
Analysis of the Differentially Expressed Proteins in Donkey Milk in Different Lactation Stages


Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Sample Collection and Preparation
2.2. Total Protein Extraction
2.3. Trypsin Treatment
2.4. DDA Spectrum Library Construction
2.4.1. Separation of Fractions
2.4.2. LC-MS/MS Analysis
2.5. Data Analysis
2.5.1. Identification and Quantitation of Protein
2.5.2. Functional Analysis of Proteins
3. Results
3.1. Characterization of Proteins in DM
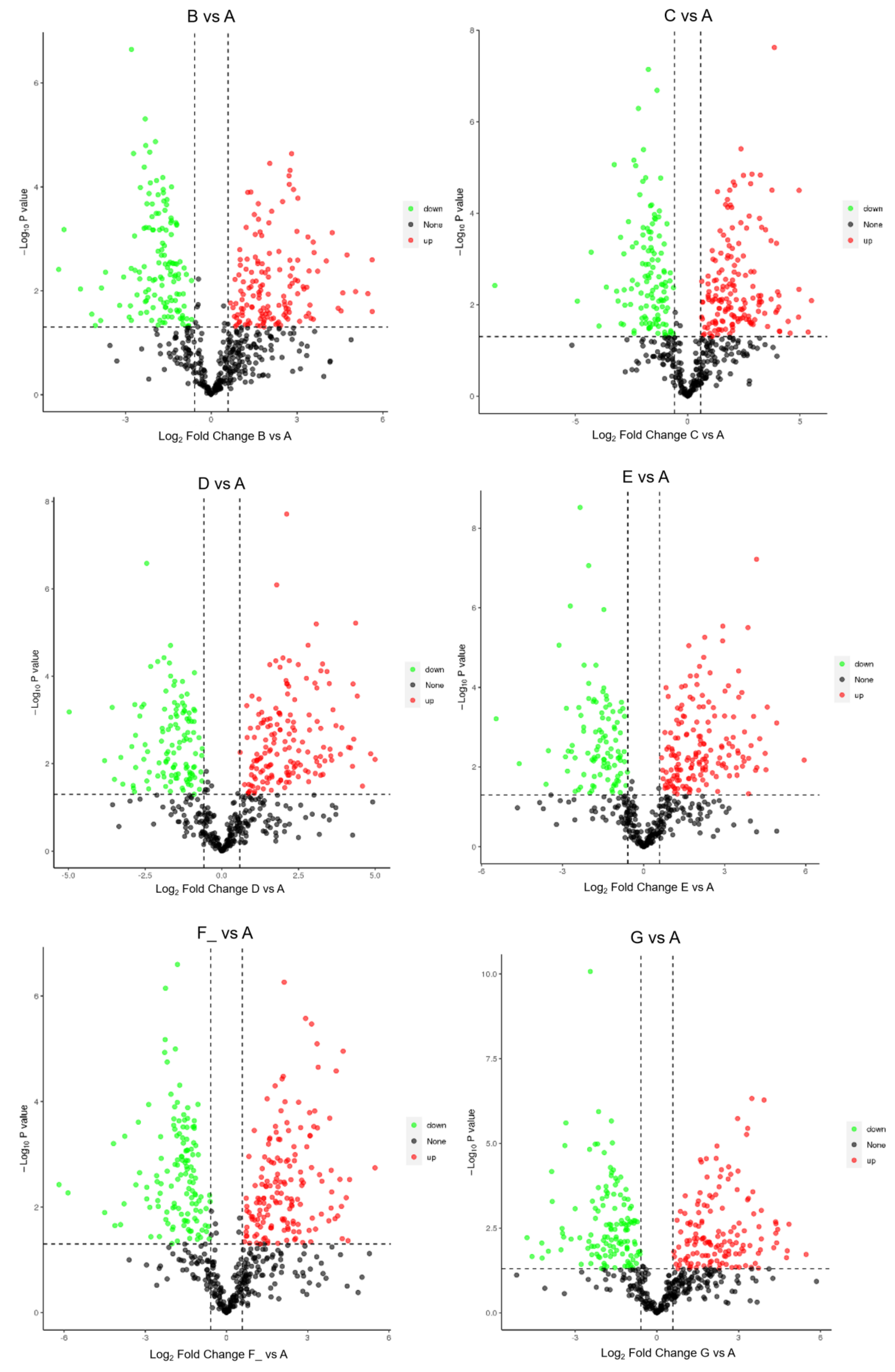
3.2. Identification of DEPs in Different Lactation Stages
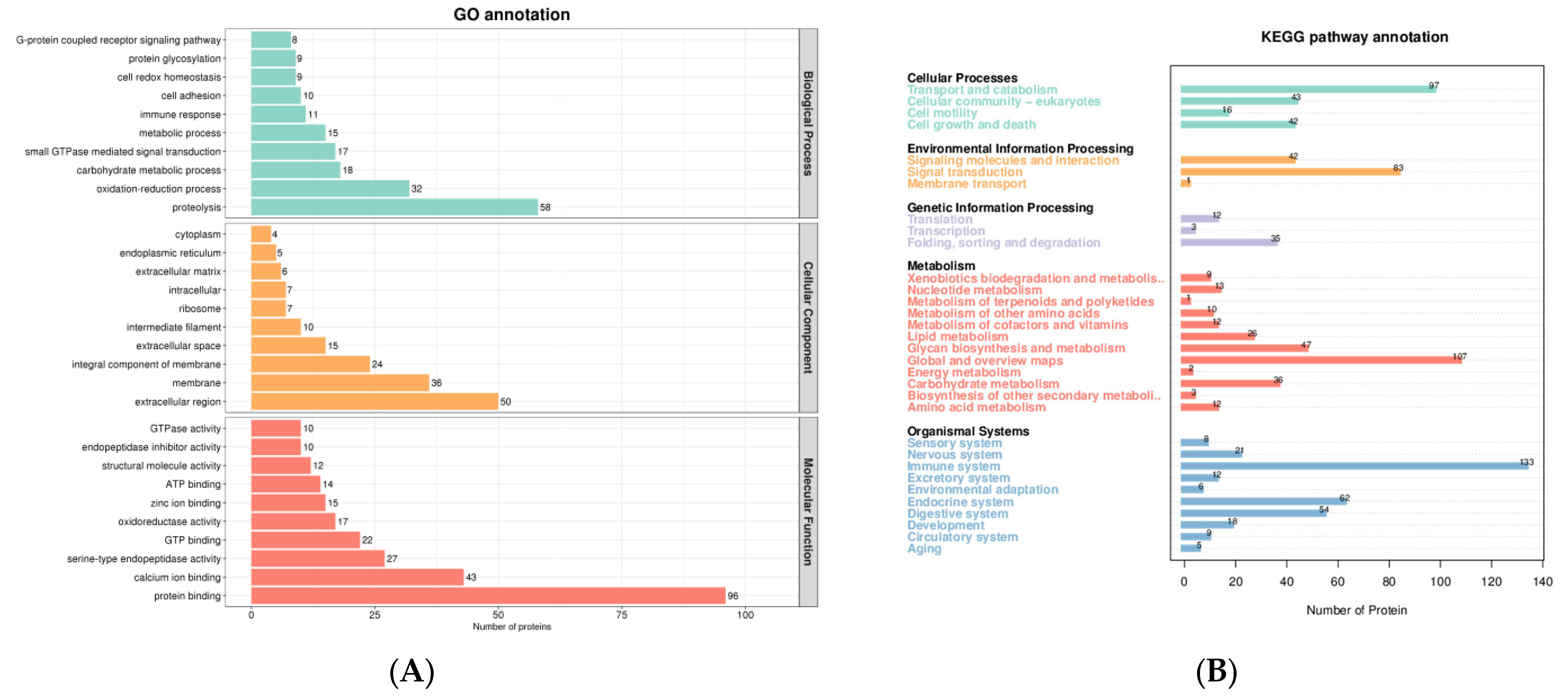
3.3. GO Analysis of DEPs
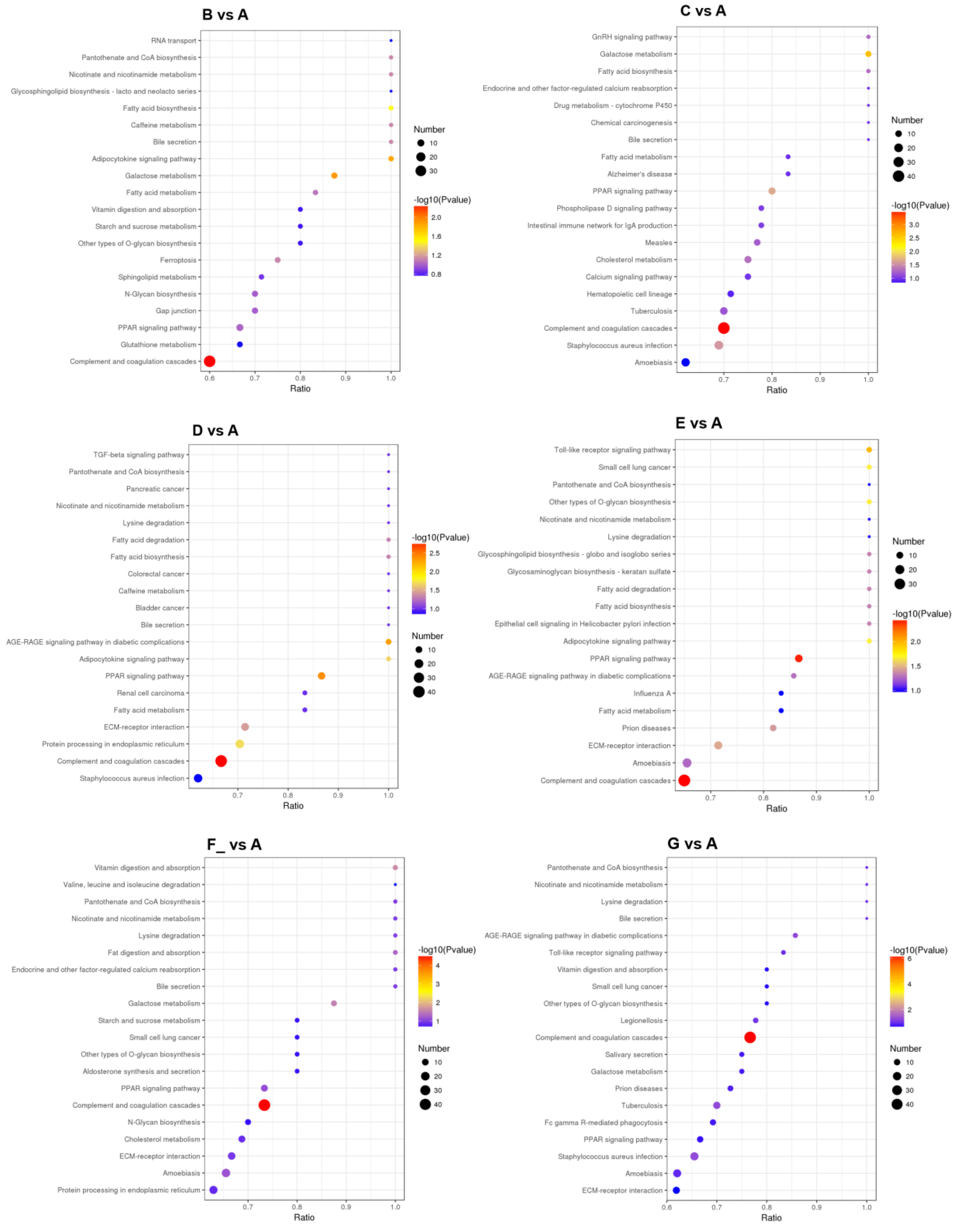
3.4. Metabolic Pathway Analysis of DEPs
4. Discussion
4.1. Nutrition and Transportation
4.2. Cell Proliferation and Differentiation
4.3. Antibacterial and Anti-Inflammatory Activity
4.4. Cosmetics
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Guo, H.Y.; Pang, K.; Zhang, X.Y.; Zhao, L.; Chen, S.W.; Dong, M.L.; Ren, F.Z. Composition, physiochemical properties, nitrogen fraction distribution, and amino acid profile of donkey milk. J. Dairy Sci. 2007, 90, 1635–1643. [Google Scholar] [CrossRef] [PubMed]
- Aspri, M.; Leni, G.; Galaverna, G.; Papademas, P. Bioactive properties of fermented donkey milk, before and after in vitro simulated gastrointestinal digestion. Food Chem. 2018, 268, 476–484. [Google Scholar] [CrossRef]
- Vincenzetti, S.; Santini, G.; Polzonetti, V.; Pucciarelli, S.; Klimanova, Y.; Polidori, P. Vitamins in Human and Donkey Milk: Functional and Nutritional Role. Nutrients 2021, 13, 1509. [Google Scholar] [CrossRef] [PubMed]
- Papademas, P.; Kamilari, E.; Aspri, M.; Anagnostopoulos, D.A.; Mousikos, P.; Kamilaris, A.; Tsaltas, D. Investigation of donkey milk bacterial diversity by 16S rDNA high-throughput sequencing on a Cyprus donkey farm. J. Dairy Sci. 2021, 104, 167–178. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ma, Q.; Liu, G.; Wang, C. Effects of donkey milk on oxidative stress and inflammatory response. J. Food Biochem. 2022, 46, e13935. [Google Scholar] [CrossRef] [PubMed]
- Spada, V.; Ferranti, P.; Chianese, L.; Salimei, E.; Addeo, F.; Picariello, G. Antibacterial potential of donkey’s milk disclosed by untargeted proteomics. J. Proteom. 2021, 231, 104007. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Fan, Y.; Shaikh, A.S.; Wang, Z.; Wang, D.; Tan, H. Dezhou donkey (Equus asinus) milk a potential treatment strategy for type 2 diabetes. J. Ethnopharmacol. 2020, 246, 112221. [Google Scholar] [CrossRef]
