, 113, 175–194. [Google Scholar] [CrossRef]
- Kurokawa, K.; Itoh, T.; Kuwahara, T.; Oshima, K.; Toh, H.; Toyoda, A.; Takami, H.; Morita, H.; Sharma, V.K.; Srivastava, T.P.; et al. Comparative Metagenomics Revealed Commonly Enriched Gene Sets in Human Gut Microbiomes. DNA Res. 2007, 14, 169–181. [Google Scholar] [CrossRef]
- Bäckhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-Bacterial Mutualism in the Human Intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef] [PubMed]
- Francisco Guarner, J.M. Gut flora in health and disease. Lancet 2003, 361, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Llopis, M.; Cassard, A.M.; Wrzosek, L.; Boschat, L.; Bruneau, A.; Ferrere, G.; Puchois, V.; Martin, J.C.; Lepage, P.; Le Roy, T.; et al. Intestinal microbiota contributes to individual susceptibility to alcoholic liver disease. Gut 2016, 65, 830–839. [Google Scholar] [CrossRef]
- Chen, P.; Stärkel, P.; Turner, J.R.; Ho, S.B.; Schnabl, B. Dysbiosis-induced intestinal inflammation activates tumor necrosis factor receptor I and mediates alcoholic liver disease in mice. Hepatology 2015, 61, 883–894. [Google Scholar] [CrossRef]
- Stärkel, P.; Schnabl, B. Bidirectional Communication between Liver and Gut during Alcoholic Liver Disease. Semin. Liver Dis. 2016, 36, 331–339. [Google Scholar] [CrossRef]
- Lang, S.; Duan, Y.; Liu, J.; Torralba, M.G.; Kuelbs, C.; Ventura-Cots, M.; Abraldes, J.G.; Bosques-Padilla, F.; Verna, E.C.; Brown, R.S., Jr.; et al. Intestinal Fungal Dysbiosis and Systemic Immune Response to Fungi in Patients with Alcoholic Hepatitis. Hepatology 2020, 71, 522–538. [Google Scholar] [CrossRef]
- Ferrere, G.; Wrzosek, L.; Cailleux, F.; Turpin, W.; Puchois, V.; Spatz, M.; Ciocan, D.; Rainteau, D.; Humbert, L.; Hugot, C.; et al. Fecal microbiota manipulation prevents dysbiosis and alcohol-induced liver injury in mice. J. Hepatol. 2016, 66, 806–815. [Google Scholar] [CrossRef] [PubMed]
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712. [Google Scholar] [CrossRef]
- Martin-Gallausiaux, C.; Marinelli, L.; Blottière, H.M.; Larraufie, P.; Lapaque, N. SCFA: Mechanisms and functional importance in the gut. Proc. Nutr. Soc. 2021, 80, 37–49. [Google Scholar] [CrossRef]
- Sun, T.; Yang, J.; Lu, L. Salidroside Improves Antibiotic-Induced Gut Microbiota Disturbance and Low Levels of Short-Chain Fatty Acids in Mice. Foods 2022, 11, 3073. [Google Scholar] [CrossRef]
- Shivashankara, A.R.; Azmidah, A.; Haniadka, R.; Rai, M.P.; Arora, R.; Baliga, M.S. Dietary agents in the prevention of alcohol-induced hepatotoxicty: Preclinical observations. Food Funct. 2012, 3, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bian, M.; Jin, H.; Lian, N.; Shao, J.; Zhang, F.; Zheng, S. Dihydroartemisinin attenuates alcoholic fatty liver through regulation of lipin-1 signaling. IUBMB Life 2019, 71, 1740–1750. [Google Scholar] [CrossRef]
- Do, M.H.; Lee, H.H.L.; Park, M.; Oh, M.-J.; Lee, E.; Kweon, M.; Park, H.-Y. Morinda citrifolia Extract Prevents Alcoholic Fatty Liver Disease by Improving Gut Health. J. Med. Food 2022, 25, 1102–1111. [Google Scholar] [CrossRef]
- Wang, X.-Y.; Luo, J.-P.; Chen, R.; Zha, X.-Q.; Wang, H. The effects of daily supplementation of dendrobium huoshanense polysaccharide on ethanol-induced subacute liver injury in mice by proteomic analysis. Food Funct. 2014, 5, 2020–2035. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Zeng, X.; Liu, Y.; Zhan, S.; Wu, Z.; Zheng, X.; Zhang, X. Dendrobium officinale polysaccharide attenuates cognitive impairment in circadian rhythm disruption mice model by modulating gut microbiota. Int. J. Biol. Macromol. 2022, 217, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Kuo, H.-C.; Chen, Y.-H.; Tsai, M.-Y. Current Advances in the Biological Activity of Polysaccharides in Dendrobium with Intriguing Therapeutic Potential. Curr. Med. Chem. 2018, 25, 1663–1681. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Fan, J.; Liu, Q.; Luo, H.; Tang, Q.; Li, C.; Zhao, J.; Zhang, X. Phytochemical profiles of edible flowers of medicinal plants of Dendrobium officinale and Dendrobium devonianum. Food Sci Nutr. 2021, 9, 6575–6586. [Google Scholar] [CrossRef]
