
Involvement of Versatile Bacteria Belonging to the Genus Arthrobacter in Milk and Dairy Products


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Occurrence of Arthrobacter spp. in Raw Milk and Dairy Products
3. Significance of Arthrobacter spp. in the Dairy Industry
3.1. Association of Arthrobacter spp. with Cheese Production
3.1.1. Implication of Arthrobacter spp. in Cheese Color
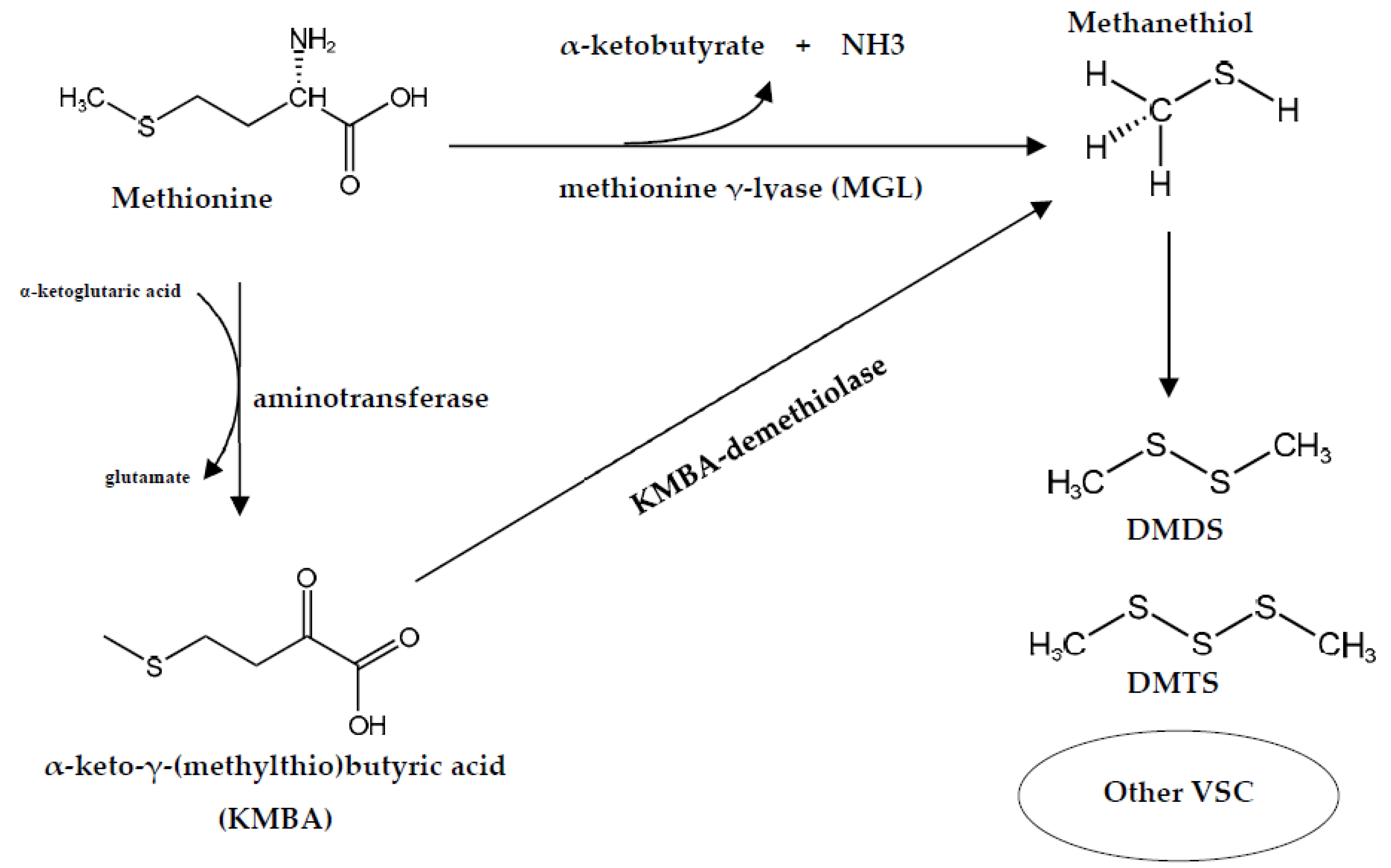
3.1.2. Implication of Arthrobacter spp. in Cheese Flavor
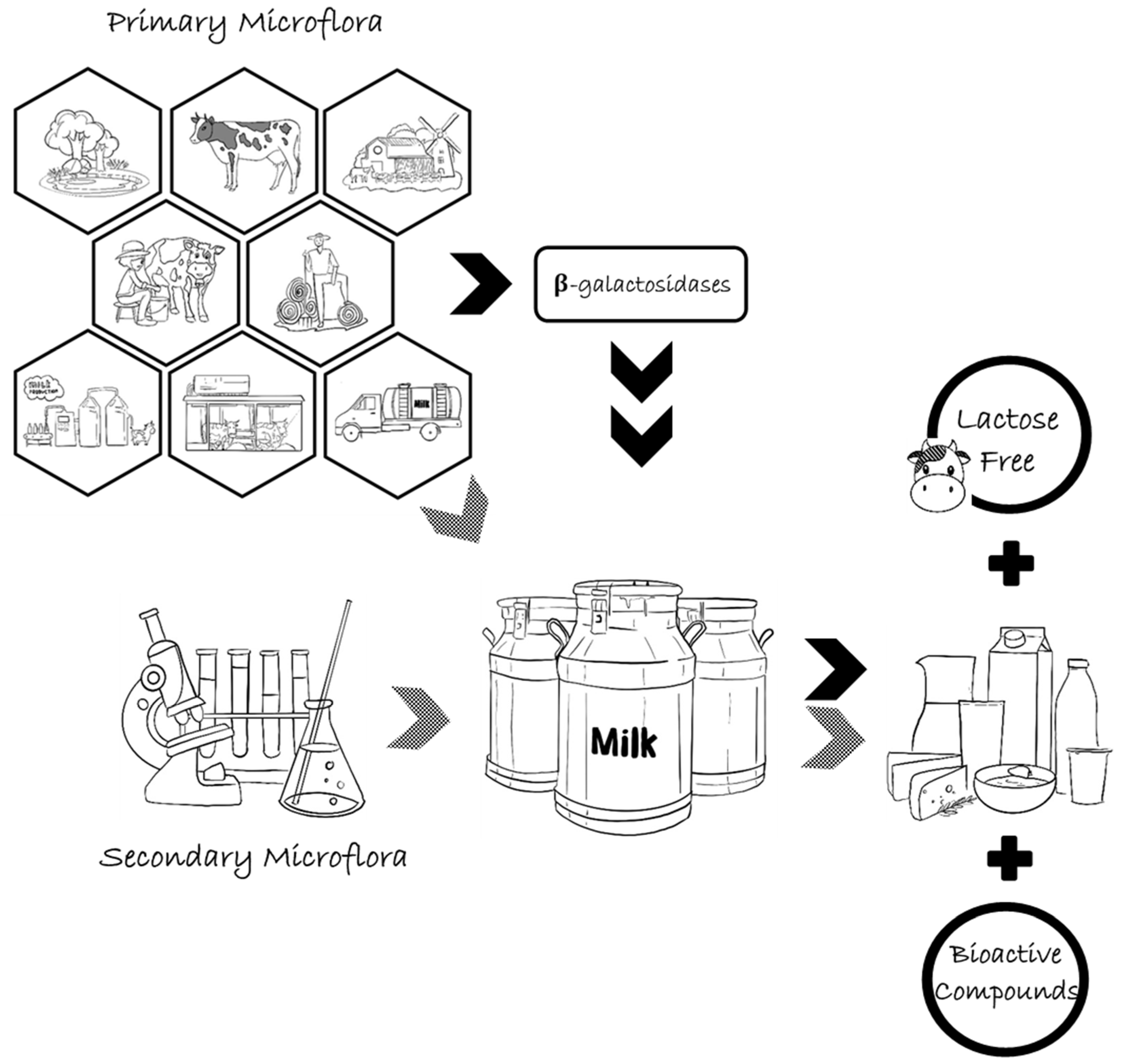
3.2. Association of Arthrobacter spp. Producing Enzymes with Milk and Dairy Products
3.3. Effect of Arthrobacter spp. on Nutritional Components in Milk and Dairy Products
3.3.1. Proteolysis
3.3.2. Lipolysis
3.3.3. Glycolysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guha, S.; Sharma, H.; Deshwal, G.K.; Rao, P.S. A comprehensive review on bioactive peptides derived from milk and milk products of minor dairy species. Food Prod. Process Nutr. 2021, 3, 2. [Google Scholar] [CrossRef]
- Melini, F.; Melini, V.; Luziatelli, F.; Ruzzi, M. Raw and heat-treated milk: From public health risks to nutritional quality. Beverages 2017, 3, 54. [Google Scholar] [CrossRef]
- Fusco, V.; Chieffi, D.; Fanelli, F.; Logrieco, A.F.; Cho, G.-S.; Kabisch, J.; Böhnlein, C.; Franz, C.M.A.P. Microbial quality and safety of milk and milk products in the 21st century. Compr. Rev. Food Sci. Food Saf. 2020, 19, 2013–2049. [Google Scholar] [CrossRef] [PubMed]
- Issa, A.T.; Tahergorabi, R. Milk bacteria and gastrointestinal tract: Microbial composition of milk. In Dietary Interventions in Gastrointestinal Diseases; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Cambridge, MA, USA, 2019; pp. 265–275. [Google Scholar] [CrossRef]
