
Activation of Immune and Antiviral Effects by Euglena Extracts: A Review

Abstract
1. Introduction
2. Effects of Euglena Intake on Allergic Diseases
3. Effects of Euglena’s Intake on the Intestinal Microbiota and Defecation
4. Effect of Euglena Intake on Symptoms of Influenza Virus Infection
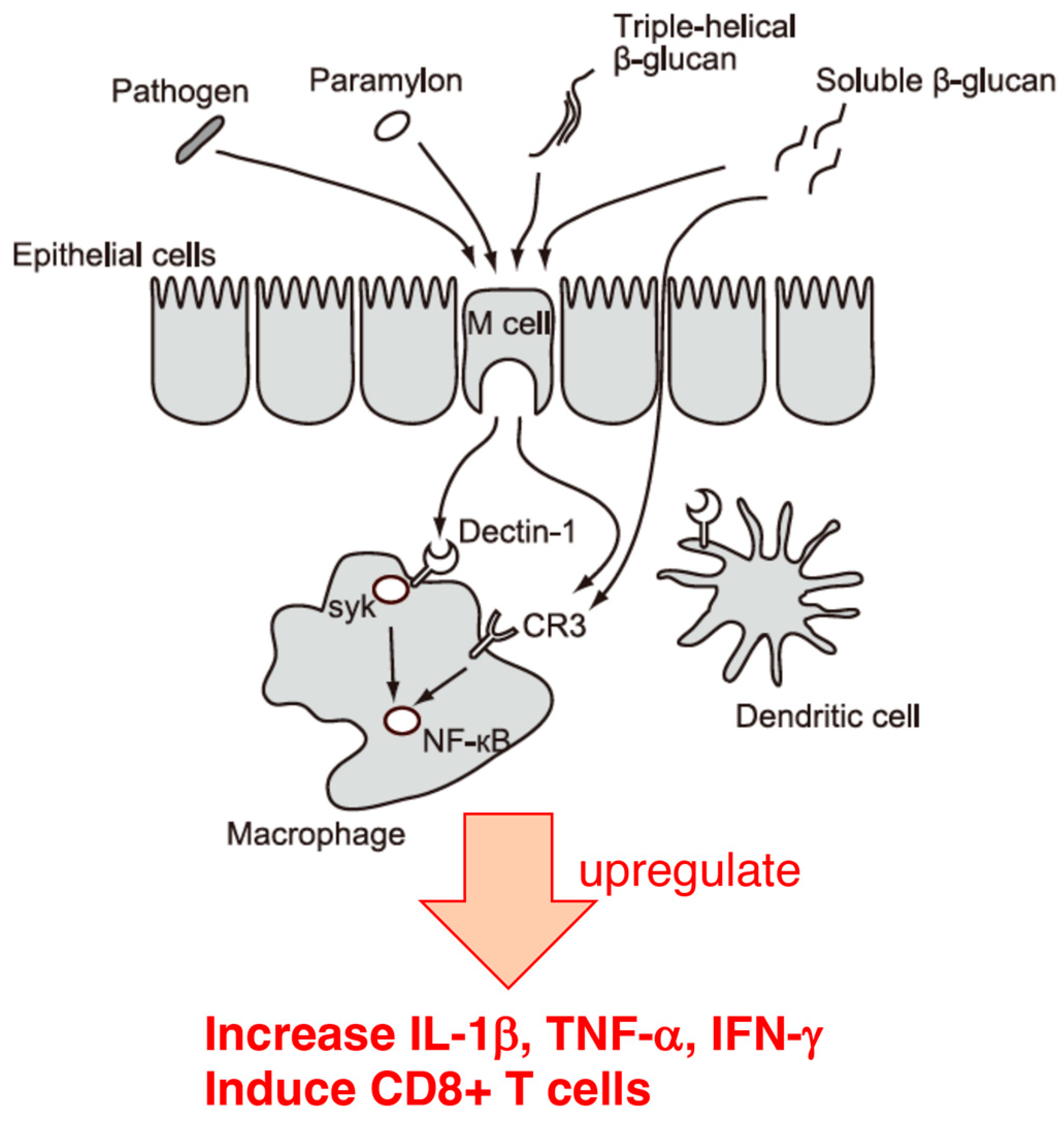
5. Effect of Euglena on Cellular Infection with the Influenza Virus
6. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ministry of Health, Labour and Welfare, Japan. 2020. The National Health and Nutrition Survey in Japan. 2018. Available online: https://www.mhlw.go.jp/content/00106684 (accessed on 1 August 2023).