- Zhang, X.; Zhang, S.; Gao, B.; Qian, Z.; Liu, J.; Wu, S.; Si, J. Identification and quantitative analysis of phenolic glycosides with antioxidant activity in methanolic extract of Dendrobium catenatum flowers and selection of quality control herb-markers. Food Res. Int. 2019, 123, 732–745. [Google Scholar] [CrossRef]
- Miao, Y.X.; Liao, M.X.; Sun, A.H.; Li, R. Study on Optimum Extraction of Polysaccharides from Flowers of Dendrobium officinale and Its Antioxidantion Activity in vitro. Food Res. Dev. 2019, 40, 52–56. [Google Scholar]
- Liang, K.-L.; Fang, P.; Shi, Q.-Q.; Su, J.; Li, B.; Chen, S.-H.; Lv, G.-Y. Antihypertensive effect and mechanism of Dendrobium officinale flos on high-blood pressure rats induced by high glucose and high fat compound alcohol. China J. Chin. Mater. Med. 2018, 43, 147–153. [Google Scholar]
- Chen, S.H.; Lv, G.Y.; Yan, M.Q. Progress in pharmacological effects related to efficacy of Dendrobium officinale. China J. Chin. Mater. Med. 2013, 4, 5. [Google Scholar]
- Li, B.-Y.; Li, H.-Y.; Zhou, D.-D.; Huang, S.-Y.; Luo, M.; Gan, R.-Y.; Mao, Q.-Q.; Saimaiti, A.; Shang, A.; Li, H.-B. Effects of Different Green Tea Extracts on Chronic Alcohol Induced-Fatty Liver Disease by Ameliorating Oxidative Stress and Inflammation in Mice. Oxidative Med. Cell. Longev. 2021, 2021, 5188205. [Google Scholar] [CrossRef] [PubMed]
- Nie, W.; Xu, F.; Zhou, K.; Yang, X.; Zhou, H.; Xu, B. Stearic acid prevent alcohol-induced liver damage by regulating the gut microbiota. Food Res. Int. 2022, 155, 111095. [Google Scholar] [CrossRef]
- Li, B.; Mao, Q.; Zhou, D.; Luo, M.; Gan, R.; Li, H.; Huang, S.; Saimaiti, A.; Shang, A.; Li, H. Effects of Tea against Alcoholic Fatty Liver Disease by Modulating Gut Microbiota in Chronic Alcohol-Exposed Mice. Foods 2021, 10, 1232. [Google Scholar] [CrossRef]
- Lu, K.-H.; Weng, C.-Y.; Chen, W.-C.; Sheen, L.-Y. Ginseng essence, a medicinal and edible herbal formulation, ameliorates carbon tetrachloride-induced oxidative stress and liver injury in rats. J. Ginseng Res. 2017, 41, 316–325. [Google Scholar] [CrossRef]
- Bokulich, N.A.; Subramanian, S. Quality-filtering vastly improves diversity estimates from illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Jiang, X.; Zhao, Y.; Zhai, X.; Yang, X. Komagataeibacter hansenii cgmcc 3917 alleviates alcohol-induced liver injury by regulating fatty acid metabolism and intestinal microbiota diversity in mice. Food Funct. 2020, 11, 4591–4604. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Cao, Q.; Yang, F.; Zhu, H.; Xu, S.; Chen, Q.; Wang, Z.; Lin, Y.; Cinar, R.; Pawlosky, R.J.; et al. Brain ethanol metabolism by astrocytic ALDH2 drives the behavioural effects of ethanol intoxication. Nat. Metab. 2021, 3, 337–351. [Google Scholar] [CrossRef]
- Chambers, G.K. The genetics of human alcohol metabolism. Gen. Pharmacol. Vasc. Syst. 1990, 21, 267–272. [Google Scholar] [CrossRef]
- Liu, J.; Wang, X.; Peng, Z.; Zhang, T.; Wu, H.; Yu, W.; Kong, D.; Liu, Y.; Bai, H.; Liu, R.; et al. The effects of Insulin Pre-Administration in Mice Exposed to Ethanol: Alleviating Hepatic Oxidative Injury through Anti-Oxidative, Anti-Apoptotic Activities and Deteriorating Hepatic Steatosis through SRBEP-1c Activation. Int. J. Biol. Sci. 2015, 11, 569–586. [Google Scholar] [CrossRef]
