
Echinacea purpurea (L.) Moench Polysaccharide Alleviates DSS-Induced Colitis in Rats by Restoring Th17/Treg Balance and Regulating Intestinal Flora


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Chemicals and Regents
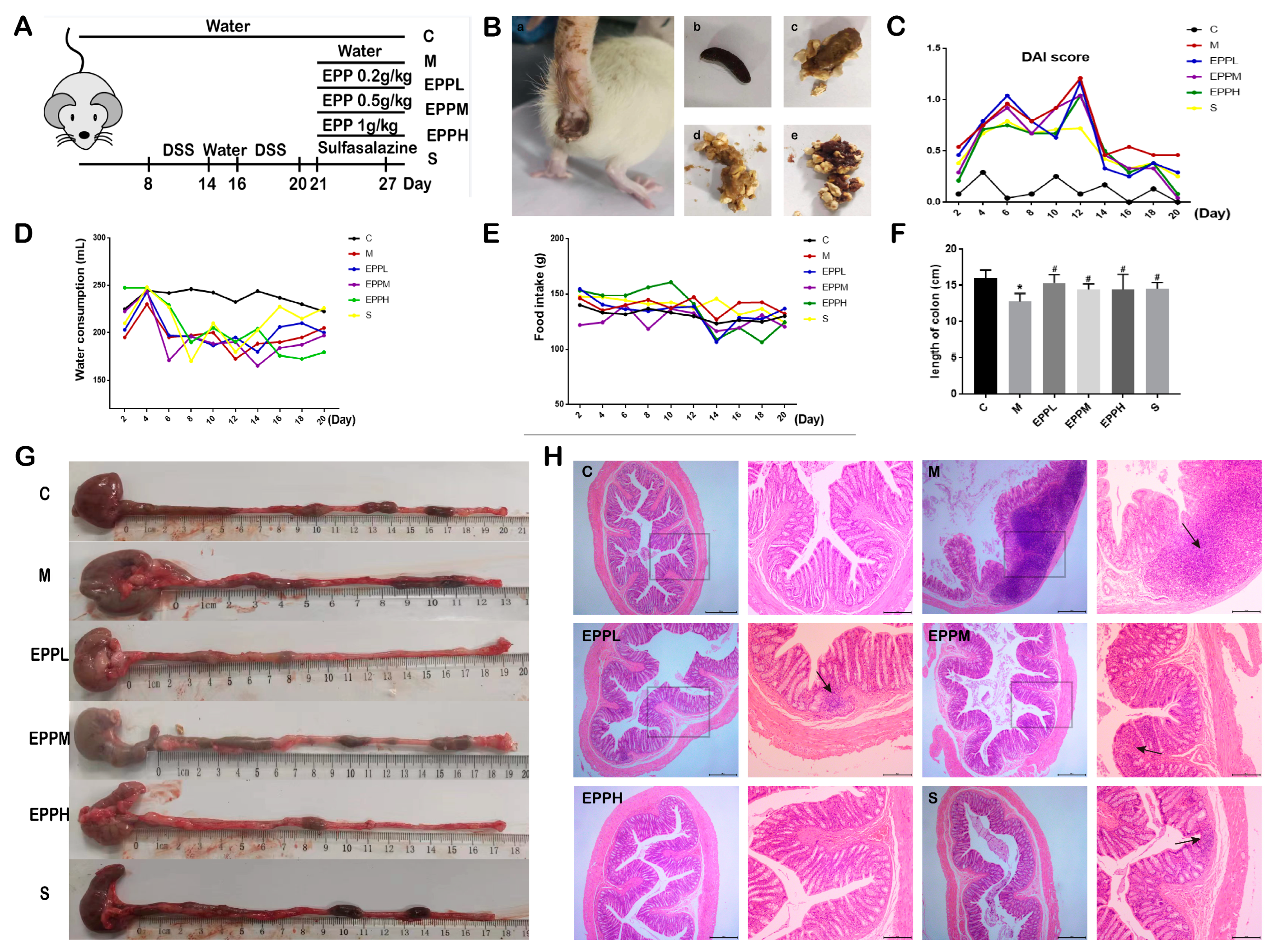
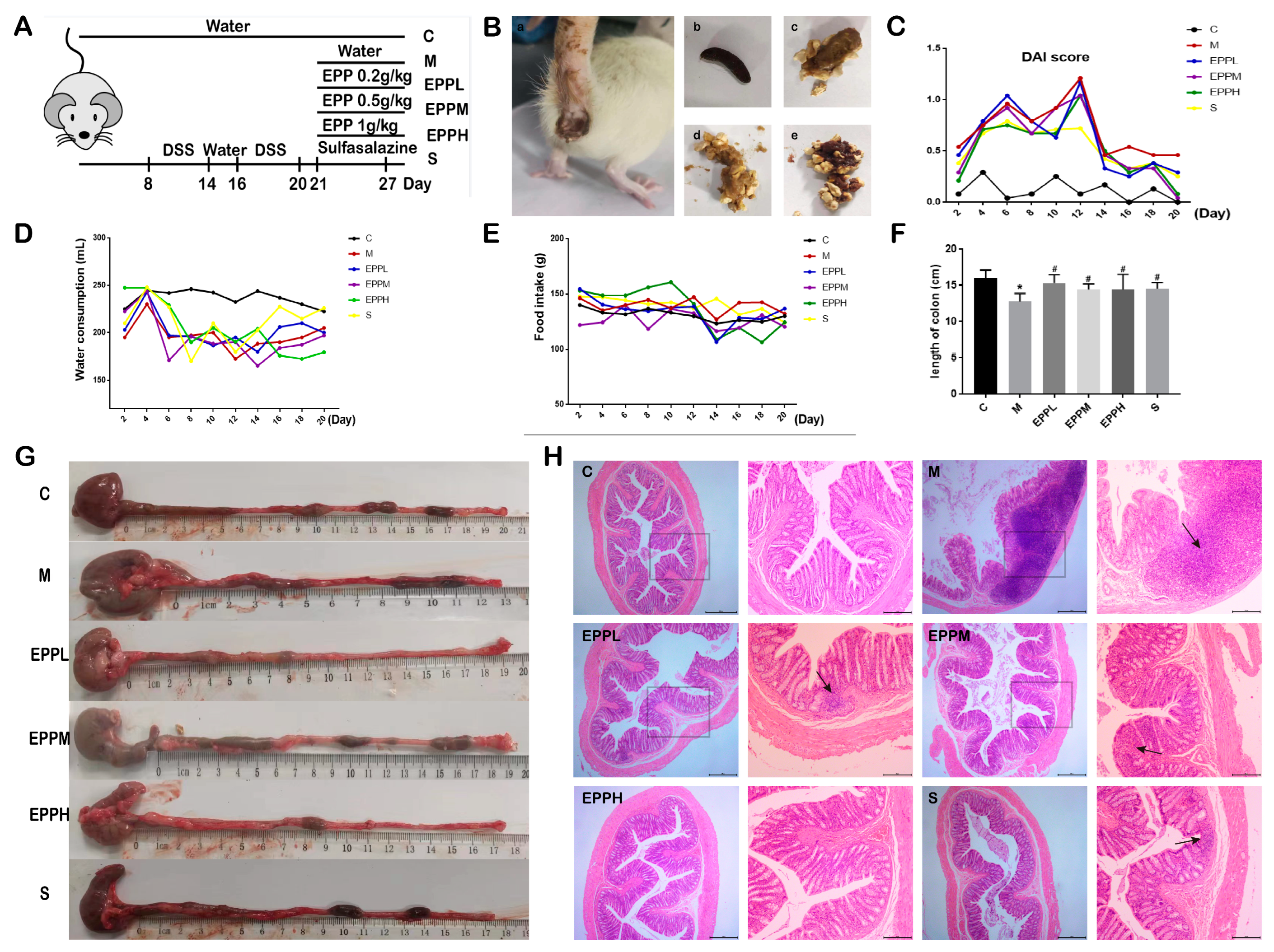
2.2. Animals Grouping and Modeling
2.3. Clinical Index Observation
2.4. Histopathology
2.5. Enzyme-Linked Immunosorbent Assay
2.6. RT-qPCR Analysis
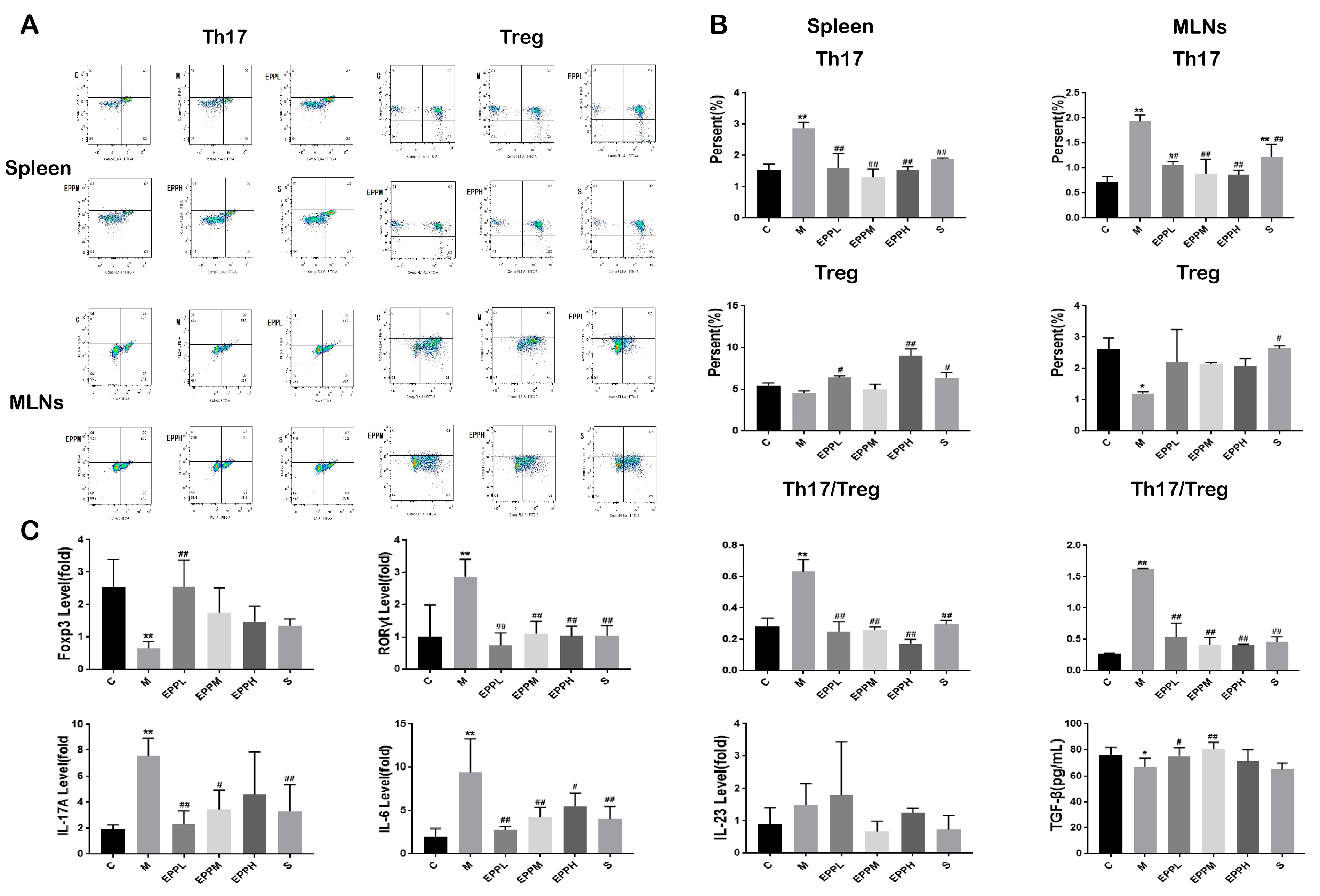
2.7. Flow Cytometry Analysis
2.8. Western Blot Analysis
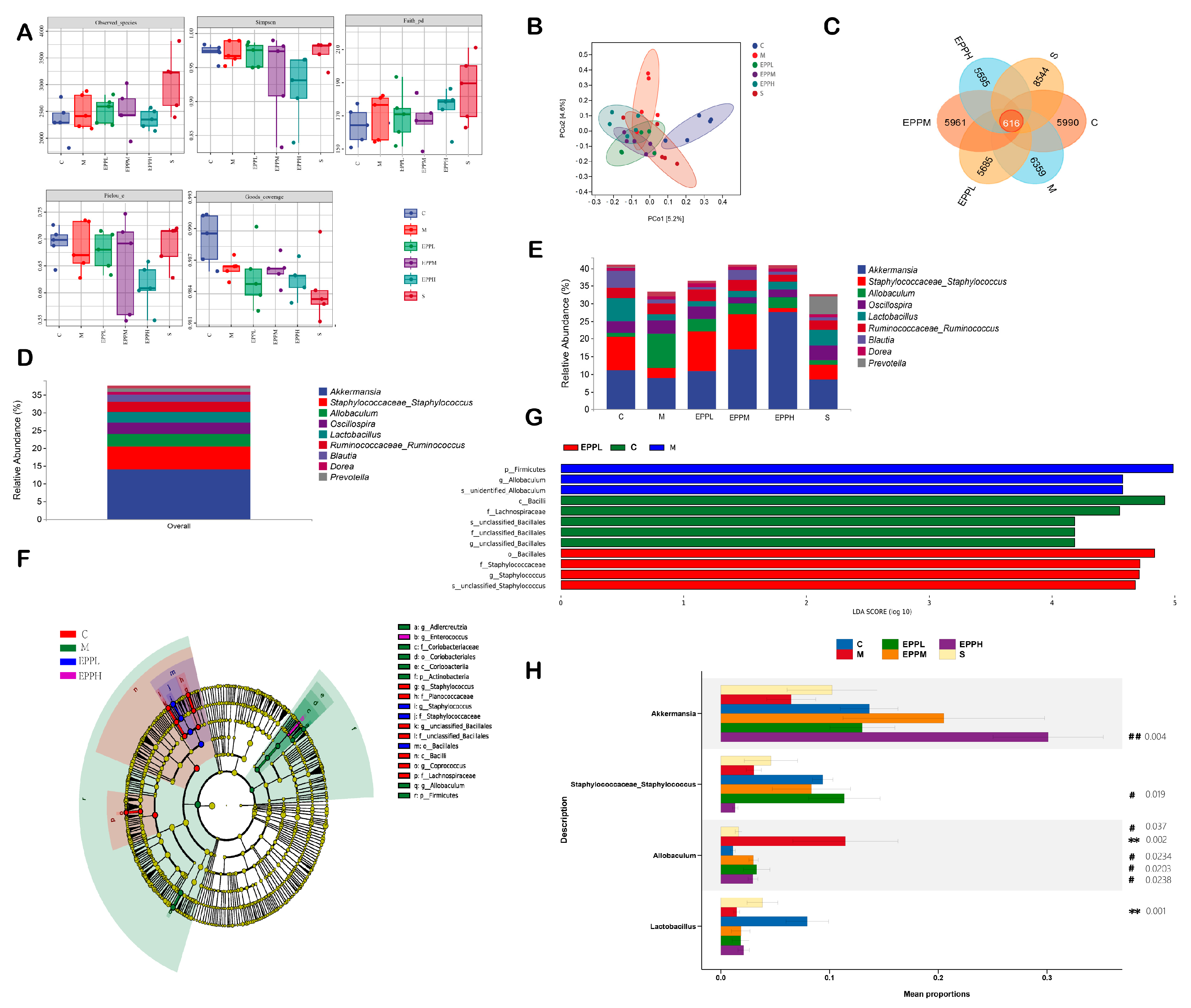
2.9. Gut Microbiota Analysis
2.10. Statistical Analysis
3. Results
3.1. EPP Improved Colitis Symptoms
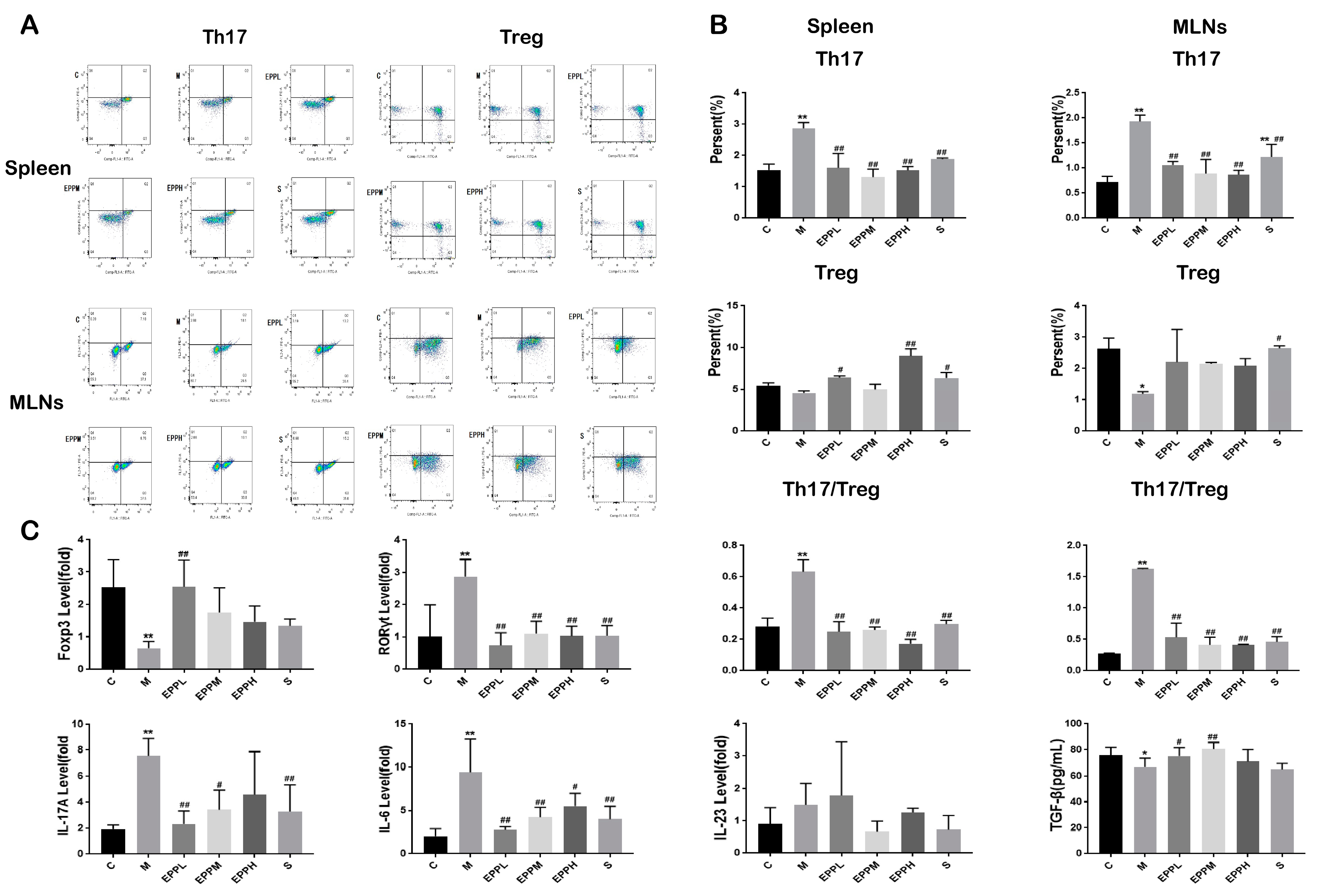
3.2. EPP Restores the Th17/Treg Balance in Colitic Rats
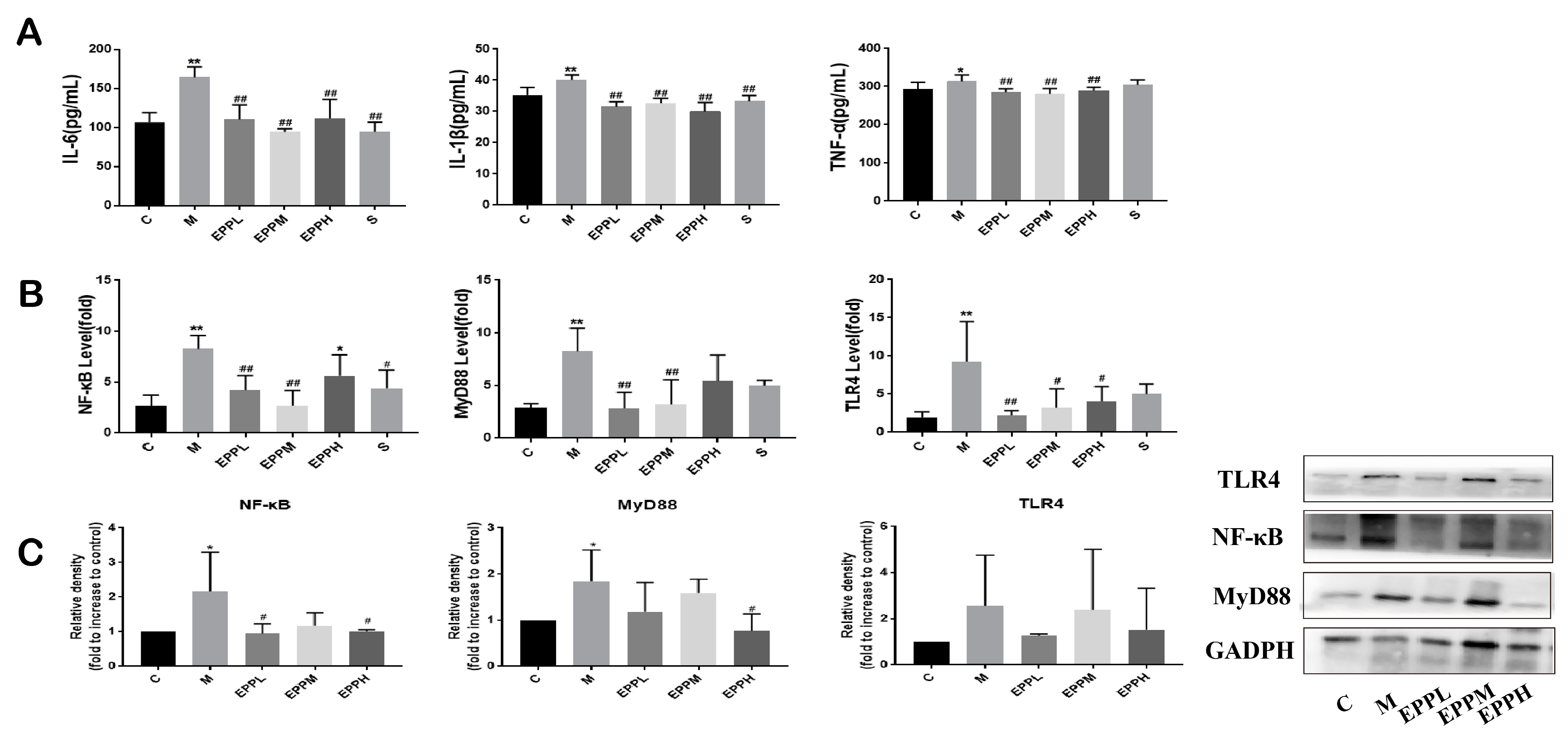