- Daou, C.; Zhang, H. Oat beta-glucan: Its role in health promotion and prevention of diseases. Compr. Rev. Food Sci. Food Saf. 2012, 11, 355–365. [Google Scholar] [CrossRef]
- Chan, G.C.-F.; Chan, W.K.; Sze, D.M.-Y. The effects of beta-glucan on human immune and cancer cells. J. Hematol. Oncol. 2009, 2, 25. [Google Scholar] [CrossRef]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2015, 50, 24–31. [Google Scholar] [CrossRef]
- Tada, R.; Ikeda, F.; Aoki, K.; Yoshikawa, M.; Kato, Y.; Adachi, Y.; Tanioka, A.; Ishibashi, K.; Tsubaki, K.; Ohno, N. Barley-derived beta-D-glucan induces immunostimulation via a dectin-1-mediated pathway. Immunol. Lett. 2009, 123, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, D.; Iwai, A.; Aoki, S.; Uchiyama, H.; Kawata, K.; Nakayama, Y.; Nikawa, Y.; Kusano, K.; Okabe, M.; Miyazaki, T. β-Glucan derived from Aureobasidium pullulans is effective for the prevention of influenza in mice. PLoS ONE 2012, 7, e41399. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived beta-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef]
- Gupta, M.; Abu-Ghannam, N.; Gallaghar, E. Barley for brewing: Characteristic changes during malting, brewing and applications of its by-products. Compr. Rev. Food Sci. Food Saf. 2010, 9, 318–328. [Google Scholar] [CrossRef]
- Reyna-Villasmil, N.; Bermúdez-Pirela, V.; Mengual-Moreno, E.; Arias, N.; Cano-Ponce, C.; Leal-Gonzalez, E.; Souki, A.; Inglett, G.E.; Israili, Z.H.; Hernández-Hernández, R.; et al. Oat-derived beta-glucan significantly improves HDLC and diminishes LDLC and non-HDL cholesterol in overweight individuals with mild hypercholesterolemia. Am. J. Ther. 2007, 14, 203–212. [Google Scholar] [CrossRef]
- Battilana, P.; Ornstein, K.; Minehira, K.; Schwarz, J.M.; Acheson, K.; Schneiter, P.; Burri, J.; Jéquier, E.; Tappy, L. Mechanisms of action of beta-glucan in postprandial glucose metabolism in healthy men. Eur. J. Clin. Nutr. 2001, 55, 327–333. [Google Scholar] [CrossRef]
- Kofuji, K.; Aoki, A.; Tsubaki, K.; Konishi, M.; Isobe, T.; Murata, Y. Antioxidant activity of β-glucan. ISRN Pharm. 2012, 2012, 125864. [Google Scholar] [CrossRef]
- Bayrak, O.; Turgut, F.; Karatas, O.F.; Cimentepe, E.; Bayrak, R.; Catal, F.; Atis, O.; Akcay, A.; Unal, D. Oral beta-glucan protects kidney against ischemia/reperfusion injury in rats. Am. J. Nephrol. 2008, 28, 190–196. [Google Scholar] [CrossRef]
- Sener, G.; Toklu, H.; Ercan, F.; Erkanli, G. Protective effect of beta-glucan against oxidative organ injury in a rat model of sepsis. Int. Immunopharmacol. 2005, 5, 1387–1396. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, T.; Furue, H.; Kimura, T.; Kondo, T.; Hattori, T.; Ogawa, N. Clinical efficacy of lentinan on neoplastic diseases. Adv. Exp. Med. Biol. 1983, 166, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Fujimoto, S.; Orita, K.; Kimura, T.; Kondo, T.; Taguchi, T.; Yoshida, K.; Ogawa, N.; Furue, H. Clinical evaluation of SPG (schizophyllan) as a therapeutic adjuvant after surgery of gastric cancer--controlled study by an envelope method. Gan Kagaku Ryoho. 1983, 10, 1135–1145. [Google Scholar]
- Isoda, N.; Eguchi, Y.; Nukaya, H.; Hosho, K.; Suga, Y.; Suga, T.; Nakazawa, S.; Sugano, K. Clinical efficacy of superfine dispersed lentinan (beta-1,3-glucan) in patients with hepatocellular carcinoma. Hepatogastroenterology 2009, 56, 437–441. [Google Scholar] [PubMed]
- Shimizu, K.; Watanabe, S.; Watanabe, S.; Matsuda, K.; Suga, T.; Nakazawa, S.; Shiratori, K. Efficacy of oral administered superfine dispersed lentinan for advanced pancreatic cancer. Hepatogastroenterology 2009, 56, 240–244. [Google Scholar] [PubMed]
- Fedorov, S.N.; Ermakova, S.P.; Zvyagintseva, T.N.; Stonik, V.A. Anticancer and cancer preventive properties of marine polysaccharides: Some results and prospects. Mar. Drugs 2013, 11, 4876–4901. [Google Scholar] [CrossRef]