- Shi, C.; Maktabdar, M. Lactic acid bacteria as biopreservation against spoilage molds in dairy products-A review. Front. Microbiol. 2022, 12, 4283. [Google Scholar] [CrossRef]
- Oikonomou, G.; Addis, M.F.; Chassard, C.; Nader-Macias, M.E.F.; Grant, I.; Delbès, C.; Bogni, C.I.; Le Loir, Y.; Even, S. Milk microbiota: What are we exactly talking about? Front. Microbiol. 2020, 11, 60. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Fouillaud, M.; Valla, A.; Caro, Y.; Dufossé, L. Bacteria belonging to the extremely versatile genus Arthrobacter as novel source of natural pigments with extended hue range. Food Res. Int. 2014, 65, 156–162. [Google Scholar] [CrossRef]
- Comi, G.; Carlo, C. Arthrobacter spp. in milk and milk products. In Reference Module in Food Science; Smithers, G., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–7. [Google Scholar] [CrossRef]
- Busse, H.-J.; Wieser, M. The Genus Arthrobacter. In The Prokaryotes; Rosenberg, E., DeLong, E.F., Lory, S., Stackebrandt, E., Thompson, F., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 105–132. [Google Scholar] [CrossRef]
- Roy, P.; Kumar, A. Arthrobacter. In Beneficial Microbes in Agro-Ecology. Bacteria and Fungi; Amaresan, N., Kumar, M.S., Annapurna, K., Kumar, K., Sankaranarayanan, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 3–11. [Google Scholar] [CrossRef]
- List of Prokaryotic Names with Standing in Nomenclature-Genus Arthrobacter. Available online: http://www/bacterio/cict.fr/a/arthrobacter.html (accessed on 2 February 2023).
