
Discovery and Engineering of a Novel Bacterial L-Aspartate α-Decarboxylase for Efficient Bioconversion



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strains and Culture Conditions
2.2. Plasmid Construction and Gene Mutagenesis
2.3. Expression and Purification of Recombinant Enzymes
2.4. Structure Modeling and Molecular Docking
2.5. Mutagenesis and Screening of L-Aspartate α-Decarboxylase
2.6. Measurement of Specific Activity
2.7. Determination of Kinetic Parameters
2.8. Characterization of Optimum Temperature and Themostability
2.9. Characterization of the Optimal pH and pH Stability
2.10. Determination of Melting Temperature (Tm)
2.11. Molecular Dynamics (MD) Simulation and Structural Analysis
2.12. Fed-Batch Cultivation in 5 L Fermenter
2.13. Whole-Cell Bioconversion
3. Results and Discussion
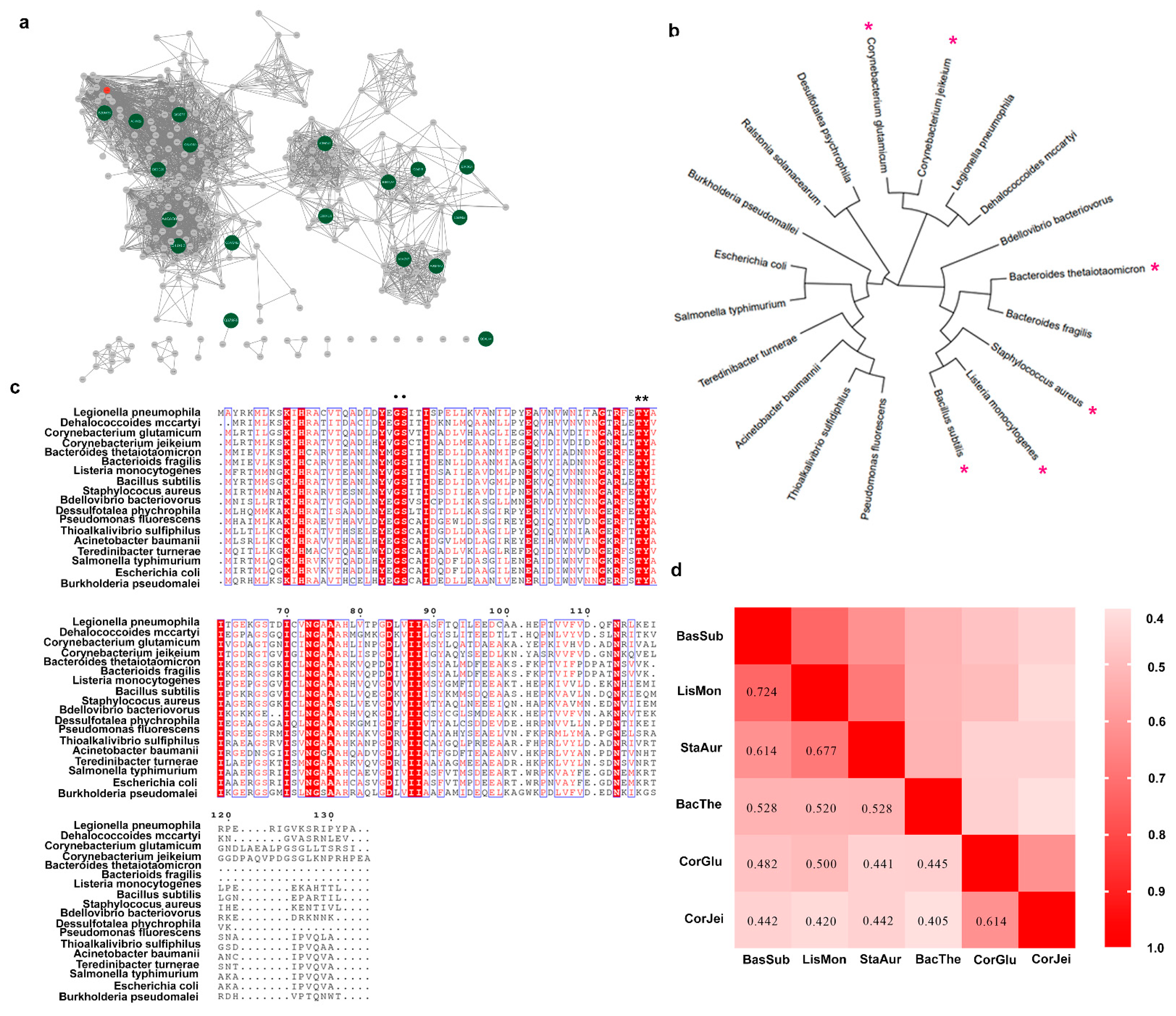
3.1. Mining a Novel L-Aspartate α-Decarboxylase
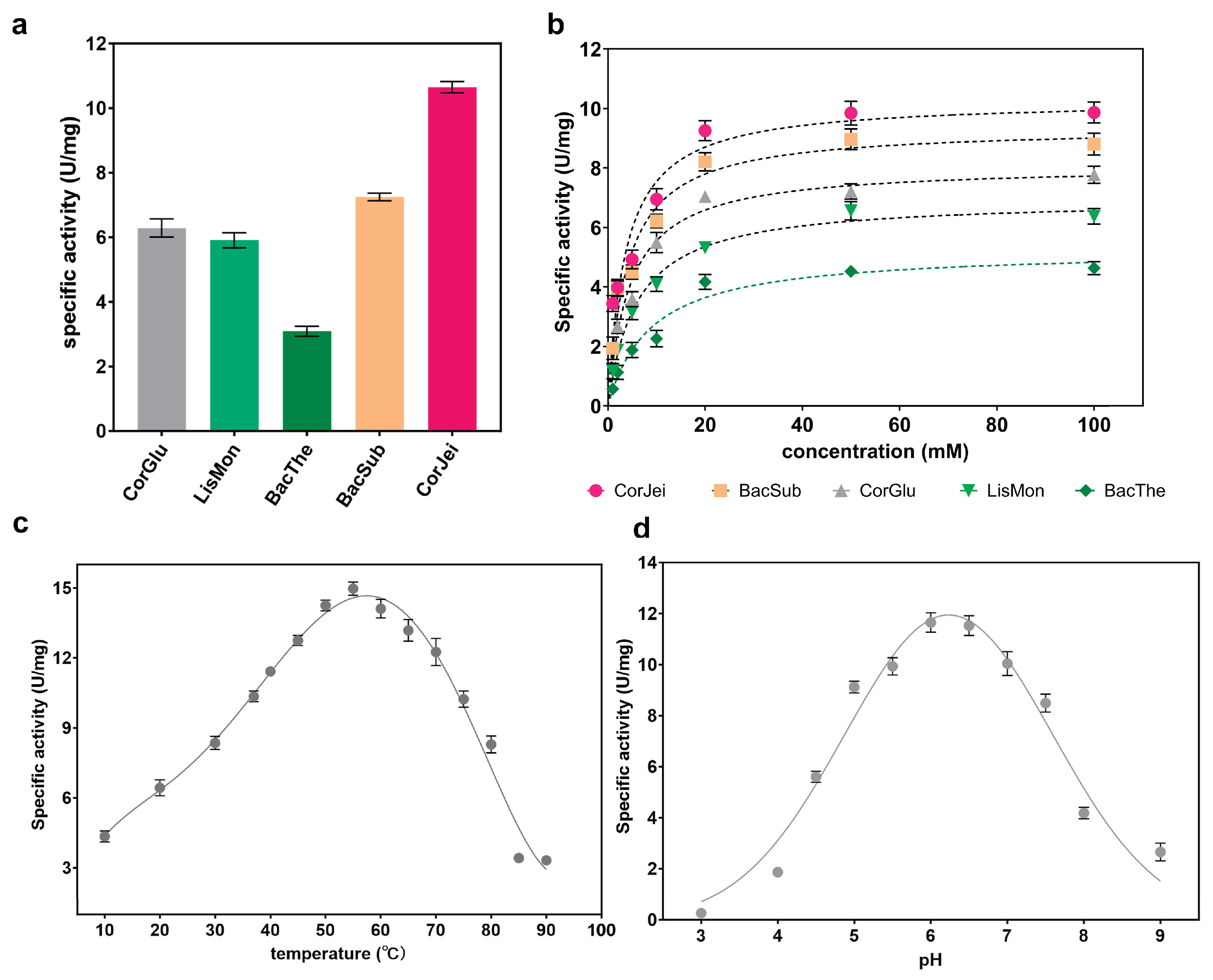
3.2. Comparison of the Whole-Cell Transformation Capacity with Different ADCs
3.3. Determination of the Enzymatic Properties
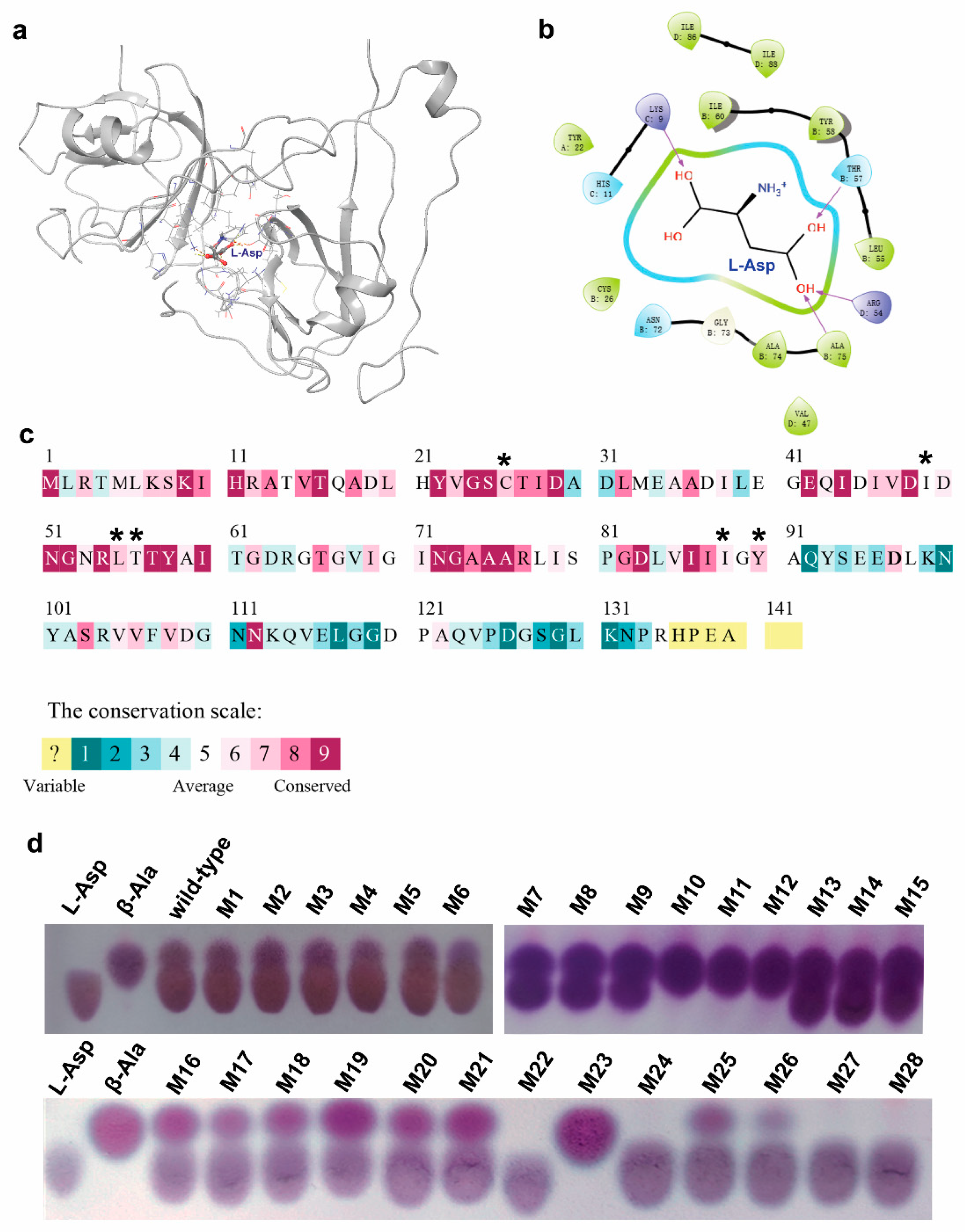
3.4. Generation of Single-Site Mutations and Screening of the Mutants
3.5. Generation of the Multiple-Site Mutations, Screening of the Mutants, and Characterization of the Catalytic Properties
3.6. Identification and Comparison of the Thermostability and pH Stability
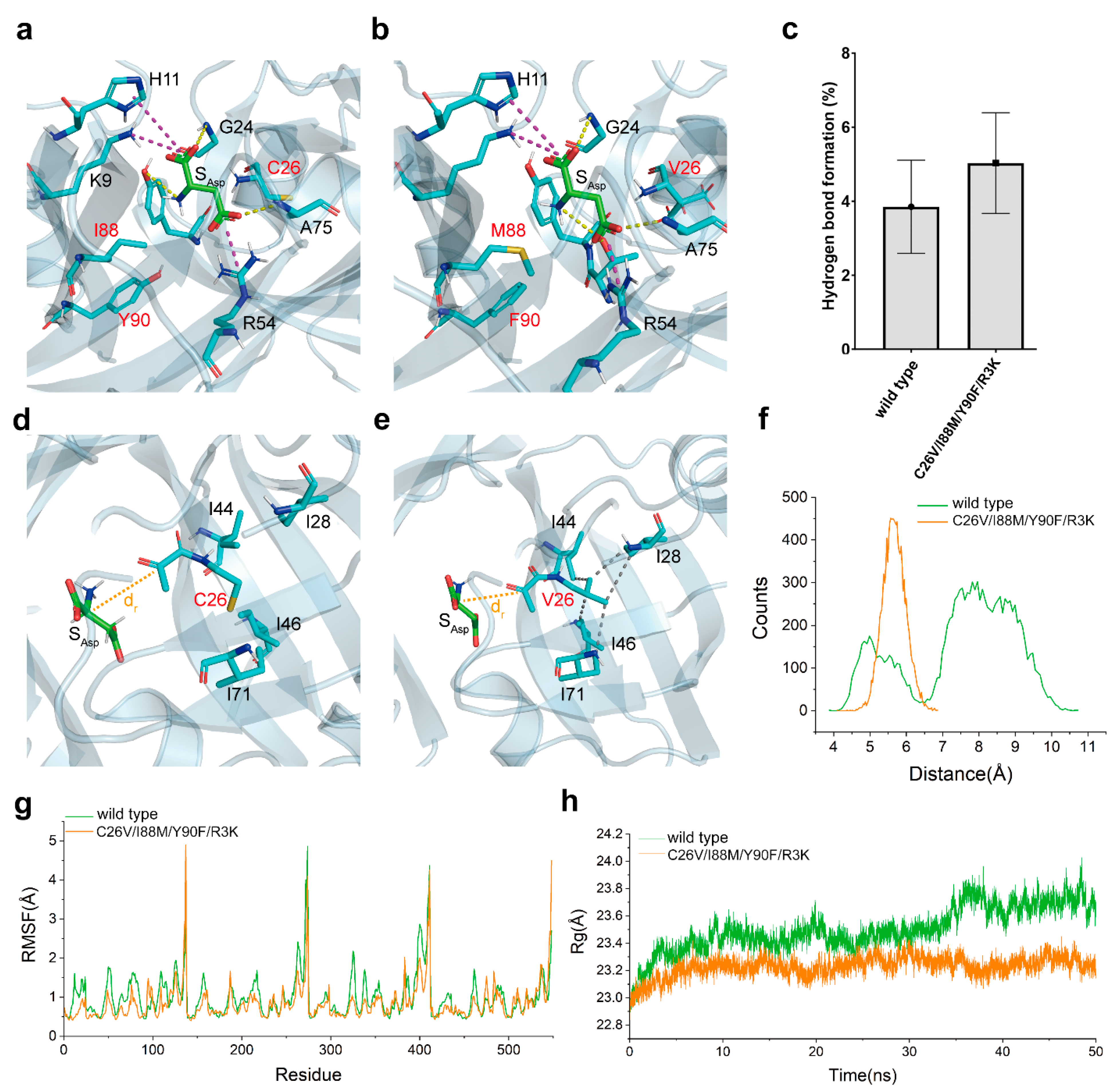
3.7. Insight into the Mechanism of Catalytic Activity Enhancement through Structural Analysis
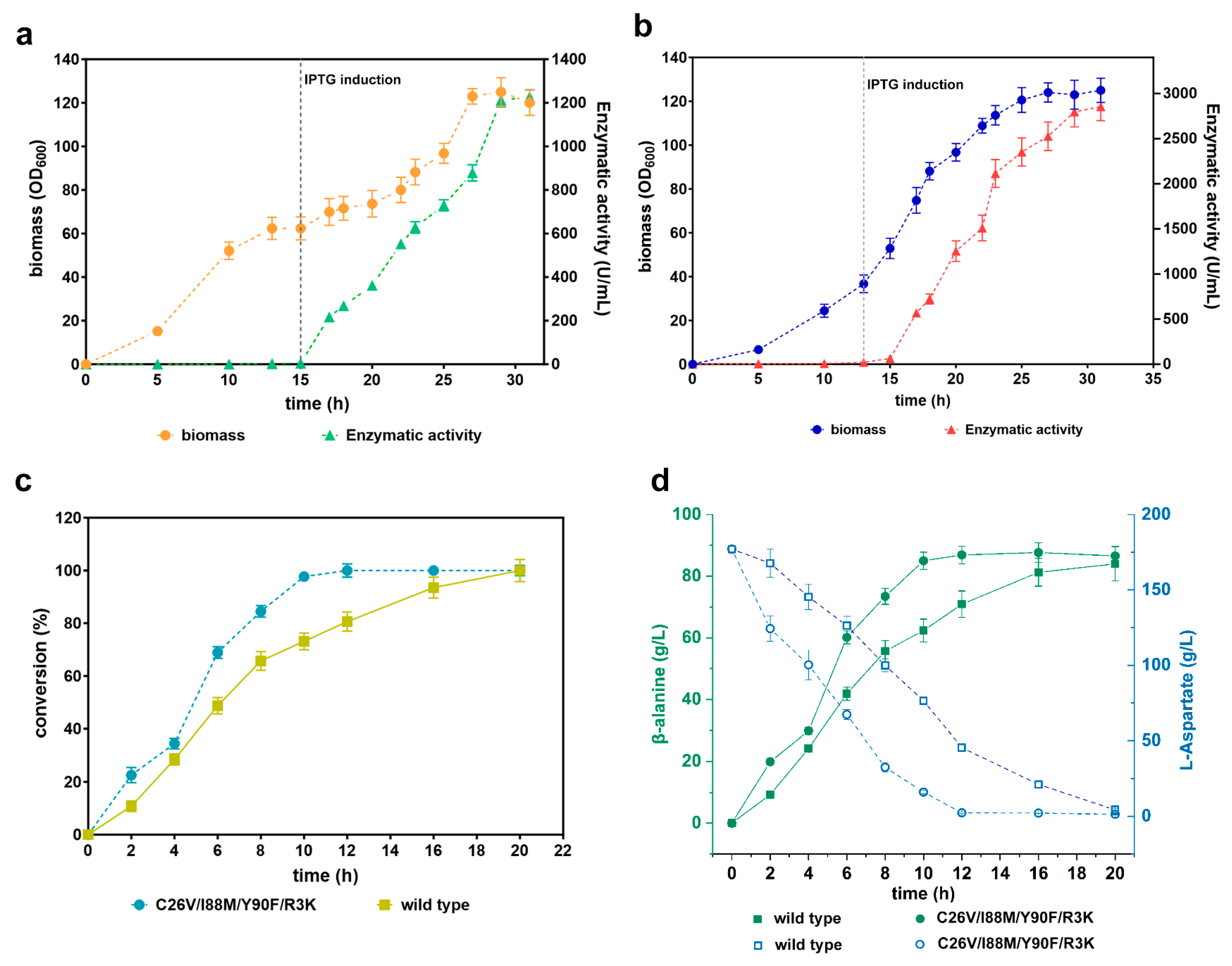
3.8. Scale-Up of the Recombinant CjADC-Producing Strains and Identification of the Bioconversion Efficiency