- Matsumoto, T.; Inui, H.; Miyatake, K.; Nakano, Y.; Murakami, K. Comparison of nutrients in Euglena with those in other respresentative food sources. Eco-Engineering 2009, 21, 81–86. [Google Scholar]
- Kott, Y.; Wachs, A.M. Amino acid composition of bulk Protein of Euglena growth in wast water. Appl. Microbiol. 1964, 12, 292–294. [Google Scholar] [CrossRef]
- Wang, Y.; Seppänen-Laakso, T.; Rischer, H.; Wiebe, M.G. Euglena gracilis growth and cell composition under different temperature, light and trophic conditions. PLoS ONE 2017, 13, e0195329. [Google Scholar] [CrossRef]
- Meyer, A.; Cirpus, P.; Ott, C.; Schlecker, R.; Zähringer, U.; Heinz, E. Biosynthesis of docosahexaenoic acid in Euglena gracilis: Biochemical and Molecular evidence for the involvement of a ∆4-fatty acyl group desaturase. Biochemistry 2003, 42, 9779–9788. [Google Scholar] [CrossRef]
- Monfils, A.K.; Triemer, R.E.; Bellairs, E.F. Characterization of paramylon morphological diversity in photosynthetic euglenoids (Euglenales, Euglenophyta). Phycologia 2011, 50, 156–169. [Google Scholar] [CrossRef]
- Huang, Y.; Wan, X.; Zhao, Z.; Liu, H.; Wu, W.; Ge, X.; Zhao, C. Metabolomic analysis and pathway profiling of paramylon production in Euglena gracilis grown on different carbon sources. Int. J. Biol. Macromol. 2023, 246, 125661. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, F.X.; Schiff, J.A. Chlorophyll-protein complexes from Euglena gracilis and mutants deficient in chlorophyll b: I. Pigment composition. Plant Physiol. 1986, 80, 223–230. [Google Scholar] [CrossRef]
- Yso, R.; Fu, W.; Du, M.; Chen, Z.-X.; Lei, A.-P.; Wang, J.-X. Carotinoids biosynthesis, accumulation, and applications of a model microalga Euglena gracilis. Mar. Drugs 2022, 20, 496. [Google Scholar] [CrossRef]
- Aleksijevic, A.; Grove, J.; Schuber, F. Studies on polyamine biosynthesis in Euglena gracilis. Biochim. Biophys. Acta 1979, 565, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Deslandes, Y.; Marchessault, R.H.; Sarko, A. Triple-helical structure of (1→3)-β-D-glucan. Macromolecules 1980, 13, 1466–1471. [Google Scholar] [CrossRef]
- Clarke, A.E.; Stone, B.A. Structure of the paramylon from Euglena gracilis. Biochim. Biophys. Acta 1960, 44, 161–163. [Google Scholar] [CrossRef]
- Kiss, J.Z.; Roberts, E.M.; Brown, R.M.; Triemer, R.E. X-ray and dissolution studies of paramylon storage granules from Euglena. Protoplasma 1988, 146, 150–156. [Google Scholar] [CrossRef]
- Chuah, C.T.; Sarko, A.; Deslandes, Y.; Marchessault, R.H. Packing analysis of carbohydrates and polysaccharides. Part 14. Triple-helical crystalline structure of curdlan and paramylon hydrates. Macromolecules 1983, 16, 1375–1382. [Google Scholar] [CrossRef]
- Barsanti, L.; Passarelli, V.; Evangelista, V.; Frassanito, A.M.; Gualtieri, P. Chemistry, physico-chemistry and applications linked to biological activities of β-glucans. Nat. Prod. Rep. 2011, 28, 457–466. [Google Scholar] [CrossRef]
- Shimada, R.; Fujita, M.; Yuasa, M.; Sawamura, H.; Watanabe, T.; Nakashima, A.; Suzuki, K. Oral administration of green algae, Euglena gracilis, inhibits hyperglycemia in OLETF rats, a model of spontaneous type 2 diabetes. Food Funct. 2016, 7, 4655–4659. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Yasuda, K.; Murata, A.; Suzuki, K.; Miura, N. Effects of Euglena gracilis Intake on Mood and Autonomic Activity under Mental Workload, and Subjective Sleep Quality: A Randomized, Double-Blind, Placebo-Controlled Trial. Nutrients 2020, 12, 3243. [Google Scholar] [CrossRef] [PubMed]
- Okubo, K.; Kurono, Y.; Ichimura, K.; Enomoto, T.; Okamoto, Y.; Kawauchi, H.; Suzuki, H.; Fujieda, S.; Masuyama, K. The Japanese Society of Allergology. 2017. Japanese guidelines for allergic rhinitis. Allergol. Int. 2017, 66, 205–219. [Google Scholar] [CrossRef] [PubMed]