- Irlinger, F.; Bimet, F.; Delettre, J.; Lefèvre, M.; Grimont, P.A.D. Arthrobacter bergerei sp. nov. and Arthrobacter arilaitensis sp. nov., novel coryneform species isolated from the surfaces of cheeses. Int. J. Syst. Evol. Microbiol. 2005, 55, 457–462. [Google Scholar] [CrossRef]
- Yan, H.; Du, W.; Ji, S.; Guo, C.; Zhang, Y.; Wang, Y.; Cao, Z.; Li, S. Bacterial composition and interactions in raw milk and teat skin of dairy cows. Ferment 2022, 8, 235. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Akabanda, F.; Agyei, D.; Jespersen, L. Microbial safety of milk production and fermented dairy products in Africa. Microorganisms 2020, 8, 752. [Google Scholar] [CrossRef]
- Flegler, A.; Runzheimer, K.; Kombeitz, V.; Mänz, A.T.; von Heidler Heilborn, D.; Etzbach, L.; Schieber, A.; Hölzl, G.; Hüttel, B.; Woehle, C.; et al. Arthrobacter bussei sp. nov., a pink-coloured organism isolated from cheese made of cow’s milk. Int. J. Syst. Evol. Microbiol. 2020, 70, 3027–3036. [Google Scholar] [CrossRef]
- Lauková, A.; Micenková, L.; Grešáková, Ľ.; Maďarová, M.; Simonová, M.P.; Focková, V.; Ščerbová, J. Microbiome associated with Slovak raw goat milk, trace minerals, and vitamin E content. Int. J. Food Sci. 2022, 30, 4595473. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Yan, Y.; Wang, L.; Dong, L.; Pang, X.; Tang, S.; Li, A.; Xiang, A.; Zhang, L.; Zheng, B. High-throughput sequencing reveals bacterial diversity in raw milk production environment and production chain in Tangshan city of China. Food Sci. Anim. Resour. 2021, 41, 452. [Google Scholar] [CrossRef] [PubMed]
- Frétin, M.; Martin, B.; Rifa, E.; Isabelle, V.-M.; Pomiès, D.; Ferlay, A.; Montel, M.C.; Delbès, C. Bacterial community assembly from cow teat skin to ripened cheeses is influenced by grazing systems. Sci. Rep. 2018, 8, 200. [Google Scholar] [CrossRef] [PubMed]
- Hahne, J.; Lipski, A. Growth interferences between bacterial strains from raw cow’s milk and their impact on growth of Listeria monocytogenes and Staphylococcus aureus. J. Appl. Microbiol. 2021, 131, 2019–2032. [Google Scholar] [CrossRef]
- Papademas, P.; Kamilari, E.; Aspri, M.; Anagnostopoulos, A.D.; Mousikos, P.; Kamilaris, A.; Tsaltas, D. Investigation of donkey milk bacterial diversity by 16S rDNA high-throughput sequencing on a Cyprus donkey farm. J. Dairy Sci. 2021, 104, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, M.F.; Gomez, A.P.; Parra, C.M. Molecular and proteomic identification of Arthrobacter gandavensis isolated from cows with subclinical mastitis in a dairy farm. Malays. J. Microbiol. 2022, 18, 309–314. [Google Scholar] [CrossRef]
- Choi, J.; Lee, S.I.; Rackerby, B.; Goddik, L.; Frojen, R.; Ha, S.-D.; Kim, J.H.; Park, S.H. Microbial communities of a variety of cheeses and comparison between core and rind region of cheeses. J. Dairy Sci. 2020, 103, 4026–4042. [Google Scholar] [CrossRef]
- Gobbetti, M.; Rizzello, C.G. Arthrobacter. In Encyclopedia of Food Microbiology, 2nd ed.; Batt, C.A., Tortorello, M.L., Eds.; Academic Press: Cambridge, MA, USA, 2014; pp. 69–76. [Google Scholar] [CrossRef]
- Chauhan, A.; Pathak, A.; Jaswal, R.; Edwards, B., III; Chappell, D.; Ball, C.; Garcia-Sillas, R.; Stothard, P.; Seaman, J. Physiological and comparative genomic analysis of Arthrobacter sp. SRS-W-1-2016 provides insights on niche adaptation for survival in uraniferous soils. Genes 2018, 9, 31. [Google Scholar] [CrossRef]
- Deeth, H.C. Heat treatment of milk: Thermization and pasteurization. In Reference Module in Food Science; Elsevier: Amsterdam, The Netherlands, 2020. [Google Scholar] [CrossRef]
- De Souza, P.V.; Zaffari Grecellé, C.B.; Barreto, F.; Ramírez-Castrillon, M.; Valente, M.; da Costa, M. Bacteria and yeasts associated to Colonial cheese production chain and assessment of their hydrolytic potential. Braz. J. Food Technol. 2021, 24, 1–10. [Google Scholar] [CrossRef]
- Penland, M.; Falentin, H.; Parayre, S.; Pawtowski, A.; Maillard, M.-B.; Thierry, A.; Mounier, J.; Coton, M.; Deutsch, S.-M. Linking Pélardon artisanal goat cheese microbial communities to aroma compounds during cheese-making and ripening. Int. J. Food Microbiol. 2021, 345, 109130. [Google Scholar] [CrossRef]
- Hanlon, M.; Choi, J.; Goddik, L.; Park, S.H. Microbial and chemical composition of Cheddar cheese supplemented with prebiotics from pasteurized milk to aging. J. Dairy Sci. 2022, 105, 2058–2068. [Google Scholar] [CrossRef] [PubMed]
- Ritschard, J.S.; Amato, L.; Kumar, Y.; Müller, B.; Meile, L.; Schuppler, M. The role of the surface smear microbiome in the development of defective smear on surface-ripened red-smear cheese. AIMS Microbiol. 2018, 4, 622–641. [Google Scholar] [CrossRef] [PubMed]
- Cosetta, C.M.; Wolfe, B.E. Deconstructing and reconstructing cheese rind microbiomes for experiments in microbial ecology and evolution. Curr. Protoc. Microbiol. 2020, 56, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Reuben, R.C.; Langer, D.; Eisenhauer, N.; Jurburg, S.D. Universal drivers of cheese microbiomes. iScience 2023, 26, 105744. [Google Scholar] [CrossRef]