4. Conclusions and Perspective
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, B.I.; Suh, S.W. Crystal structure of the schiff base intermediate prior to decarboxylation in the catalytic cycle of aspartate alpha-decarboxylase. J. Mol. Biol. 2004, 340, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Gopalan, G.; Chopra, S.; Ranganathan, A.; Swaminathan, K. Crystal structure of uncleaved L-aspartate-alpha-decarboxylase from Mycobacterium tuberculosis. Proteins 2006, 65, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Könst, P.M.; Franssen, M.C.R.; Scott, E.L.; Sanders, J.P.M. A study on the applicability of L-aspartate α-decarboxylase in the biobased production of nitrogen containing chemicals. Green Chem. 2009, 11, 1646–1652. [Google Scholar] [CrossRef]
- Guo, J.; Sun, X.; Yuan, Y.; Chen, Q.; Ou, Z.; Deng, Z.; Ma, T.; Liu, T. Metabolic Engineering of Saccharomyces cerevisiae for Vitamin B5 Production. J. Agric. Food Chem. 2023, 71, 7408–7417. [Google Scholar] [CrossRef]
- Albert, A.; Dhanaraj, V.; Genschel, U.; Khan, G.; Ramjee, M.K.; Pulido, R.; Sibanda, B.L.; von Delft, F.; Witty, M.; Blundell, T.L.; et al. Crystal structure of aspartate decarboxylase at 2.2 A resolution provides evidence for an ester in protein self-processing. Nat. Struct. Biol. 1998, 5, 289–293. [Google Scholar] [CrossRef]
- Dusch, N.; Puhler, A.; Kalinowski, J. Expression of the Corynebacterium glutamicum panD gene encoding L-aspartate-alpha-decarboxylase leads to pantothenate overproduction in Escherichia coli. Appl. Environ. Microbiol. 1999, 65, 1530–1539. [Google Scholar] [CrossRef]