- Yasueda, H.; Yui, Y.; Shimizu, T. Isolation and partial characterization of the major allergen from Japanese cedar (Cryptomeria japonica) polln. J. Allergy Clin. Immunol. 1983, 71, 77. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, M.; Inouye, S.; Taniai, M.; Ando, S.; Usui, M.; Matuhasi, T. Identification of the second major allergen of Japanese cedar pollen. Allergy 1990, 45, 309–312. [Google Scholar] [CrossRef]
- Fujiwara, D.; Wakabayashi, H.; Watanabe, H.; Nishida, S.; Iino, H. A double-blind trial of Lactobacillus paracasei strain KW3110 administration for immunomodulation in patients with pollen allergy. Allergol. Int. 2005, 54, 143–149. [Google Scholar] [CrossRef]
- Maeda-Yamamoto, M.; Ema, K.; Monobe, M.; Shibuichi, I.; Shinoda, Y.; Yamamoto, T.; Fujisawa, T. The efficancy of early treatment of seasonal allergic rhinitis with benifuuki green tea containing O-methylated catechin before pollen exposure: An open randomized study. Allergol. Int. 2009, 58, 437–444. [Google Scholar] [CrossRef]
- Sato, Y.; Akiyama, H.; Sugamura, H.; Watanabe, T.; Hamano-Nagaoka, M.; Inakuma, T.; Goda, Y.; Maitani, T. The feeding of β-carotene down-regulates serum IgE levels and inhibits the type I allergic response in mice. Biol. Pharm. Bull. 2004, 27, 978–984. [Google Scholar] [CrossRef]
- Kopf, M.; Le Gros, G.; Bachmann, M.; Lamers, M.C.; Bluethmann, H.; Köhler, G. Disruption of the murine IL-4gene blocks Th2 cytokine respnses. Nature 1993, 362, 245–248. [Google Scholar] [CrossRef]
- Kondo, Y.; Kato, A.; Hojo, H.; Nozoe, S.; Takeuchi, M.; Ochi, K. Cytokine-related immunopotentiating activities of paramylon, a β-(1→3)-D-glucan from Euglena gracilis. J. Pharmacobio.-Dyn. 1992, 15, 617–621. [Google Scholar] [CrossRef]
- Sugiyama, A.; Suzuki, K.; Mitra, S.; Arashida, R.; Yoshida, E.; Nakano, R.; Yabuta, Y.; Takeuchi, T. Hepatoprotective effects of pramylon, a β-1,3-D-glucan isolated from Euglena gracilis Z, on acute liver injury induced by carbon tetrachloride in rats. J. Vet. Med. Sci. 2009, 71, 885–890. [Google Scholar] [CrossRef]
- Koizumi, N.; Sakagami, H.; Utsumi, A.; Fujinaga, S.; Takada, M.; Asano, K.; Sugawara, I.; Ichikawa, S.; Kondo, H.; Mori, S.; et al. Anti-HIV (human immunodeficiency virus) activity of sulfated paramylon. Antivir. Res. 1993, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Sakagami, H.; Kikuchi, K.; Takeda, M.; Sato, T.; Ichikawa, S.; Fujimaki, M.; Wada, C.; Komatsu, N. Macrophage stimulation activity of antimicrobial N, N-dimethylaminoethyl paramylon. In Vivo 1991, 5, 101–105. [Google Scholar] [PubMed]
- Sugiyama, A.; Hata, S.; Suzuki, K.; Yoshida, E.; Nakano, R.; Mitra, S.; Arashida, R.; Asayama, Y.; Yabuta, Y.; Takeuchi, T. Oral administration of paramylon, a β-1,3-glucan isolated from Euglena gracilis Z inhibits development of atopic dermatitis-like skin lesions in NC/Nga mice. J. Vet. Med. Sci. 2010, 72, 755–763. [Google Scholar] [CrossRef]
- Matsuda, H.; Watanabe, N.; Geba, G.P.; Sperl, J.; Tsudzuki, M.; Hiroi, J.; Matsumoto, M.; Ushio, H.; Saito, S.; Askenase, P.W.; et al. Development of atopic dermatitis-like skin lesion with IgE hyperproduction in NC/Nga mice. Int. Immunol. 1997, 9, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Suto, H.; Matsuda, H.; Mitsuishi, K.; Hira, K.; Uchida, T.; Unno, T.; Ogawa, H.; Ra, C. NC/Nga mice: A mouse model for atopic dermatitis. Int. Arch. Allergy Immunol. 1999, 120, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Iida, M.; Desamero, M.J.; Yasuda, K.; Nakashima, A.; Suzuki, K.; Chambers, J.K.; Uchida, K.; Ogawa, R.; Hachimura, S.; Nakayama, J.; et al. Effect of orally administered Euglena gracilis and its reserve polysaccharide, paramylon, on gastric dysplasia in A4gnt knockout mice. Sci. Rep. 2021, 11, 13640. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Shimada, R.; Matsuyama, A.; Yuasa, M.; Sawamura, H.; Yoshida, E.; Suzuki, K. Antitumor activity of the β-glucan paramylon from Euglena against preneoplastic colonic aberrant crypt foci in mice. Food Funct. 2013, 4, 1685–1690. [Google Scholar] [CrossRef]