3.3. EPP Regulated Inflammatory Response
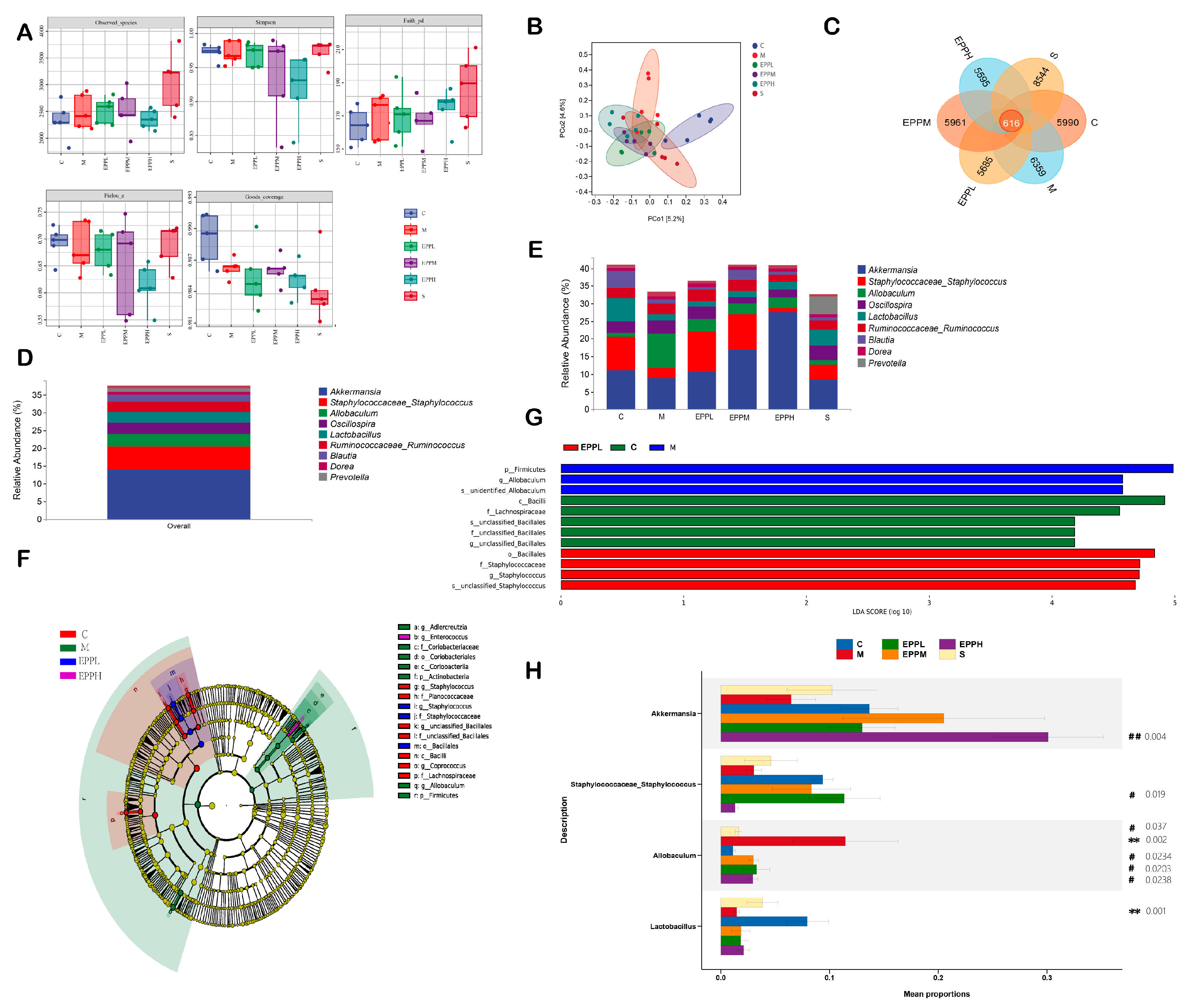
3.4. EPP Regulates Intestinal Microbial Balance in Colitis Rats
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Coskun, M.; Vermeire, S.; Nielsen, O.H. Novel Targeted Therapies for Inflammatory Bowel Disease. Trends Pharmacol. Sci. 2017, 38, 127–142. [Google Scholar] [CrossRef] [PubMed]
- Jones-Hall, Y.L.; Nakatsu, C.H. The Intersection of TNF, IBD and the Microbiome. Gut Microbes 2016, 7, 58–62. [Google Scholar] [CrossRef]
- Gaidos, J.K.; Kane, S.V. Managing IBD Therapies in Pregnancy. Curr. Treat. Options Gastroenterol. 2017, 15, 71–83. [Google Scholar] [CrossRef]
- Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar] [CrossRef]
- Littman, D.R.; Rudensky, A.Y. Th17 and regulatory T cells in mediating and restraining inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef]