- Li, Q.; Li, M.; Zhang, J.; Shi, X.; Yang, M.; Zheng, Y.; Cao, X.; Yue, X.; Ma, S. Donkey milk inhibits triple-negative breast tumor progression and is associated with increased cleaved-caspase-3 expression. Food Funct. 2020, 11, 3053–3065. [Google Scholar] [CrossRef]
- Kocic, H.; Langerholc, T.; Kostic, M.; Stojanovic, S.; Najman, S.; Krstic, M.; Nesic, I.; Godic, A.; Wollina, U. The Regenerative Potential of Donkey and Human Milk on the Redox-Sensitive and Proliferative Signaling Pathways of Skin Fibroblasts. Oxid. Med. Cell. Longev. 2020, 2020, 5618127. [Google Scholar] [CrossRef]
- Jones, P.; Binns, D.; Chang, H.Y.; Fraser, M.; Li, W.; McAnulla, C.; McWilliam, H.; Maslen, J.; Mitchell, A.; Nuka, G.; et al. InterProScan 5: Genome-scale protein function classification. Bioinformatics 2014, 30, 1236–1240. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic Acids Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Cunsolo, V.; Saletti, R.; Muccilli, V.; Gallina, S.; Di Francesco, A.; Foti, S. Proteins and bioactive peptides from donkey milk: The molecular basis for its reduced allergenic properties. Food Res. Int. 2017, 99 Pt 1, 41–57. [Google Scholar] [CrossRef]
- Valle, E.; Pozzo, L.; Giribaldi, M.; Bergero, D.; Gennero, M.S.; Dezzutto, D.; McLean, A.; Borreani, G.; Coppa, M.; Cavallarin, L. Effect of farming system on donkey milk composition. J. Sci. Food Agric. 2018, 98, 2801–2808. [Google Scholar] [CrossRef] [PubMed]
- Raspa, F.; Cavallarin, L.; McLean, A.K.; Bergero, D.; Valle, E. A Review of the Appropriate Nutrition Welfare Criteria of Dairy Donkeys: Nutritional Requirements, Farm Management Requirements and Animal-Based Indicators. Animals 2019, 9, 315. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, C.; Paolino, R.; Freschi, P.; Calluso, A.M. Short communication: Jenny milk production and qualitative characteristics. J. Dairy Sci. 2012, 95, 2910–2915. [Google Scholar] [CrossRef] [PubMed]
- Martini, M.; Altomonte, I.; Salari, F.; Caroli, A.M. Short communication: Monitoring nutritional quality of Amiata donkey milk: Effects of lactation and productive season. J. Dairy Sci. 2014, 97, 6819–6822. [Google Scholar] [CrossRef]
- Malacarne, M.; Criscione, A.; Franceschi, P.; Bordonaro, S.; Formaggioni, P.; Marletta, D.; Summer, A. New Insights into Chemical and Mineral Composition of Donkey Milk throughout Nine Months of Lactation. Animals 2019, 9, 1161. [Google Scholar] [CrossRef]