- Martino, C.; Zaramela, L.S.; Gao, B.; Embree, M.; Tarasova, J.; Parker, S.J.; Wang, Y.; Chu, H.; Chen, P.; Lee, K.-C.; et al. Acetate reprograms gut microbiota during alcohol consumption. Nat. Commun. 2022, 13, 4630. [Google Scholar] [CrossRef]
- Siregar, A.S.; Nyiramana, M.M.; Kim, E.-J.; Shin, E.-J.; Woo, M.S.; Kim, J.-M.; Kim, J.H.; Lee, D.K.; Hahm, J.R.; Kim, H.J.; et al. Dipeptide YA is Responsible for the Positive Effect of Oyster Hydrolysates on Alcohol Metabolism in Single Ethanol Binge Rodent Models. Mar. Drugs 2020, 18, 512. [Google Scholar] [CrossRef]
- Xu, X.-Y.; Zheng, J.; Meng, J.-M.; Gan, R.-Y.; Mao, Q.-Q.; Shang, A.; Li, B.-Y.; Wei, X.-L.; Li, H.-B. Effects of Food Processing on In Vivo Antioxidant and Hepatoprotective Properties of Green Tea Extracts. Antioxidants 2019, 8, 572. [Google Scholar] [CrossRef]
- Wu, W.; Lv, L.; Shi, D.; Ye, J.; Fang, D.; Guo, F.; Li, Y.; He, X.; Li, L. Protective Effect of Akkermansia muciniphila against Immune-Mediated Liver Injury in a Mouse Model. Front. Microbiol. 2017, 8, 1804. [Google Scholar] [CrossRef]
- Morrison, D.J.; Preston, T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes 2016, 7, 189–200. [Google Scholar] [CrossRef]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Tett, A.; Pasolli, E.; Masetti, G.; Ercolini, D.; Segata, N. Prevotella diversity, niches and interactions with the human host. Nat. Rev. Microbiol. 2021, 19, 585–599. [Google Scholar] [CrossRef]
- Liu, X.; Mao, B.; Gu, J.; Wu, J.; Cui, S.; Wang, G.; Zhao, J.; Zhang, H.; Chen, W. Blautia-a new functional genus with potential probiotic properties? Gut Microbes 2021, 13, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hosomi, K.; Saito, M.; Park, J.; Murakami, H.; Shibata, N.; Ando, M.; Nagatake, T.; Konishi, K.; Ohno, H.; Tanisawa, K.; et al. Oral administration of Blautia wexlerae ameliorates obesity and type 2 diabetes via metabolic remodeling of the gut microbiota. Nat. Commun. 2022, 13, 4477. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Peng, D.; Li, W.; Chen, S.; Liu, B.; Huang, P.; Wu, J.; Du, B.; Li, P. Probiotic-fermented Pueraria lobata (Willd.) Ohwi alleviates alcoholic liver injury by enhancing antioxidant defense and modulating gut microbiota. J. Sci. Food Agric. 2022, 102, 6877–6888. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Fan, J.; Luo, H.; Liang, Z.; Guan, Y.; Lei, X.; Bo, N.; Zhao, M. Alleviation of Alcoholic Fatty Liver by Dendrobium officinale Flower Extracts due to Regulation of Gut Microbiota and Short-Chain Fatty Acids in Mice Exposed to Chronic Alcohol. Foods 2023, 12, 1428. https://doi.org/10.3390/foods12071428
Zhang J, Fan J, Luo H, Liang Z, Guan Y, Lei X, Bo N, Zhao M. Alleviation of Alcoholic Fatty Liver by Dendrobium officinale Flower Extracts due to Regulation of Gut Microbiota and Short-Chain Fatty Acids in Mice Exposed to Chronic Alcohol. Foods. 2023; 12(7):1428. https://doi.org/10.3390/foods12071428
Chicago/Turabian StyleZhang, Jingchi, Jiakun Fan, Hui Luo, Zhengwei Liang, Yanhui Guan, Xin Lei, Nianguo Bo, and Ming Zhao. 2023. "Alleviation of Alcoholic Fatty Liver by Dendrobium officinale Flower Extracts due to Regulation of Gut Microbiota and Short-Chain Fatty Acids in Mice Exposed to Chronic Alcohol" Foods 12, no. 7: 1428. https://doi.org/10.3390/foods12071428
APA StyleZhang, J., Fan, J., Luo, H., Liang, Z., Guan, Y., Lei, X., Bo, N., & Zhao, M. (2023). Alleviation of Alcoholic Fatty Liver by Dendrobium officinale Flower Extracts due to Regulation of Gut Microbiota and Short-Chain Fatty Acids in Mice Exposed to Chronic Alcohol. Foods, 12(7), 1428. https://doi.org/10.3390/foods12071428