- Monnet, C.; Loux, V.; Gibrat, J.F.; Spinnler, E.; Barbe, V.; Vacherie, B.; Gavory, F.; Gourbeyre, E.; Siguier, P.; Chandler, M.; et al. The arthrobacter arilaitensis Re117 genome sequence reveals its genetic adaptation to the surface of cheese. PLoS ONE 2010, 5, e15489. [Google Scholar] [CrossRef]
- Niccum, B.A.; Kastman, E.K.; Kfoury, N.; Robbat Jr., A.; Wolfe, B.E. Strain-level diversity impacts cheese rind microbiome assembly and function. mSystems 2020, 5, e00149-20. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Fouillaud, M.; Dufossé, L. The influence of pH, NaCl, and the deacidifying yeasts Debaryomyces hansenii and Kluyveromyces marxianus on the production of pigments by the cheese-ripening bacteria Arthrobacter arilaitensis. Foods 2018, 7, 190. [Google Scholar] [CrossRef]
- Kaczyński, Ł.K.; Cais-Sokolińska, D.; Bielska, P.; Teichert, J.; Biegalski, J.; Yiğit, A.; Chudy, S. The influence of the texture and color of goat’s salad cheese on the emotional reactions of consumers compared to cow’s milk cheese and Feta cheese. Eur. Food Res. Technol. 2023, 1–16. [Google Scholar] [CrossRef]
- Speight, K.C.; Schiano, A.N.; Harwood, W.S.; Drake, M.A. Consumer insights on prepackaged cheddar cheese shreds using focus groups, conjoint analysis, and qualitative multivariate analysis. J. Dairy Sci. 2019, 102, 6971–6986. [Google Scholar] [CrossRef]
- Giuffrida, D.; Monnet, C.; Laurent, F.; Cacciola, F.; Oteri, M.; Le Piver, M.; Caro, Y.; Donato, P.; Mondello, L.; Roueyre, D.; et al. Carotenoids from the ripening bacterium Brevibacterium linens impart color to the rind of the French cheese, Fourme de Montbrison (PDO). Nat. Prod. Res. 2020, 34, 10–15. [Google Scholar] [CrossRef]
- Possas, A.; Bonilla-Luque, O.M.; Valero, A. From cheese-making to consumption: Exploring the microbial safety of cheeses through predictive microbiology models. Foods 2021, 10, 355. [Google Scholar] [CrossRef] [PubMed]
- Mayo, B.; Rodríguez, J.; Vázquez, L.; Flórez, A.B. Microbial interactions within the cheese ecosystem and their application to improve quality and safety. Foods 2021, 10, 602. [Google Scholar] [CrossRef] [PubMed]
- Gallegos-Acevedo, M.-A.; Chávez-Martíneza, M.; Corral-Luna, A.; Rentería-Monterrubiom, A.-L.; Burrola-Barraza, E.; Lechuga-Valles, R.; Dominguez-Vuveros, J.; Castillo-González, A.-R.; Sánchez-Vega, R. Microbial characterization and diversity of artisanal Ranchero cheese with emphasis in Lactococcus strains. Food Sci. Technol. 2019, 39, 143–148. [Google Scholar] [CrossRef]
- Bockelmann, W. Development of defined surface starter cultures for the ripening of smear cheeses. Int. Dairy J. 2002, 12, 123–131. [Google Scholar] [CrossRef]
- Comi, G.; Cantoni, C. Psychrotrophic Bacteria: Arthrobacter spp. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 372–378. [Google Scholar] [CrossRef]
- Jonnala, B.R.Y.; McSweeney, P.L.H.; Cotter, P.D.; Zhong, S.; Sheehan, J.J.; Kopec, R.E. Comparison of the carotenoid profiles of commonly consumed smear-ripened cheeses. Lebensm. Wiss. Technol. 2021, 135, 110241. [Google Scholar] [CrossRef]
- López, G.D.; Álvarez-Rivera, G.; Carazzone, C.; Ibáñez, E.; Leidy, C.; Cifuentes, A. Bacterial carotenoids: Extraction, characterization, and applications. Crit. Rev. Anal. Chem. 2021, 16, 1–24. [Google Scholar] [CrossRef]
- Meléndez-Martínez, A.J.; Mandić, A.I.; Bantis, F.; Böhm, V.; Borge, G.I.A.; Brnčić, M.; Bysted, A.; Cano, M.P.; Dias, M.G.; Elgersma, A.; et al. A comprehensive review on carotenoids in foods and feeds: Status quo, applications, patents, and research needs. Crit. Rev. Food Sci. Nutr. 2022, 62, 1999–2049. [Google Scholar] [CrossRef]
- Giuffrida, D.; Sutthiwong, S.; Dugo, P.; Donato, P.; Cacciola, F.; Girard-Valenciennes, E.; Le Mao, Y.; Monnet, C.; Fouillaud, M.; Caro, Y.; et al. Characterisation of the C50 carotenoids produced by strains of the cheese-ripening bacterium Arthrobacter arilaitensis. Int. Dairy J. 2016, 55, 10–16. [Google Scholar] [CrossRef]
- Vila, E.; Hornero-Méndez, D.; Azziz, G.; Lareo, C.; Saravia, V. Carotenoids from heterotrophic bacteria isolated from Fildes Peninsula, King George Island, Antarctica. Biotechnol. Rep. 2019, 21, e00306. [Google Scholar] [CrossRef]
- Yabuzaki, J. Carotenoids Database: Structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, bax004. [Google Scholar] [CrossRef]
- Flegler, A.; Lipski, A. The C50 carotenoid bacterioruberin regulates membrane fluidity in pink-pigmented Arthrobacter species. Arch. Microbiol. 2022, 204, 70. [Google Scholar] [CrossRef] [PubMed]
- Siziya, I.N.; Hwang, C.Y.; Seo, M.-J. Antioxidant potential and capacity of microorganism-sourced C30 carotenoids—A review. Antioxidants 2022, 11, 1963. [Google Scholar] [CrossRef] [PubMed]
- Leclercq-Perlat, M.-N.; Sicard, M.; Perrot, N.; Trelea, I.C.; Picque, D.; Corrieu, G. Temperature and relative humidity influence the ripening descriptors of Camembert-type cheeses throughout ripening. J. Dairy Sci. 2015, 98, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Segat, A.; Kelly, A.L.; Sheehan, J.J. Colorants in cheese manufacture: Production, chemistry, interactions, and regulation. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1220–1242. [Google Scholar] [CrossRef]