- Kimura, Y.; Sumiyoshi, M.; Suzuki, T.; Sakanaka, M. Inhibitory effects of water-soluble low-molecular-weight beta-(1,3-1,6) d-glucan purified from Aureobasidium pullulans GM-NH-1A1 strain on food allergic reactions in mice. Int. Immunopharmacol. 2007, 7, 963–972. [Google Scholar] [CrossRef]
- Yamada, J.; Hamuro, J.; Hatanaka, H.; Hamabata, K.; Kinoshita, S. Alleviation of seasonal allergic symptoms with super fine beta-1,3-glucan: A randomized study. J. Allergy Clin. Immunol. 2007, 119, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Koizumi, M.; Yoshii, Y.; Yuasa, M.; Sawamura, H.; Watanabe, T.; Yoshida, E.; Suzuki, K. Effect of euglena on cedar pollen allergies in cry j1-sensitized mice. J. Integr. Study Diet. Habits 2013, 24, 171–176. [Google Scholar] [CrossRef][Green Version]
- Lazar, V.; Ditu, L.-M.; Pircalabioru, G.G.; Gheorghe, I.; Curutiu, C.; Holban, A.M.; Picu, A.; Petcu, L.; Chifiriuc, M.C. Aspects of gut microbiota and immune system interactions in infectious disease, immunopathology, and cancer. Front. Immunol. 2018, 9, 1830. [Google Scholar] [CrossRef]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef]
- Cho, I.; Blaser, M.J. The human microbiome: At the interface of health and disease. Nat. Rev. Genet. 2012, 13, 260–270. [Google Scholar] [CrossRef]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Laparra, J.M.; Sanz, Y. Interactions of gut microbiota with functional food components and nutraceuticals. Pharmacol. Res. 2010, 61, 219–225. [Google Scholar] [CrossRef]
- Meijer, K.; de Vos, P.; Priebe, M.G. Butyrate and other short-chain fatty acids as modulators of immunity: What relevance for health? Curr. Opin. Clin. Nutr. Metab. Care 2010, 13, 715–721. [Google Scholar] [CrossRef]
- Hague, A.; Elder, D.J.; Hicks, D.J.; Paraskeva, C. Apoptosis in colorectal tumour cells: Induction by the short chain fatty acids butyrate, propionate and acetate and by the bile salt deoxycholate. Int. J. Cancer 1995, 60, 400–406. [Google Scholar] [CrossRef]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef]
- Schneider, S.M.; Le Gall, P.; Girard-Pipau, F.; Piche, T.; Pompei, A.; Nano, J.L.; Hébuterne, X.; Rampal, P. Total artificial nutrition is associated with major changes in the fecal flora. Eur. J. Nutr. 2000, 39, 248–255. [Google Scholar] [CrossRef] [PubMed]
- Graf, D.; Di Cagno, R.; Fåk, F.; Flint, H.J.; Nyman, M.; Saarela, M.; Watzl, B. Contribution of diet to the composition of the human gut microbiota. Microb. Ecol. Health Dis. 2015, 26, 26164. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, K.; Inoue, J.; Sasaki, D.; Hoshi, N.; Shirai, T.; Fukuda, I.; Azuma, T.; Kondo, A.; Osawa, R. Construction of a model culture system of human colonic microbiota to detect decreased Lachnospiraceae abundance and butyrogenesis in the feces of ulcerative colitis patients. Biotechnol. J. 2019, 14, e1800555. [Google Scholar] [CrossRef]
- Nakashima, A.; Sasaki, K.; Sasaki, D.; Yasuda, K.; Suzuko, K.; Kondo, A. The alga Euglena gracilis stimulates Faecalibacterium in the gut and contributes to increased defecation. Sci. Rep. 2021, 11, 1074. [Google Scholar] [CrossRef] [PubMed]
- Miquel, S.; Martin, R.; Rossi, O.; Bermúdez-Humarán, L.G.; Chatel, J.M.; Sokol, H.; Thomas, M.; Wells, J.M.; Langella, P. Faecalibacterium prausnitzii and human intestinal health. Curr. Opin. Microbiol. 2013, 16, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef]
- Jia, L.; Li, D.; Feng, N.; Shamoon, M.; Sun, Z.; Ding, L.; Zhang, H.; Chen, W.; Sun, J.; Chen, Y.Q. Anti-diabetic effects of Clostridium butyricum CGMCC0313.1 through promoting the growth of gut butyrate-producing bacteria in type 2 diabetic mice. Sci. Rep. 2017, 7, 7046. [Google Scholar] [CrossRef]
- Gissibl, A.; Sun, A.; Care, A.; Nevalainen, H.; Sunna, A. Bioproducts from Euglena gracilis: Synthesis and applications. Front. Bioeng. Biotechnol. 2019, 7, 108. [Google Scholar] [CrossRef]