- Bradshaw, C.V.; Suarez Trujillo, A.; Luecke, S.M.; Logan, L.D.; Mohallem, R.; Aryal, U.K.; Stewart, K.R.; Casey, T.M.; Minor, R.C. Shotgun proteomics of homogenate milk reveals dynamic changes in protein abundances between colostrum, transitional, and mature milk of swine. J. Anim. Sci. 2021, 99, skab240. [Google Scholar] [CrossRef]
- Li, M.; Kang, S.; Zheng, Y.; Shao, J.; Zhao, H.; An, Y.; Cao, G.; Li, Q.; Yue, X.; Yang, M. Comparative metabolomics analysis of donkey colostrum and mature milk using ultra-high-performance liquid tandem chromatography quadrupole time-of-flight mass spectrometry. J. Dairy Sci. 2020, 103, 992–1001. [Google Scholar] [CrossRef]
- Young, P.A.; Senkal, C.E.; Suchanek, A.L.; Grevengoed, T.J.; Lin, D.D.; Zhao, L.; Crunk, A.E.; Klett, E.L.; Fullekrug, J.; Obeid, L.M.; et al. Long-chain acyl-CoA synthetase 1 interacts with key proteins that activate and direct fatty acids into niche hepatic pathways. J. Biol. Chem. 2018, 293, 16724–16740. [Google Scholar] [CrossRef]
- Huh, J.Y.; Reilly, S.M.; Abu-Odeh, M.; Murphy, A.N.; Mahata, S.K.; Zhang, J.; Cho, Y.; Seo, J.B.; Hung, C.W.; Green, C.R.; et al. TANK-Binding Kinase 1 Regulates the Localization of Acyl-CoA Synthetase ACSL1 to Control Hepatic Fatty Acid Oxidation. Cell Metab. 2020, 32, 1012–1027.e7. [Google Scholar] [CrossRef] [PubMed]
- Monks, J.; Ladinsky, M.S.; McManaman, J.L. Organellar Contacts of Milk Lipid Droplets. Contact 2020, 3, 2515256419897226. [Google Scholar] [CrossRef] [PubMed]
- Monks, J.; Orlicky, D.J.; Libby, A.E.; Dzieciatkowska, M.; Ladinsky, M.S.; McManaman, J.L. Perilipin-2 promotes lipid droplet-plasma membrane interactions that facilitate apocrine lipid secretion in secretory epithelial cells of the mouse mammary gland. Front. Cell Dev. Biol. 2022, 10, 958566. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.; Snider, J.M.; Cariello, C.P.; Lambert, J.M.; Anderson, A.K.; Cowart, L.A.; Snider, A.J. Sphingosine kinase 1 is required for myristate-induced TNFalpha expression in intestinal epithelial cells. Prostaglandins Other Lipid Mediat. 2020, 149, 106423. [Google Scholar] [CrossRef]
- Carr, J.M.; Mahalingam, S.; Bonder, C.S.; Pitson, S.M. Sphingosine kinase 1 in viral infections. Rev. Med. Virol. 2013, 23, 73–84. [Google Scholar] [CrossRef] [PubMed]
- Ceballos-Marquez, A.; Barkema, H.W.; Stryhn, H.; Wichtel, J.J.; Neumann, J.; Mella, A.; Kruze, J.; Espindola, M.S.; Wittwer, F. The effect of selenium supplementation before calving on early-lactation udder health in pastured dairy heifers. J. Dairy Sci. 2010, 93, 4602–4612. [Google Scholar] [CrossRef] [PubMed]