- Mukhia, S.; Khatri, A.; Acharya, V.; Kumar, R. Comparative genomics and molecular adaptational analysis of Arthrobacter from Sikkim Himalaya provided insights into its survivability under multiple high-altitude stress. Genomics 2021, 113, 151–158. [Google Scholar] [CrossRef]
- Seel, W.; Baust, D.; Sons, D.; Albers, M.; Etzbach, L.; Fuss, J.; Lipski, D. Carotenoids are used as regulators for membrane fluidity by Staphylococcus xylosus. Sci. Rep. 2020, 10, 330. [Google Scholar] [CrossRef]
- Guo, R.; Liu, T.; Guo, C.; Chen, G.; Fan, J.; Zhang, Q. Carotenoid biosynthesis is associated with low-temperature adaptation in Rhodosporidium kratochvilovae. BMC Microbiol. 2022, 22, 319. [Google Scholar] [CrossRef]
- Pérez-Gálvez, A.; Viera, I.; Roca, M. Carotenoids and chlorophylls as antioxidants. Antioxidants 2020, 9, 505. [Google Scholar] [CrossRef]
- Caferri, R.; Guardini, Z.; Bassi, R.; Dall′Osto, L. Assessing photoprotective functions of carotenoids in photosynthetic systems of plants and green algae. In Methods in Enzymology; Wurtzel, E.T., Ed.; Academic Press: Cambridge, MA, USA, 2022; Volume 674, pp. 53–84. [Google Scholar] [CrossRef]
- Chia, G.W.N.; Seviour, T.; Kjelleberg, S.; Hinks, J. Carotenoids improve bacterial tolerance towards biobutanol through membrane stabilization. Environ. Sci. Nano. 2021, 8, 328–341. [Google Scholar] [CrossRef]
- Kumar, S.V.; Taylor, G.; Hasim, S.; Collier, C.P.; Farmer, A.T.; Campagna, S.R.; Bible, A.N.; Doktycz, M.J.; Morrell-Falvey, J. Loss of carotenoids from membranes of Pantoea sp. YR343 results in altered lipid composition and changes in membrane biophysical properties. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1338–1345. [Google Scholar] [CrossRef]
- Manrique-Moreno, M.; Jemioła-Rzemińska, M.; Múnera-Jaramillo, J.; López, G.-D.; Suesca, E.; Leidy, C.; Strzałka, K. Staphylococcus aureus carotenoids modulate the thermotropic phase behavior of model systems that mimic its membrane composition. Membranes 2022, 12, 945. [Google Scholar] [CrossRef] [PubMed]
- Ram, S.; Mitra, M.; Shah, F.; Tirkey, S.R.; Mishra, S. Bacteria as an alternate biofactory for carotenoid production: A review of its applications, opportunities and challenges. J. Funct. Foods 2020, 67, 103867. [Google Scholar] [CrossRef]
- Mostofian, B.; Johnson, Q.R.; Smith, J.C.; Cheng, X. Carotenoids promote lateral packing and condensation of lipid membranes. Phys. Chem. Chem. Phys. 2020, 22, 12281–12293. [Google Scholar] [CrossRef] [PubMed]
- Daly, D.; McSweeney, P.; Sheehan, J. Pink discolouration defect in commercial cheese: A review. Dairy Sci. Technol. 2012, 92, 439–453. [Google Scholar] [CrossRef]
- Jonnala, B.R.Y.; McSweeney, P.L.H.; Cotter, P.D.; Sheehan, J.J. Recreating pink defect in cheese with different strains of Thermus bacteria. Int. J. Dairy Technol. 2021, 74, 700–708. [Google Scholar] [CrossRef]
- Quigley, L.; O’Sullivan, D.J.; Daly, D.; O’Sullivan, O.; Burdikova, Z.; Vana, R.; Beresford, T.P.; Paul Ross, R.; Fitzgerald, G.F.; McSweeney, P.L.H.; et al. Thermus and the pink discoloration defect in cheese. mSystems 2016, 1, e00023-16. [Google Scholar] [CrossRef]
- Cleary, J.L.; Kolachina, S.; Wolfe, B.E.; Sanchez, L.M. Coproporphyrin III produced by the bacterium Glutamicibacter arilaitensis binds zinc and is upregulated by fungi in cheese rinds. mSystems 2018, 3, e00036-18. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Sukdee, P.; Lekhavat, S.; Dufossé, L. Identification of red pigments produced by cheese-ripening bacterial strains of Glutamicibacter arilaitensis using HPLC. Dairy 2021, 2, 396–409. [Google Scholar] [CrossRef]
- Blaya, J.; Barzideh, Z.; LaPointe, G. Symposium review: Interaction of starter cultures and nonstarter lactic acid bacteria in the cheese environment. J. Dairy Sci. 2018, 101, 3611–3629. [Google Scholar] [CrossRef]
- Cao, W.; Aubert, J.; Maillard, M.-B.; Boissel, F.; Leduc, A.; Thomas, J.-L.; Deutsch, S.-M.; Camier, B.; Kerjouh, A.; Parayre, S.; et al. Fine-tuning of process parameters modulates specific metabolic bacterial activities and aroma Compound production in semi-hard cheese. J. Agric. Food Chem. 2021, 69, 8511–8529. [Google Scholar] [CrossRef]
- Coelho, M.C.; Malcata, F.X.; Silva, C.C.G. Lactic acid bacteria in raw-milk cheeses: From starter cultures to probiotic functions. Foods 2022, 11, 2276. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Shi, X.; Wang, B. A review on the general cheese processing technology, flavor biochemical pathways and the influence of yeasts in cheese. Front. Microbiol. 2021, 12, 703284. [Google Scholar] [CrossRef] [PubMed]
- Khattab, A.R.; Guirguis, H.A.; Tawfik, S.T.; Farag, M.A. Cheese ripening: A review on modern technologies towards flavor enhancement, process acceleration and improved quality assessment. Trends. Food Sci. Technol. 2019, 88, 343–360. [Google Scholar] [CrossRef]