- Sugimoto, R.; Ishibashi-Ohgo, N.; Atsuji, K.; Miwa, Y.; Iwata, O.; Nakashima, A.; Suzuki, K. Euglena extract suppresses adipocyte-differentiation in human adipose-derived stem cells. PLoS ONE 2018, 13, e0192404. [Google Scholar] [CrossRef]
- Okouchi, R.E.S.; Yamamoto, K.; Ota, T.; Seki, K.; Imai, M.; Ota, R.; Asayama, Y.; Nakashima, A.; Suzuki, K.; Tsuduki, T. Simultaneous intake of Euglena gracilis and vegetables exerts synergistic anti-obesity and anti-inflammatory effects by modulating the gut microbiota in diet-induced obese mice. Nutrients 2019, 11, 204. [Google Scholar] [CrossRef]
- Rios-Covian, D.; Gueimonde, M.; Duncan, S.H.; Flint, H.J.; de los Reyes-Gavilan, C.G. Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 2015, 362, fnv176. [Google Scholar] [CrossRef]
- Asayama, Y.; Suzuki, K.; Nakashima, A.; Shioya, N.; Sugimura, H. The study of the effects of food containing Euglena gracilis on improvement in constipation. Jpn. Pharmacol. Ther. 2017, 45, 1359–1364. [Google Scholar]
- Kawano, Y.; Nakano, Y.; Kitaoka, S.; Katou, K.; Shigeoka, S.; Ohnishi, T. Effects of Euglena cells on the absorption and tissue distribution of dietary cholesterol in rats. J. Jpn. Soc. Nutr. Food. Sci. 1987, 40, 193–198. [Google Scholar] [CrossRef]
- Yang, J.; Wang, H.-P.; Zhou, L.; Xu, C.-F. Effect of dietary fiber on constipation: A meta analysis. World J. Gastroenterol. 2012, 18, 7378–7383. [Google Scholar] [CrossRef] [PubMed]
- Pituch, A.; Walkowiak, J.; Banaszkiewicz, A. Butyric acid in functional constipation. Prz. Gastroenterol. 2013, 8, 295–298. [Google Scholar] [CrossRef] [PubMed]
- Trompette, A.; Gollwitzer, E.S.; Pattaroni, C.; Lopez-Mejia, I.C.; Riva, E.; Pernot, J.; Ubags, N.; Fajas, L.; Nicod, L.P.; Marsland, B.J. Dietary fiber confers protection against flu by shaping Ly6c– patrolling monocyte hematopoiesis and CD8+ T cell metabolism. Immunity 2018, 48, 992–1005.e8. [Google Scholar] [CrossRef] [PubMed]
- Taubenberger, J.K.; Morens, D.M. Influenza: The once and future pandemic. Public Health Rep. 2010, 125, 16–26. [Google Scholar] [CrossRef] [PubMed]
- McCaughey, C. Influenza: A virus of our times. Ulster Med. J. 2010, 79, 46–51. [Google Scholar]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Bronchoalveolar interferon-α, tumornecrosis factor-α, interleukin-1, and inflammation during acute influenza inpigs: A possible model for humans. J. Infect. Dis. 1998, 177, 1076–1079. [Google Scholar] [CrossRef][Green Version]
- Julkunen, I.; Sareneva, T.; Pirhonen, J.; Ronni, T.; Melén, K.; Matikaine, S. Molecular pathogenesis of influenzaA virus infection and virus-induced regulation of cytokine gene expression. Cytokine Growth Factor Rev. 2001, 12, 171–180. [Google Scholar] [CrossRef]
- Kaiser, L.; Fritz, R.S.; Straus, S.E.; Gubareva, L.; Hayden, F.G. Symptom pathogenesis during acuteinfluenza: Interleukin-6 and other cytokine responses. J. Med. Virol. 2001, 64, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Mok, K.P.; Wong, C.H.K.; Cheung, C.Y.; Chan, M.C.; Lee, S.M.Y.; Nicholls, J.M.; Peiris, J.S.M. Viral genetic determinants of H5N1 influenza viruses that contribute to cytokine dysregulation. J. Infect. Dis. 2009, 200, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Arimori, Y.; Nakamura, R.; Yamada, H.; Shibata, K.; Maeda, N.; Kase, T.; Yoshikai, Y. Type I interferon limits influenza virus-induced acute lung injury by regulation of excessive inflammation in mice. Antivir. Res. 2013, 99, 230–237. [Google Scholar] [CrossRef]
- Arimori, Y.; Nakamura, R.; Yamada, H.; Shibata, K.; Maeda, N.; Kase, T.; Yoshikai, Y. Type I interferon plays opposing roles in cytotoxicity and interferon-γ production by natural killer and CD8 T cells after influenza A virus infection in mice. J. Innate Immun. 2014, 6, 456–466. [Google Scholar] [CrossRef]
- Vetvicka, V.; Vetvickova, J. Glucan supplementation enhances the immune response against an influenza challenge in mice. Ann. Transl. Med. 2015, 3, 22. [Google Scholar] [CrossRef]