- Pei, W.; Zhang, J.; Deng, S.; Tigu, F.; Li, Y.; Li, Q.; Cai, Z.; Li, Y. Molecular engineering of L-aspartate-alpha-decarboxylase for improved activity and catalytic stability. Appl. Microbiol. Biotechnol. 2017, 101, 6015–6021. [Google Scholar] [CrossRef]
- Yu, X.J.; Huang, C.Y.; Xu, X.D.; Chen, H.; Liang, M.J.; Xu, Z.X.; Xu, H.X.; Wang, Z. Protein Engineering of a Pyridoxal-5’-Phosphate-Dependent l-Aspartate-alpha-Decarboxylase from Tribolium castaneum for beta-Alanine Production. Molecules 2020, 25, 1280. [Google Scholar] [CrossRef]
- Arakane, Y.; Lomakin, J.; Beeman, R.W.; Muthukrishnan, S.; Gehrke, S.H.; Kanost, M.R.; Kramer, K.J. Molecular and functional analyses of amino acid decarboxylases involved in cuticle tanning in Tribolium castaneum. J. Biol. Chem. 2009, 284, 16584–16594. [Google Scholar] [CrossRef]
- Liu, P.; Ding, H.; Christensen, B.M.; Li, J. Cysteine sulfinic acid decarboxylase activity of Aedes aegypti aspartate 1-decarboxylase: The structural basis of its substrate selectivity. Insect. Biochem. Mol. Biol. 2012, 42, 396–403. [Google Scholar] [CrossRef]
- Schmitzberger, F.; Kilkenny, M.L.; Lobley, C.M.; Webb, M.E.; Vinkovic, M.; Matak-Vinkovic, D.; Witty, M.; Chirgadze, D.Y.; Smith, A.G.; Abell, C.; et al. Structural constraints on protein self-processing in L-aspartate-alpha-decarboxylase. EMBO J. 2003, 22, 6193–6204. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, D.C.F.; Patel, V.; Bartlett, C.P.; Nozaki, S.; Grant, T.D.; Gowdy, J.A.; Thompson, G.S.; Kalverda, A.P.; Snell, E.H.; Niki, H.; et al. The structure of the PanD/PanZ protein complex reveals negative feedback regulation