- Arshad, M.A.; Ebeid, H.M.; Hassan, F.U. Revisiting the Effects of Different Dietary Sources of Selenium on the Health and Performance of Dairy Animals: A Review. Biol. Trace Elem. Res. 2021, 199, 3319–3337. [Google Scholar] [CrossRef]
- Lemly, A.D. Symptoms and implications of selenium toxicity in fish: The Belews Lake case example. Aquat. Toxicol. 2002, 57, 39–49. [Google Scholar] [CrossRef]
- Pieszka, M.; Bederska-Lojewska, D.; Szczurek, P.; Pieszka, M. The Membrane Interactions of Nano-Silica and Its Potential Application in Animal Nutrition. Animals 2019, 9, 1041. [Google Scholar] [CrossRef]
- Zhang, F.; Pu, J.; Gu, Z.; Mao, H. DIA proteomics reveals hypotensive and immune-enhancing constituents in buffalo whey from different altitudes. Int. J. Biol. Macromol. 2020, 164, 4146–4154. [Google Scholar] [CrossRef]
- Albertos, I.; Lopez, M.; Jimenez, J.M.; Cao, M.J.; Corell, A.; Castro-Alija, M.J. Characterisation of Zamorano-Leonese Donkey Milk as an Alternative Sustainably Produced Protein Food. Front. Nutr. 2022, 9, 872409. [Google Scholar] [CrossRef] [PubMed]
- Zivkov Balos, M.; Ljubojevic Pelic, D.; Jaksic, S.; Lazic, S. Donkey Milk: An Overview of its Chemical Composition and Main Nutritional Properties or Human Health Benefit Properties. J. Equine Vet. Sci. 2023, 121, 104225. [Google Scholar] [CrossRef] [PubMed]
- Fedosov, S.N.; Nexo, E.; Heegaard, C.W. Vitamin B(12) and its binding proteins in milk from cow and buffalo in relation to bioavailability of B(12). J. Dairy Sci. 2019, 102, 4891–4905. [Google Scholar] [CrossRef] [PubMed]
- Hine, B.; Boggs, I.; Green, R.; Miller, J.W.; Hovey, R.C.; Humphrey, R.; Wheeler, T.T. Transcobalamin derived from bovine milk stimulates apical uptake of vitamin B12 into human intestinal epithelial cells. J. Cell. Biochem. 2014, 115, 1948–1954. [Google Scholar] [CrossRef] [PubMed]
- Naik, S.; Bhide, V.; Babhulkar, A.; Mahalle, N.; Parab, S.; Thakre, R.; Kulkarni, M. Daily milk intake improves vitamin B-12 status in young vegetarian Indians: An intervention trial. Nutr. J. 2013, 12, 136. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, X.; Ji, X.; Jin, Y.; Liu, W. The amino acid transporter PAT1 regulates mTORC1 in a nutrient-sensitive manner that requires its transport activity. Cell Signal. 2019, 53, 59–67. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Wan, X.; Du, X.; Zhong, Z.; Peng, J.; Xiong, Q.; Chai, J.; Jiang, S. Insights into the Interaction of Lysosomal Amino Acid Transporters SLC38A9 and SLC36A1 Involved in mTORC1 Signaling in C2C12 Cells. Biomolecules 2021, 11, 1314. [Google Scholar] [CrossRef]