- Hassanzadeh-Kiabi, N.; Yáñez, A.; Dang, I.; Martins, G.A.; Underhill, D.M.; Goodridge, H.S. Autocrine type I IFN signaling in dendritic cells stimulated with fungal β-glucans or lipopolysaccharides promotes CD8 T cell activation. J. Immunol. 2017, 198, 375–382. [Google Scholar] [CrossRef]
- Gibbert, K.; Schlaak, J.F.; Yang, D.; Ditmer, U. IFN-α subtypes: Distinct biological activities in anti-viral therapy. Br. J. Pharmacol. 2013, 168, 1048–1058. [Google Scholar] [CrossRef]
- Hunter, C.A.; Chizzonite, R.; Remington, J.S. IL-1 beta is required for IL-12 to induce production of IFN-gamma by NK cells. A role for IL-1 beta in the T cell-independent mechanism of resistance against intracellular pathogens. J. Immunol. 1995, 155, 4347–4354. [Google Scholar] [CrossRef]
- Sabel, M.S.; Arora, A.; Su, G.; Mathiowitz, E.; Reineke, J.J.; Chang, A.E. Synergistic effect of intratumoral IL-12 and TNF-alpha microspheres: Systemic anti-tumor immunity is mediated by both CD8+ CTL and NK cells. Surgery 2007, 142, 749–760. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Gordon, S. Immune recognition. A new receptor for beta-glucans. Nature 2001, 413, 36–37. [Google Scholar] [CrossRef] [PubMed]
- Ujita, M.; Nagayama, H.; Kanie, S.; Koike, S.; Ikeyama, Y.; Ozaki, T.; Okumura, H. Carbohydrate binding specificity of recombinant human macrophage β-glucan receptor dectin-1. Biosci. Biotechnol. Biochem. 2009, 73, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Russo, R.; Barsanti, L.; Evangelista, V.; Frassanito, A.M.; Longo, V.; Pucci, L.; Penno, G.; Gualtieri, P. Euglena gracilis paramylon activates human lymphocytes by upregulating pro-inflammatory factors. Food Sci. Nutr. 2016, 5, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Legentil, L.; Paris, F.; Ballet, C.; Trouvelot, S.; Daire, X.; Vetvicka, V.; Ferrières, V. Molecular interactions of β-(1→3)-glucans with their receptors. Molecules 2015, 20, 9745–9766. [Google Scholar] [CrossRef] [PubMed]
- Bohn, J.A.; BeMiller, J.N. (1→3)-β-D-Glucans as biological response modifiers: A review of structure-functional activity relationships. Carbohydr. Polym. 1995, 28, 3–14. [Google Scholar] [CrossRef]
- Nakashima, A.; Suzuki, K.; Asayama, Y.; Konno, M.; Saito, K.; Yamazaki, N.; Takimoto, H. Oral administration of Euglena gracilis Z and its carbohydrate storage substance provides survival protection against influenza virus infection in mice. Biochem. Biophys. Res. Commun. 2017, 494, 379–383. [Google Scholar] [CrossRef]
- Nakashima, A.; Yamada, K.; Iwata, O.; Sugimoto, R.; Atsuji, K.; Ogawa, T.; Ishibashi-Ohgo, N.; Suzuki, K. β-Glucan in foods and its physiological functions. J. Nutr. Sci. Vitaminol. 2018, 64, 8–17. [Google Scholar] [CrossRef]
- Belshe, R.B.; Burk, K.; Newman, F.; Cerruti, R.L.; Sim, I.S. Resistance of influenza A virus to amantadine and rimantadine: Results of one decade of surveillance. J. Infect. Dis. 1989, 159, 430–435. [Google Scholar] [CrossRef]
- Bloom, J.D.; Gong, L.I.; Baltimore, D. Permissive secondary mutations enable the evolution of influenza oseltamivir resistance. Science 2010, 328, 1272–1275. [Google Scholar] [CrossRef]
- Le, Q.M.; Kiso, M.; Someya, K.; Sakai, Y.T.; Nguyen, T.H.; Ngyuen, K.H.; Pham, N.D.; Ngyen, H.H.; Yamada, S.; Muramoto, Y.; et al. Avian flu: Isolation of drug resistant H5N1 influenza A virus. Nature 2005, 437, 1108. [Google Scholar] [CrossRef]
- Seibert, C.W.; Rahmat, S.; Krammer, F.; Palese, P.; Bouvier, N.M. Efficient transmission of pandemic H1N1 influenza viruses with high-level oseltamivir resistance. J. Virol. 2012, 86, 5386–5389. [Google Scholar] [CrossRef]
- Nakashima, A.; Horio, Y.; Suzuki, K.; Isegawa, Y. Antiviral activity and underlying action mechanism of Euglena extract against influenza virus. Nutrients 2021, 13, 3911. [Google Scholar] [CrossRef]