of pantothenate biosynthesis by coenzyme A. Chem. Biol. 2015, 22, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Yuan, S.F.; Nair, P.H.; Borbon, D.; Coleman, S.M.; Fan, P.H.; Lin, W.L.; Alper, H.S. Metabolic engineering of E. coli for beta-alanine production using a multi-biosensor enabled approach. Metab. Eng. 2022, 74, 24–35. [Google Scholar] [CrossRef]
- Tadi, S.R.R.; Nehru, G.; Sivaprakasam, S. Combinatorial approach for improved production of whole-cell 3-aminopropionic acid in recombinant Bacillus megaterium: Codon optimization, gene duplication and process optimization. 3 Biotech 2021, 11, 333. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Piao, X.; Cui, S.; Hu, M.; Tao, Y. Enhanced production of beta-alanine through co-expressing two different subtypes of L-aspartate-alpha-decarboxylase. J. Ind. Microbiol. Biotechnol. 2020, 47, 465–474. [Google Scholar] [CrossRef]
- Zhang, T.; Zhang, R.; Xu, M.; Zhang, X.; Yang, T.; Liu, F.; Yang, S.; Rao, Z. Glu56Ser mutation improves the enzymatic activity and catalytic stability of Bacillus subtilis. Process Chem. 2018, 70, 117–123. [Google Scholar] [CrossRef]
- Hu, S.; Fei, M.; Fu, B.; Yu, M.; Yuan, P.; Tang, B.; Yang, H.; Sun, D. Development of probiotic E. coli Nissle 1917 for beta-alanine production by using protein and metabolic engineering. Appl. Microbiol. Biotechnol. 2023, 107, 2277–2288. [Google Scholar] [CrossRef]
- Wang, J.Y.; Rao, Z.M.; Xu, J.Z.; Zhang, W.G. Enhancing beta-alanine production from glucose in genetically modified Corynebacterium glutamicum by metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2021, 105, 9153–9166. [Google Scholar] [CrossRef]