- Billings, P.C.; Whitbeck, J.C.; Adams, C.S.; Abrams, W.R.; Cohen, A.J.; Engelsberg, B.N.; Howard, P.S.; Rosenbloom, J. The transforming growth factor-beta-inducible matrix protein (beta)ig-h3 interacts with fibronectin. J. Biol. Chem. 2002, 277, 28003–28009. [Google Scholar] [CrossRef]
- Kim, J.E.; Kim, S.J.; Jeong, H.W.; Lee, B.H.; Choi, J.Y.; Park, R.W.; Park, J.Y.; Kim, I.S. RGD peptides released from beta ig-h3, a TGF-beta-induced cell-adhesive molecule, mediate apoptosis. Oncogene 2003, 22, 2045–2053. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, P.K.; Bae, S.M.; Son, H.N.; Thoudam, D.S.; Kim, J.E.; Lee, B.H.; Park, R.W.; Kim, I.S. Transforming growth factor-beta-induced protein (TGFBIp/beta ig-h3) activates platelets and promotes thrombogenesis. Blood 2009, 114, 5206–5215. [Google Scholar] [CrossRef]
- Sia, G.M.; Clem, R.L.; Huganir, R.L. The human language-associated gene SRPX2 regulates synapse formation and vocalization in mice. Science 2013, 342, 987–991. [Google Scholar] [CrossRef] [PubMed]
- Yim, A.; Smith, C.; Brown, A.M. Osteopontin/secreted phosphoprotein-1 harnesses glial-, immune-, and neuronal cell ligand-receptor interactions to sense and regulate acute and chronic neuroinflammation. Immunol. Rev. 2022, 311, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Takeda, T.; Sakata, M.; Minekawa, R.; Yamamoto, T.; Hayashi, M.; Tasaka, K.; Murata, Y. Human milk induces fetal small intestinal cell proliferation—Involvement of a different tyrosine kinase signaling pathway from epidermal growth factor receptor. J. Endocrinol. 2004, 181, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Howangyin, K.Y.; Zlatanova, I.; Pinto, C.; Ngkelo, A.; Cochain, C.; Rouanet, M.; Vilar, J.; Lemitre, M.; Stockmann, C.; Fleischmann, B.K.; et al. Myeloid-Epithelial-Reproductive Receptor Tyrosine Kinase and Milk Fat Globule Epidermal Growth Factor 8 Coordinately Improve Remodeling After Myocardial Infarction via Local Delivery of Vascular Endothelial Growth Factor. Circulation 2016, 133, 826–839. [Google Scholar] [CrossRef] [PubMed]
- Ajith Kumar, A.; Siva Kumar, N. Biochemical Characterization of a Lysosomal alpha-Mannosidase from the Starfish Asterias rubens. Protein J. 2018, 37, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Paciotti, S.; Codini, M.; Tasegian, A.; Ceccarini, M.R.; Cataldi, S.; Arcuri, C.; Fioretti, B.; Albi, E.; Beccari, T. Lysosomal alpha-mannosidase and alpha-mannosidosis. Front. Biosci. 2017, 22, 157–167. [Google Scholar]