- Mayne, C.G.; Williams, C.B. Induced and natural regulatory T cells in the development of inflammatory bowel disease. Inflamm. Bowel Dis. 2013, 19, 1772–1788. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wang, J. Gut microbiota and inflammatory bowel disease. Wires Mech Dis. 2022, 14, e1540. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, F.A.; Szamosi, J.C.; Rossi, L.; Griffin, L.; Nieves, K.; Bihan, D.; Lewis, I.A.; Pittman, Q.J.; Swain, M.G.; Surette, M.G.; et al. Colitis-associated microbiota drives changes in behaviour in male mice in the absence of inflammation. Brain Behav. Immun. 2022, 102, 266–278. [Google Scholar] [CrossRef]
- Lu, Y.; Li, X.; Liu, S.; Zhang, Y.; Zhang, D. Toll-like Receptors and Inflammatory Bowel Disease. Front. Immunol. 2018, 9, 72. [Google Scholar] [CrossRef] [PubMed]
- Tan, T.G.; Sefik, E.; Geva-Zatorsky, N.; Kua, L.; Naskar, D.; Teng, F.; Pasman, L.; Ortiz-Lopez, A.; Jupp, R.; Wu, H.-J.J.; et al. Identifying species of symbiont bacteria from the human gut that, alone, can induce intestinal Th17 cells in mice. Proc. Natl. Acad. Sci.USA 2016, 113, E8141–E8150. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Nunez, G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.Y.; Staniforth, V.; Chiao, M.T.; Hou, C.-C.; Wu, H.-M.; Yeh, K.-C.; Chen, C.-H.; Hwang, P.-I.; Wen, T.-N.; Shyur, L.-F.; et al. Genomics and proteomics of immune modulatory effects of a butanol fraction of echinacea purpurea in human dendritic cells. BMC Genom. 2008, 9, 479. [Google Scholar] [CrossRef]
- Sultan, M.T.; Butt, M.S.; Qayyum, M.M.; Suleria, H.A. Immunity: Plants as effective mediators. Crit. Rev. Food Sci. 2014, 54, 1298–1308. [Google Scholar] [CrossRef]