- Barr, I.G.; Hurt, A.C.; Iannello, P.; Tomasov, C.; Deed, N.; Komadina, N. Increased adamantane resistance in influenza A(H3) viruses in Australia and neighbouring countries in 2005. Antivir. Res. 2007, 73, 112–117. [Google Scholar] [CrossRef] [PubMed]
- Nitsch-Osuch, A.; Brydak, L.B. Influenza viruses resistant to neuraminidase inhibitors. Acta Biochim. Pol. 2014, 61, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Todd, B.; Tchesnokov, E.P.; Götte, M. The active form of the influenza cap-snatching endonuclease inhibitor baloxavir marboxil is a tight binding inhibitor. J. Biol. Chem. 2021, 296, 100486. [Google Scholar] [CrossRef] [PubMed]
- Ishiguro, S.; Upreti, D.; Robben, N.; Burghart, R.; Loyd, M.; Ogun, D.; Le, T.; Delzeit, J.; Nakashima, A.; Thakkar, R.; et al. Water extract from Euglena gracilis prevents lung carcinoma growth in mice by attenuation of the my-eloid-derived cell population. Biomed. Pharmacother. 2020, 127, 110166. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-F.; Bai, L.-P.; Huang, W.-B.; Li, X.-Z.; Zhao, S.-S.; Zhong, N.-S.; Jiang, Z.-H. Comparison of in vitro antiviral activity of tea polyphenols against influenza A and B viruses and structure-activity relationship analysis. Fitoterapia 2014, 93, 47–53. [Google Scholar] [CrossRef]
- Nagamine, T.; Nakajima, K.; Takada, H.; Sekine, Y.; Suzuki, K. Induction of type 1 interferon receptor by zinc in U937 cells. Cytokine 2009, 46, 346–350. [Google Scholar] [CrossRef]
- Te Velthuis, A.J.; van den Worm, S.H.; Sims, A.C.; Baric, R.S.; Snijder, E.J.; van Hemert, M.J. Zn2+ inhibits coronavirus and arterivirus RNA polymerase activity in vitro and zinc ionophores block the replication of these viruses in cell culture. PLoS Pathog. 2010, 6, e1001176. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vitamins | Minerals | Amino Acids | Unsaturated Fatty Acids | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Content | Ref. | Content | Ref. | Ref. | Ref. | ||||
| (µg/106 cells) | (µg/106 cells) | ||||||||
| Vitamin B1 | 1.4 ± 0.1 | [19] | Zn2+ | 0.795 ± 0.104 | [19] | Alanine | [20] | Docosahexaenoic acid | [21,22] |
| Vitamin B2 | 3.5 ± 0.2 | [19] | Mg2+ | 0.165 ± 0.076 | [19] | Arginine | [20] | Eicosapentaenoic acid | [21] |
| Vitamin B6 | 7.5 ± 0.6 | [19] | Fe2+ | 0.153 ± 0.092 | [19] | Aspertic acid | [20] | α-Linolenic acid | [19,21] |
| Vitamin B12 | 0.05 ± 0.01 | [19] | Mn2+ | 0.240 ± 0.056 | [19] | Cystine | [20] | Arachidonic acid | [19,21] |
| Folic acid | 1.7 ± 0.5 | [19] | Ca2+ | 0.046 ± 0.070 | [19] | Glycine | [20] | Palmitoleic acid | [21] |
| Pantotheric acid | 18.2 ± 0.4 | [19] | Cu2+ | 0.004 ± 0.0023 | [19] | Glutamic acid | [20] | Oleic acid | [19,21] |
| Biotin | 4.6 ± 0.3 | [19] | Ni2+ | 0.006 ± 0.0014 | [19] | Histidine | [20] | Linoleic acid | [19,21] |
| Vitamin C | 27.2 ± 0.4 | [19] | Isoleucine | [19,20] | Eicosadienoic acid | [21] | |||
| Tocopherol | 517.5 ± 10.2 | [19] | Leucine | [19,20] | Docosatetraenoic Acid | [21] | |||
| β-Carotene | 0.71 ± 0.15 | [19] | Lysine | [19,20] | Docosapentaenoic acid | [21,22] | |||
| Phenylalanine | [20] | Dihomo-γ-linolenic acid | [21] | ||||||
| Preoline | [20] | ||||||||
| Serine | [20] | ||||||||
| Threonine | [19,20] | ||||||||
| Tryptophan | [19,20] | ||||||||
| Tyrosine | [20] | ||||||||
| Varine | [20] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Isegawa, Y. Activation of Immune and Antiviral Effects by Euglena Extracts: A Review. Foods 2023, 12, 4438. https://doi.org/10.3390/foods12244438
Isegawa Y. Activation of Immune and Antiviral Effects by Euglena Extracts: A Review. Foods. 2023; 12(24):4438. https://doi.org/10.3390/foods12244438
Chicago/Turabian StyleIsegawa, Yuji. 2023. "Activation of Immune and Antiviral Effects by Euglena Extracts: A Review" Foods 12, no. 24: 4438. https://doi.org/10.3390/foods12244438
APA StyleIsegawa, Y. (2023). Activation of Immune and Antiviral Effects by Euglena Extracts: A Review. Foods, 12(24), 4438. https://doi.org/10.3390/foods12244438