- Li, Y.F.; Hess, S.; Pannell, L.K.; White Tabor, C.; Tabor, H. In vivo mechanism-based inactivation of S-adenosylmethionine decarboxylases from Escherichia coli, Salmonella typhimurium, and Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2001, 98, 10578–10583. [Google Scholar] [CrossRef]
- Mo, Q.; Li, Y.; Wang, J.; Shi, G. Identification of mutations restricting autocatalytic activation of bacterial L-aspartate alpha-decarboxylase. Amino Acids 2018, 50, 1433–1440. [Google Scholar] [CrossRef]
- Qian, Y.; Lu, C.; Liu, J.; Song, W.; Chen, X.; Luo, Q.; Liu, L.; Wu, J. Engineering protonation conformation of l-aspartate-alpha-decarboxylase to relieve mechanism-based inactivation. Biotechnol. Bioeng. 2020, 117, 1607–1614. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Maguire, J.B.; Haddox, H.K.; Strickland, D.; Halabiya, S.F.; Coventry, B.; Griffin, J.R.; Pulavarti, S.; Cummins, M.; Thieker, D.F.; Klavins, E.; et al. Perturbing the energy landscape for improved packing during computational protein design. Proteins 2021, 89, 436–449. [Google Scholar] [CrossRef] [PubMed]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.; Mittal, J.; Feig, M.; Mackerell, A.D., Jr. Optimization of the additive CHARMM all-atom protein force field targeting improved sampling of the backbone phi, psi and side-chain chi(1) and chi(2) dihedral angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef]
- Phillips, J.C.; Hardy, D.J.; Maia, J.D.C.; Stone, J.E.; Ribeiro, J.V.; Bernardi, R.C.; Buch, R.; Fiorin, G.; Henin, J.; Jiang, W.; et al. Scalable molecular dynamics on CPU and GPU architectures with NAMD. J. Chem. Phys. 2020, 153, 044130. [Google Scholar] [CrossRef] [PubMed]