- Abbondante, S.; Pearlman, E. Breaching bacterial biofilm with neutrophil alpha-mannosidase. J. Leukoc. Biol. 2019, 105, 1085. [Google Scholar] [CrossRef]
- Zito, E. PRDX4, an endoplasmic reticulum-localized peroxiredoxin at the crossroads between enzymatic oxidative protein folding and nonenzymatic protein oxidation. Antioxid. Redox Signal. 2013, 18, 1666–1674. [Google Scholar] [CrossRef]
- Liang, X.; Yan, Z.; Ma, W.; Qian, Y.; Zou, X.; Cui, Y.; Liu, J.; Meng, Y. Peroxiredoxin 4 protects against ovarian ageing by ameliorating D-galactose-induced oxidative damage in mice. Cell Death Dis. 2020, 11, 1053. [Google Scholar] [CrossRef]
- Yamada, S.; Ding, Y.; Sasaguri, Y. Peroxiredoxin 4: Critical roles in inflammatory diseases. J. UOEH 2012, 34, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Mort, J.S.; Buttle, D.J. Cathepsin B. Int. J. Biochem. Cell Biol. 1997, 29, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Gaire, B.P.; Subedi, L.; Teramoto, H.; Hu, B. The Role of Cathepsin B in Ischemia-Reperfusion Injury after Stroke. In Cerebral Ischemia; Pluta, R., Ed.; Exon Publications: Brisbane, Australia, 2021. [Google Scholar]
- Yvon, S.; Schwebel, L.; Belahcen, L.; Tormo, H.; Peter, M.; Haimoud-Lekhal, D.A.; Eutamene, H.; Jard, G. Effects of thermized donkey milk with lysozyme activity on altered gut barrier in mice exposed to water-avoidance stress. J. Dairy Sci. 2019, 102, 7697–7706. [Google Scholar] [CrossRef] [PubMed]
- Nagai, T.; Suzuki, N.; Nagashima, T. Collagen from common minke whale (Balaenoptera acutorostrata) unesu. Food Chem. 2008, 111, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; He, H.; Chen, Y.; Liao, F.; Tang, J.; Li, L.; Fan, Y.; Li, L.; Xiong, L. Effects of donkey milk on UVB-induced skin barrier damage and melanin pigmentation: A network pharmacology and experimental validation study. Front. Nutr. 2023, 10, 1121498. [Google Scholar] [CrossRef] [PubMed]
- Eklund, L.; Piuhola, J.; Komulainen, J.; Sormunen, R.; Ongvarrasopone, C.; Fassler, R.; Muona, A.; Ilves, M.; Ruskoaho, H.; Takala, T.E.; et al. Lack of type XV collagen causes a skeletal myopathy and cardiovascular defects in mice. Proc. Natl. Acad. Sci. USA 2001, 98, 1194–1199. [Google Scholar] [CrossRef] [PubMed]
- Rasi, K.; Piuhola, J.; Czabanka, M.; Sormunen, R.; Ilves, M.; Leskinen, H.; Rysa, J.; Kerkela, R.; Janmey, P.; Heljasvaara, R.; et al. Collagen XV is necessary for modeling of the extracellular matrix and its deficiency predisposes to cardiomyopathy. Circ. Res. 2010, 107, 1241–1252. [Google Scholar] [CrossRef]