- Bockelmann, W.; Hoppe-Seyler, T. The surface flora of bacteria smear-ripened cheese from cow’s and goat’s milk. Int. Dairy J. 2001, 11, 307–314. [Google Scholar] [CrossRef]
- Martín, I.; Rodríguez, A.; García, C.; Córdoba, J.J. Evolution of volatile compounds during ripening and final sensory changes of traditional raw ewe’s milk cheese “Torta del Casar” maturated with selected protective lactic acid bacteria. Foods 2022, 11, 2658. [Google Scholar] [CrossRef]
- Peralta, G.H.; Wolf, I.V.; Perotti, M.C.; Bergamini, C.V.; Hynes, E.R. Formation of volatile compounds, peptidolysis and carbohydrate fermentation by mesophilic lactobacilli and streptoccocci cultures in a cheese extract. Dairy Sci. Technol. 2016, 96, 603–621. [Google Scholar] [CrossRef]
- Lu, Y.; Nawrath, M.; Sun, J.; Liu, S.Q. Effects of physicochemical parameters on volatile sulphur compound formation from L-methionine catabolism by non-growing cells of Kluyveromyces lactis. AMB Expr. 2018, 8, 109. [Google Scholar] [CrossRef]
- Jorge, J.M.; de Silva, M.V.D.M.; Brêda, G.C.; de Souza, C.P.; Leão, R.A.C.; Almeida, R.V.; Bornscheuer, U.T.; de Souza, R.O.M.A. α-Ketobutyrate production under continuous-flow conditions catalyzed by immobilized L-methionine γ-lyase. Eur. J. Org. Chem. 2022, 2022, 165–170. [Google Scholar] [CrossRef]
- Anastasiou, R.; Kazou, M.; Georgalaki, M.; Aktypis, A.; Zoumpopoulou, G.; Tsakalidou, E. Omics approaches to assess flavor development in cheese. Foods 2022, 11, 188. [Google Scholar] [CrossRef]
- Shabnam, W.A. Arthrobacter as biofactory of therapeutic enzymes. Int. J. Pharm. Pharm. Sci. 2018, 10, 1–5. [Google Scholar] [CrossRef]
- Gao, P.; Su, Y.; Zhang, W.; Pang, X.; Xie, N.; Zhang, M.; Lv, J.; Zhang, S. Chemical and flavor characteristics of enzyme-modified cheese made by two-stage processing. Gels 2022, 8, 160. [Google Scholar] [CrossRef] [PubMed]
- Ahn, D.U.; Lee, E.J.; Feng, X.; Zhang, W.; Lee, J.H.; Jo, C.; Nam, K. Mechanisms of volatile production from sulfur-containing amino acids by irradiation. Radiat. Phys. Chem. 2016, 119, 80–84. [Google Scholar] [CrossRef]
- Lee, D.; Jeong, S.; Ahn, J.; Ha, N.-C.; Kwon, A.-R. Crystal structure of bacterial cystathionine Γ-lyase in the cysteine biosynthesis pathway of Staphylococcus aureus. Crystals 2019, 9, 656. [Google Scholar] [CrossRef]
- Sen, U.; Givvimani, S.; Abe, O.A.; Lederer, E.D.; Tyagi, S.C. Cystathionine β-synthase and cystathionine γ-lyase double gene transfer ameliorate homocysteine-mediated mesangial inflammation through hydrogen sulfide generation. Am. J. Physiol. Cell Physiol. 2011, 300, 155–163. [Google Scholar] [CrossRef]
- Sutthiwong, N.; Dufossé, L. Production of carotenoids by Arthrobacter arilaitensis strains isolated from smear-ripened cheeses. FEMS Microbiol. Lett. 2014, 360, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Rutkiewicz, M.; Bujacz, A.; Wanarska, M.; Wierzbicka-Wos, A.; Cieslinski, H. Active site architecture and reaction mechanism determination of cold adapted β-d-galactosidase from Arthrobacter sp. 32cB. Int. J. Mol. Sci. 2019, 20, 4301. [Google Scholar] [CrossRef]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of β-galactosidase and its applications in food industry. 3 Biotech 2017, 7, 79. [Google Scholar] [CrossRef]
- Vera, C.; Guerrero, C.; Aburto, C.; Cordova, A.; Illanes, A. Conventional and non-conventional applications of β-galactosidases. Biochim. Biophys. Acta Proteins. Proteom. 2020, 1868, 140271. [Google Scholar] [CrossRef]
- Pawlak-Szukalska, A.; Wanarska, M.; Popinigis, A.T.; Kur, J. A novel cold-active β-d-galactosidase with transglycosylation activity from the Antarctic Arthrobacter sp. 32cB–Gene cloning, purification and characterization. Process Biochem. 2014, 49, 2122–2133. [Google Scholar] [CrossRef]
- De Albuquerque, T.L.; de Sousa, M.; de Silva, N.C.G.; Neto, C.A.C.G.; Gonçalves, L.R.B.; Fernandez-Lafuente, R.; Rocha, M.V.P. β-galactosidase from Kluyveromyces lactis: Characterization, production, immobilization and applications-A review. Int. J. Biol. Macromol. 2021, 191, 881–898. [Google Scholar] [CrossRef]
- De Sousa, C.C.; de Resende, M.M.; Falleiros, L.N.S.S.; Ribeiro, E.J. Synthesis and immobilization of β-galactosidase from Kluyveromyces marxianus using ion exchange resin. Ind. Biotechnol. 2021, 17, 27–37. [Google Scholar] [CrossRef]
- Otieno, D.O. Synthesis of β-alactooligosaccharides from lactose using microbial β-galactosidases. Compr. Rev. Food Sci. Food Saf. 2010, 9, 471–482. [Google Scholar] [CrossRef]
- Fai, A.E.C.; Silva, J.B.; Andrade, C.J.; Bution, M.L.; Pastore, G.M. Production of prebiotic galactooligosaccharides from lactose by Pseudozyma tsukubaensis and Pichia kluyveri. Biocatal. Agric. Biotechnol. 2014, 3, 343–350. [Google Scholar] [CrossRef]
- Shi, X.; Wu, D.; Xu, Y.; Yu, X. Engineering the optimum pH of β-galactosidase from Aspergillus oryzae for efficient hydrolysis of lactose. J. Dairy Sci. 2022, 105, 4772–4782. [Google Scholar] [CrossRef]