- Brooks, B.R.; Brooks, C.L., 3rd; Mackerell, A.D., Jr.; Nilsson, L.; Petrella, R.J.; Roux, B.; Won, Y.; Archontis, G.; Bartels, C.; Boresch, S.; et al. CHARMM: The biomolecular simulation program. J. Comput. Chem. 2009, 30, 1545–1614. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef] [PubMed]
- Vanommeslaeghe, K.; Raman, E.P.; MacKerell, A.D., Jr. Automation of the CHARMM General Force Field (CGenFF) II: Assignment of bonded parameters and partial atomic charges. J. Chem. Inf. Model. 2012, 52, 3155–3168. [Google Scholar] [CrossRef]
- Grant, B.J.; Skjaerven, L.; Yao, X.Q. The Bio3D packages for structural bioinformatics. Protein Sci. 2021, 30, 20–30. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, Z.; Cui, W.; Zhou, Z. Establishment of bioprocess for synthesis of nicotinamide by recombinant Escherichia coli expressing high-molecular-mass nitrile hydratase. Appl. Biochem. Biotechnol. 2017, 182, 1458–1466. [Google Scholar] [CrossRef]
- Oberg, N.; Zallot, R.; Gerlt, J.A. EFI-EST, EFI-GNT, and EFI-CGFP: Enzyme Function Initiative (EFI) Web Resource for Genomic Enzymology Tools. J. Mol. Biol. 2023, 435, 168018. [Google Scholar] [CrossRef]
- Zallot, R.; Oberg, N.; Gerlt, J.A. The EFI Web Resource for Genomic Enzymology Tools: Leveraging Protein, Genome, and Metagenome Databases to Discover Novel Enzymes and Metabolic Pathways. Biochemistry 2019, 58, 4169–4182. [Google Scholar] [CrossRef]
- Cui, W.; Shi, Z.; Fang, Y.; Zhou, L.; Ding, N.; Zhou, Z. Significance of Arg3, Arg54, and Tyr58 of L-aspartate alpha-decarboxylase from Corynebacterium glutamicum in the process of self-cleavage. Biotechnol. Lett. 2014, 36, 121–126. [Google Scholar] [CrossRef]