- Zaferani, A.; Talsma, D.T.; Yazdani, S.; Celie, J.W.; Aikio, M.; Heljasvaara, R.; Navis, G.J.; Pihlajaniemi, T.; van den Born, J. Basement membrane zone collagens XV and XVIII/proteoglycans mediate leukocyte influx in renal ischemia/reperfusion. PLoS ONE 2014, 9, e106732. [Google Scholar] [CrossRef]
- Satish, L.; LaFramboise, W.A.; O’Gorman, D.B.; Johnson, S.; Janto, B.; Gan, B.S.; Baratz, M.E.; Hu, F.Z.; Post, J.C.; Ehrlich, G.D.; et al. Identification of differentially expressed genes in fibroblasts derived from patients with Dupuytren’s Contracture. BMC Med. Genom. 2008, 1, 10. [Google Scholar] [CrossRef]
- Wu, J.; Ren, W.; Zheng, Z.; Huang, Z.; Liang, T.; Li, F.; Shi, Z.; Jiang, Q.; Yang, X.; Guo, L. Mmu_circ_003795 regulates osteoblast differentiation and mineralization in MC3T3-E1 and MDPC23 by targeting COL15A1. Mol. Med. Rep. 2020, 22, 1737–1746. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | Flow Rate (mL/min) | Mobile Phase A (%) | Mobile Phase B (%) |
|---|---|---|---|
| 0 | 1 | 97 | 3 |
| 10 | 1 | 95 | 5 |
| 20 | 1 | 80 | 20 |
| 27 | 1 | 60 | 40 |
| 29 | 1 | 50 | 50 |
| 30 | 1 | 30 | 70 |
| 35 | 1 | 0 | 100 |
| Time (min) | Flow Rate (mL/min) | Mobile Phase A (%) | Mobile Phase B (%) |
|---|---|---|---|
| 0 | 1 | 97 | 3 |
| 10 | 1 | 95 | 5 |
| 12 | 1 | 90 | 10 |
| 13 | 1 | 85 | 15 |
| 50 | 1 | 60 | 40 |
| 53 | 1 | 40 | 60 |
| 54 | 1 | 0 | 100 |
| 58 | 1 | 0 | 0 |
| 60 | 1 | 0 | 0 |
| 70 | 1 | 0 | 0 |
| Compared Samples | Number of DEPs | Regulated Type | Fold Change > 1.5 |
|---|---|---|---|
| B vs. A | 620 | Up-regulated | 161 |
| Down-regulated | 166 | ||
| C vs. A | 622 | Up-regulated | 178 |
| Down-regulated | 201 | ||
| D vs. A | 611 | Up-regulated | 176 |
| Down-regulated | 188 | ||
| E vs. A | 604 | Up-regulated | 177 |
| Down-regulated | 185 | ||
| F_ vs. A | 620 | Up-regulated | 176 |
| Down-regulated | 191 | ||
| G vs. A | 623 | Up-regulated | 157 |
| Down-regulated | 216 | ||
| B, C, D, E, F_, G vs. A | 805 | Up-regulated | 226 |
| Down-regulated | 219 |
| GO Class | Up-Regulated | Down-Regulated |
|---|---|---|
| Molecular functions | Serine-type endopeptidase activity | Small molecule binding |
| Endopeptidase activity | Guanyl nucleotide binding | |
| Serine-type peptidase activity | Purine nucleotide binding | |
| Peptidase activity | Nucleotide binding | |
| Acting on L-amino acid peptides | Organic cyclic compound binding | |
| Endopeptidase inhibitor activity | Heterocyclic compound binding | |
| Serine-type endopeptidase inhibitor activity | Purine ribonucleoside binding | |
| Lipid binding | Purine ribonucleotide binding | |