- Montilla, A.; Megías-Pérez, R.; Olano, A.; Villamiel, M. Presence of galactooligosaccharides and furosine in special dairy products designed for elderly people. Food Chem. 2015, 172, 481–485. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, A.L.; Rodrigues, L.R.; Lima, N.M.; Teixeira, J.A. An Overview of the recent developments on fructooligosaccharide production and applications. Food Bioprocess Technol. 2014, 7, 324–337. [Google Scholar] [CrossRef]
- Botvynko, A.; Bednářová, A.; Henke, S.; Shakhno, N.; Čurda, L. Production of galactooligosaccharides using various combinations of the commercial β-galactosidases. Biochem. Biophys. Res. Commun. 2019, 517, 762–766. [Google Scholar] [CrossRef]
- Duarte, A.W.F.; Dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Wentzel, L.C.P.; Lario, L.D.; Rodrigues, A.; Pagnocca, F.C.; et al. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef]
- Sarmiento, F.; Peralta, R.; Blamey, J.M. Cold and hot extremozymes: Industrial relevance and current trends. Front. Bioeng. Biotechnol. 2015, 3, 148. [Google Scholar] [CrossRef]
- Xia, Y.; He, L.; Mao, J.; Fang, P.; Ma, X.; Wang, Z. Purification, characterization, and gene cloning of a new cold-adapted β-galactosidase from Erwinia sp. E602 isolated in northeast China. J. Dairy Sci. 2018, 1, 6946–6954. [Google Scholar] [CrossRef]
- Zolnere, K.; Ciprovica, I. The comparison of commercially available β-galactosidase for dairy industry: Review. Res. Rural. Dev. 2017, 1, 215–222. [Google Scholar] [CrossRef]
- Mangiagalli, M.; Lotti, M. Cold-active β-galactosidases: Insight into cold adaption mechanisms and biotechnological exploitation. Mar. Drugs 2021, 19, 43. [Google Scholar] [CrossRef]
- Rutkiewicz, M.; Wanarska, M.; Bujacz, A. Mapping the transglycosylation relevant sites of cold-adapted β-d-galactosidase from Arthrobacter sp. 32cB. Int. J. Mol. Sci. 2020, 21, 5354. [Google Scholar] [CrossRef] [PubMed]
- Hamid, B.; Bashir, Z.; Yatoo, A.M.; Mohiddin, F.; Majeed, N.; Bansal, M.; Poczai, P.; Almalki, W.H.; Sayyed, R.Z.; Shati, A.A.; et al. Cold-active enzymes and their potential industrial applications—A Review. Molecules 2022, 27, 5885. [Google Scholar] [CrossRef] [PubMed]
- Luan, S.; Duan, X.A. Novel thermal-activated β-galactosidase from Bacillus aryabhattai GEL-09 for lactose hydrolysis in milk. Foods 2022, 11, 372. [Google Scholar] [CrossRef]
- Nakagawa, T.; Fujimoto, Y.; Ikehata, R.; Miyaji, T.; Tomizuka, N. Purification and molecular characterization of cold-active beta-galactosidase from Arthrobacter psychrolactophilus strain F2. Appl. Microbiol. Biotechnol. 2006, 72, 720–725. [Google Scholar] [CrossRef] [PubMed]
- Panesar, P.S.; Bali, V. Prebiotics. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Academic Press: Cambridge, MA, USA, 2016; pp. 464–471. [Google Scholar] [CrossRef]
- De Santi, C.; Altermark, B.; de Pascale, D.; Willassen, N.P. Bioprospecting around Arctic islands: Marine bacteria as rich source of biocatalysts. J. Basic Microbiol. 2016, 56, 238–253. [Google Scholar] [CrossRef] [PubMed]
- Díez-Municio, M.; Herrero, M.; Olano, A.; Moreno, F.J. Synthesis of novel bioactive lactose-derived oligosaccharides by microbial glycoside hydrolases. Microb. Biotechnol. 2014, 7, 315–331. [Google Scholar] [CrossRef]
- Xavier, J.R.; Ramana, K.V.; Sharma, R.K. β-galactosidase: Biotechnological applications in food processing. J. Food Biochem. 2018, 42, e12564. [Google Scholar] [CrossRef]
- Dekker, P.J.T. Enzyme exogenous to milk in dairy technology: β-d-galactosidases. In Encyclopedia of Dairy Sciences, 2nd ed.; Fuquay, J.W., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 276–283. [Google Scholar]
- Dekker, P.J.T.; Koenders, D.; Bruins, M.J. Lactose-free dairy products: Market developments, production, nutrition and health benefits. Nutrients 2019, 11, 551. [Google Scholar] [CrossRef]
- Alshanberi, A.M.; Al-Shaeri, M.A.; Ansari, S.A. Overviewing the application of β-galactosidase “Immobilized on nanoparticles” in dairy industries. Braz. Arch. Biol. Technol. 2021, 64, 1–20. [Google Scholar] [CrossRef]
- Orrego, D.; Klotz-Ceberio, B. Enzymatic synthesis of galacto-oligosaccharides from concentrated sweet whey permeate and its application in a dairy product. Appl. Sci. 2022, 12, 10229. [Google Scholar] [CrossRef]
- Nayik, G.A.; Jagdale, Y.D.; Gaikwad, S.A.; Devkatte, A.N.; Dar, A.H.; Ansari, M.J. Nutritional profile, processing and potential aroducts: A comparative review of goat milk. Dairy 2022, 3, 622–647. [Google Scholar] [CrossRef]
- Crovadore, J.; Grizard, D.; Chablais, R.; Cochard, B.; Blanc, P.; Lefort, F. Whole-genome sequences of two Arthrobacter sp. strains, 4041 and 4042, potentially usable in agriculture and environmental depollution. Microbiol. Resour. Announc. 2018, 7, e01054-18. [Google Scholar] [CrossRef]
- Niewerth, H.; Schuldes, J.; Parschat, K.; Kiefer, P.; Vorholt, J.A.; Daniel, R.; Fetzner, S. Complete genome sequence and metabolic potential of the quinaldine-degrading bacterium Arthrobacter sp. Rue61a. BMC Genom. 2012, 13, 534. [Google Scholar] [CrossRef]