- Senchina, D.S.; Martin, A.E.; Buss, J.E.; Kohut, M.L. Effects of Echinacea extracts on macrophage antiviral activities. Phytother. Res. 2010, 24, 810–816. [Google Scholar] [CrossRef]
- Burlou-Nagy, C.; Banica, F.; Jurca, T.; Vicaș, L.G.; Marian, E.; Muresan, M.E.; Bácskay, I.; Kiss, R.; Fehér, P.; Pallag, A. Echinacea purpurea (L.) Moench: Biological and Pharmacological Properties. A Review. Plants 2022, 11, 1244. [Google Scholar] [CrossRef]
- Zhang, H.; Lang, W.; Wang, S.; Li, B.; Li, G.; Shi, Q. Echinacea polysaccharide alleviates LPS-induced lung injury via inhibiting inflammation, apoptosis and activation of the TLR4/NF-kappaB signal pathway. Int. Immunopharmacol. 2020, 88, 106974. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Zhu, H.; Xu, W.; Liu, C.; Hu, B.; Guo, Y.; Cheng, Y.; Qian, H. Echinacea purpurea polysaccharide prepared by fractional precipitation prevents alcoholic liver injury in mice by protecting the intestinal barrier and regulating liver-related pathways. Int. J. Biol. Macromol. 2021, 187, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Yu, T.; He, Y.; Chen, H.; Lu, X.; Ni, H.; Ma, Y.; Chen, Y.; Li, C.; Cao, R.; Ma, L.; et al. Polysaccharide from Echinacea purpurea plant ameliorates oxidative stress-induced liver injury by promoting Parkin-dependent autophagy. Phytomedicine 2022, 104, 154311. [Google Scholar] [CrossRef] [PubMed]
- Gu, D.; Yang, S.; Shi, D.; Guo, S.; Liu, C. Explore the mechanism of echinacea to enhance immune function based on network pharmacology. HLJ Anim. Sci. Vet. Med. 2022, 09, 136–137. [Google Scholar]