- Ghiffary, M.R.; Prabowo, C.P.S.; Adidjaja, J.J.; Lee, S.Y.; Kim, H.U. Systems metabolic engineering of Corynebacterium glutamicum for the efficient production of beta-alanine. Metab. Eng. 2022, 74, 121–129. [Google Scholar] [CrossRef]
- Song, C.W.; Lee, J.; Ko, Y.S.; Lee, S.Y. Metabolic engineering of Escherichia coli for the production of 3-aminopropionic acid. Metab. Eng. 2015, 30, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Abidin, M.Z.; Saravanan, T.; Zhang, J.; Tepper, P.G.; Strauss, E.; Poelarends, G.J. Modular Enzymatic Cascade Synthesis of Vitamin B(5) and Its Derivatives. Chemistry 2018, 24, 17434–17438. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhou, H.Y.; Li, B.; Ding, W.Q.; Liu, Z.Q.; Zheng, Y.G. Multiplex modification of Escherichia coli for enhanced beta-alanine biosynthesis through metabolic engineering. Bioresour. Technol. 2021, 342, 126050. [Google Scholar] [CrossRef] [PubMed]
- Miton, C.M.; Buda, K.; Tokuriki, N. Epistasis and intramolecular networks in protein evolution. Curr. Opin. Struct. Biol. 2021, 69, 160–168. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CorJei | BacSub | CorGlu | LisMon | BacThe | |
|---|---|---|---|---|---|
| Specific activity (U/mg) | 10.7 ± 0.3 | 7.2 ± 0.2 | 6.3 ± 0.1 | 5.8 ± 0.1 | 2.6 ± 0.2 |
| Km (mM) | 3.6 ± 0.2 | 4.2 ± 0.3 | 4.5 ± 0.1 | 6.0 ± 0.2 | 8.9 ± 0.4 |
| kcat (s−1) | 106.8 ± 0.2 | 90.6 ± 0.3 | 80.8 ± 0.2 | 69.6 ± 1.1 | 52.5 ± 1.2 |
| kcat/Km (mM−1·s−1) | 29.7 ± 0.7 | 21.6 ± 0.6 | 17.8 ± 0.4 | 11.6 ± 0.3 | 5.9 ± 0.2 |