- Han, S.R.; Kim, B.; Jang, J.H.; Park, H.; Oh, T.J. Complete genome sequence of Arthrobacter sp. PAMC25564 and its comparative genome analysis for elucidating the role of CAZymes in cold adaptation. BMC Genom. 2021, 22, 403. [Google Scholar] [CrossRef]
- Chen, C.; Deng, J.; Lv, X.; Li, J.; Du, G.; Li, H.; Liu, L. Biocatalytic synthesis of lactosucrose using a recombinant thermostable β-fructofuranosidase from Arthrobacter sp. 10138. Bioengineered 2020, 1, 416–427. [Google Scholar] [CrossRef]
- Solihin, J.; Waturangi, D.E.; Purwadaria, T. Induction of amylase and protease as antibiofilm agents by starch, casein, and yeast extract in Arthrobacter sp. CW01. BMC Microbiol. 2021, 21, 232. [Google Scholar] [CrossRef]
- McAuliffe, O.; Kilcawley, K. Symposium review: Genomic investigations of flavor formation by dairy microbiota. J. Dairy Sci. 2019, 102, 909–922. [Google Scholar] [CrossRef]
- Kieliszek, M.; Pobiega, K.; Piwowarek, K.; Kot, A.M. Characteristics of the proteolytic enzymes produced by lactic acid bacteria. Molecules 2021, 26, 1858. [Google Scholar] [CrossRef]
- Gabriel, H.-F.; Beatriz, G.; Manuel, C.; Laura, C.; Luis, G.J. Transcriptional response of the xerotolerant Arthrobacter sp. Helios strain to PEG-induced drought stress. Front. Microbiol. 2022, 13, 1–16. [Google Scholar] [CrossRef]
- Marcone, S.; Belton, O.; Fitzgerald, D.J. Milk-derived bioactive peptides and their health promoting effects: A potential role in atherosclerosis. Br. J. Clin. Pharmacol. 2017, 83, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Adams, C.; Sawh, F.; Green-Johnson, J.M.; Jones Taggart, H.; Strap, J.L. Characterization of casein-derived peptide bioactivity: Differential effects on angiotensin-converting enzyme inhibition and cytokine and nitric oxide production. J. Dairy Sci. 2020, 103, 5805–5815. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Youn, B. Hypolipidemic roles of casein-derived peptides by regulation of trans-intestinal cholesterol excretion and bile acid synthesis. Nutrients 2020, 12, 3058. [Google Scholar] [CrossRef] [PubMed]
- Samtiya, M.; Samtiya, S.; Badgujar, P.C.; Puniya, A.K.; Dhewa, T.; Aluko, R.E. Health-promoting and therapeutic attributes of milk-derived bioactive peptides. Nutrients 2022, 14, 3001. [Google Scholar] [CrossRef] [PubMed]
- Akarca, G. Lipolysis and aroma occurrence in Erzincan Tulum cheese, which is produced by adding probiotic bacteria and ripened in various packages. Food Sci. Technol. 2020, 40, 102–116. [Google Scholar] [CrossRef]
- Nomura, S.O.; Karger, A.B.; Weir, N.L.; Duprez, D.A.; Tsai, M.Y. Free fatty acids, cardiovascular disease, and mortality in the multi-ethnic study of atherosclerosis. J. Clin. Lipidol. 2020, 14, 531–541. [Google Scholar] [CrossRef]
- Xiong, R.G.; Zhou, D.D.; Wu, S.X.; Huang, S.Y.; Saimaiti, A.; Yang, Z.J.; Shang, A.; Zhao, C.N.; Gan, R.Y.; Li, H.B. Health benefits and side effects of short-chain fatty acids. Foods 2022, 11, 2863. [Google Scholar] [CrossRef]
- Huang, L.; Gao, L.; Chen, C. Role of medium-chain fatty acids in healthy metabolism: A clinical perspective. Trends. Endocrinol. Metab. 2021, 32, 351–366. [Google Scholar] [CrossRef]
- Ibrahim, A.H. Impact of hydrolyzed lactose by β-galactosidase enzyme on the physicochemical and organoleptic properties of fermented camel milk. Emir. J. Food Agric. 2018, 30, 778–790. [Google Scholar] [CrossRef]
- Bintsis, T. Lactic acid bacteria as starter cultures: An update in their metabolism and genetics. AIMS Microbiol. 2018, 4, 665–684. [Google Scholar] [CrossRef] [PubMed]
- Dominici, S.; Marescotti, F.; Sanmartin, C.; Macaluso, M.; Taglieri, I.; Venturi, F.; Zinnai, A.; Facioni, M.S. Lactose: Characteristics, food and drug-related applications, and its possible substitutions in meeting the needs of people with lactose intolerance. Foods 2022, 11, 1486. [Google Scholar] [CrossRef] [PubMed]
- Pachekrepapol, U.; Somboonchai, N.; Krimjai, W. Physicochemical, rheological, and microbiological properties of lactose-free functional yogurt supplemented with fructooligosaccharides. J. Food Process Preserv. 2020, 45, 404–419. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sutthiwong, N.; Lekavat, S.; Dufossé, L. Involvement of Versatile Bacteria Belonging to the Genus Arthrobacter in Milk and Dairy Products. Foods 2023, 12, 1270. https://doi.org/10.3390/foods12061270
Sutthiwong N, Lekavat S, Dufossé L. Involvement of Versatile Bacteria Belonging to the Genus Arthrobacter in Milk and Dairy Products. Foods. 2023; 12(6):1270. https://doi.org/10.3390/foods12061270
Chicago/Turabian StyleSutthiwong, Nuthathai, Supaporn Lekavat, and Laurent Dufossé. 2023. "Involvement of Versatile Bacteria Belonging to the Genus Arthrobacter in Milk and Dairy Products" Foods 12, no. 6: 1270. https://doi.org/10.3390/foods12061270
APA StyleSutthiwong, N., Lekavat, S., & Dufossé, L. (2023). Involvement of Versatile Bacteria Belonging to the Genus Arthrobacter in Milk and Dairy Products. Foods, 12(6), 1270. https://doi.org/10.3390/foods12061270