| Thiol oxidase activity | Purine ribonucleoside triphosphate binding | |
| GTP binding | ||
| Carbohydrate derivative binding | ||
| Biological process | Macromolecule metabolic process | |
| Blood coagulation, proteolysis | ||
| Hyaluronan metabolic process | ||
| Glycosaminoglycan metabolic process | ||
| Single multicellular organism process | ||
| Lipoprotein metabolic process | ||
| Platelet activation | ||
| Protein metabolic process | ||
| Metabolic process | ||
| Lipid transport | ||
| Organic substance metabolic process | ||
| Response to stress | ||
| Macromolecule localization | ||
| Cellular component | Extracellular region | |
| Extracellular region part | ||
| Extracellular space | ||
| Fibrinogen complex |
| Regulated Type | Metabolic Pathway |
|---|---|
| Up-regulated | Fatty acid biosynthesis |
| Fatty acid metabolism | |
| Sphingolipid metabolism | |
| Protein processing in the endoplasmic reticulum | |
| Adipocytokine signaling pathway | |
| Peroxisome | |
| Caffeine metabolism | |
| Nicotinate and nicotinamide metabolism | |
| Bile secretion | |
| Purine metabolism | |
| Galactose metabolism | |
| Fatty acid degradation | |
| Platinum drug resistance | |
| Amino sugar and nucleotide sugar metabolism | |
| Long-term depression | |
| N-glycan biosynthesis | |
| RNA transport | |
| Down-regulated | Complement and coagulation cascades |
| Staphylococcus aureus infection | |
| Systemic lupus erythematosus | |
| Glycosaminoglycan biosynthesis—keratan sulfate | |
| Neuroactive ligand–receptor interaction | |
| Glycosphingolipid biosynthesis—lacto and neolacto series |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, M.; Huang, F.; Du, X.; Liu, G.; Wang, C. Analysis of the Differentially Expressed Proteins in Donkey Milk in Different Lactation Stages. Foods 2023, 12, 4466. https://doi.org/10.3390/foods12244466
Zhou M, Huang F, Du X, Liu G, Wang C. Analysis of the Differentially Expressed Proteins in Donkey Milk in Different Lactation Stages. Foods. 2023; 12(24):4466. https://doi.org/10.3390/foods12244466
Chicago/Turabian StyleZhou, Miaomiao, Fei Huang, Xinyi Du, Guiqin Liu, and Changfa Wang. 2023. "Analysis of the Differentially Expressed Proteins in Donkey Milk in Different Lactation Stages" Foods 12, no. 24: 4466. https://doi.org/10.3390/foods12244466
APA StyleZhou, M., Huang, F., Du, X., Liu, G., & Wang, C. (2023). Analysis of the Differentially Expressed Proteins in Donkey Milk in Different Lactation Stages. Foods, 12(24), 4466. https://doi.org/10.3390/foods12244466