| No. | Mutants | Specific Activity (U/mg) |
|---|---|---|
| M10 | I88L | 9.1 ± 0.2 |
| M11 | Y90F | 12.9 ± 0.2 |
| M12 | C26V | 12.4 ± 0.1 |
| M23 | I88M | 13.6 ± 0.3 |
| I88M/Y90F | I88M/C26V | Y90F/C26V | I88L/Y90F | C26V/I88M/Y90F | I88M/Y90F/C26V/R3K | |
|---|---|---|---|---|---|---|
| Specific activity (U/mg) | 15.8 ± 0.2 | 11.3 ± 0.1 | 7.8 ± 0.4 | 6.2 ± 0.3 | 18.8 ± 0.5 | 28.8 ± 0.8 |
| Km (mM) | 3.2 ± 0.4 | 3.6 ± 0.2 | 4.1 ± 0.3 | 3.9 ± 0.1 | 2.6 ± 0.6 | 2.2 ± 0.5 |
| kcat (s−1) | 136.8 ± 0.3 | 110.5 ± 1.3 | 98.6 ± 1.2 | 88.6 ± 2.8 | 171.5 ± 0.8 | 209.2 ± 0.8 |
| kcat/Km (mM−1·s−1) | 42.7 ± 0.7 | 30.5 ± 0.4 | 24.0 ± 0.3 | 22.7 ± 0.5 | 61.2 ± 1.3 | 95.1 ± 1.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, W.; Liu, H.; Ye, Y.; Han, L.; Zhou, Z. Discovery and Engineering of a Novel Bacterial L-Aspartate α-Decarboxylase for Efficient Bioconversion. Foods 2023, 12, 4423. https://doi.org/10.3390/foods12244423
Cui W, Liu H, Ye Y, Han L, Zhou Z. Discovery and Engineering of a Novel Bacterial L-Aspartate α-Decarboxylase for Efficient Bioconversion. Foods. 2023; 12(24):4423. https://doi.org/10.3390/foods12244423
Chicago/Turabian StyleCui, Wenjing, Hao Liu, Yan Ye, Laichuang Han, and Zhemin Zhou. 2023. "Discovery and Engineering of a Novel Bacterial L-Aspartate α-Decarboxylase for Efficient Bioconversion" Foods 12, no. 24: 4423. https://doi.org/10.3390/foods12244423
APA StyleCui, W., Liu, H., Ye, Y., Han, L., & Zhou, Z. (2023). Discovery and Engineering of a Novel Bacterial L-Aspartate α-Decarboxylase for Efficient Bioconversion. Foods, 12(24), 4423. https://doi.org/10.3390/foods12244423