- Gu, D.; Wang, H.; Yan, M.; Li, Y.; Yang, S.; Shi, D.; Guo, S.; Wu, L.; Liu, C. Echinacea purpurea (L.) Moench extract suppresses inflammation by inhibition of C3a/C3aR signaling pathway in TNBS-induced ulcerative colitis rats. J. Ethnopharmacol. 2023, 307, 116221. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Bai, L.; Tan, Y.; Sun, J.; Qu, Q.; Shi, D.; Guo, S.; Liu, C. The immunoregulatory effect of sulfated Echinacea purpurea polysaccharide on chicken bone marrow-derived dendritic cells. Int. J. Biol. Macromol. 2019, 139, 1123–1132. [Google Scholar] [CrossRef]
- Chi, X.; Yang, S.; Yang, B.; Sun, H.; Yao, L.; Gu, D.; Guo, S.; Shi, D.; Wu, W.; Liu, C. Efficacy of Echinacea purpurea extract on damp-heat diarrhea in rats and its mechanism. Act. Vet. Zootec. Sin. 2022, 54, 380–391. [Google Scholar]
- Sun, R.; Jin, D.; Fei, F.; Xu, Z.; Cao, B.; Li, J. Mushroom polysaccharides from Grifola frondosa (Dicks.) Gray and Inonotus obliquus (Fr.) Pilat ameliorated dextran sulfate sodium-induced colitis in mice by global modulation of systemic metabolism and the gut microbiota. Front. Pharmacol. 2023, 14, 1172963. [Google Scholar] [CrossRef]
- Wang, J.; Gao, H.; Xie, Y.; Wang, P.; Li, Y.; Zhao, J.; Wang, C.; Ma, X.; Wang, Y.; Mao, Q.; et al. Lycium barbarum polysaccharide alleviates dextran sodium sulfate-induced inflammatory bowel disease by regulating M1/M2 macrophage polarization via the STAT1 and STAT6 pathways. Front Pharmacol. 2023, 14, 1044576. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T.W. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Yurkovetskiy, L.A.; Pickard, J.M.; Chervonsky, A.V. Microbiota and autoimmunity: Exploring new avenues. Cell Host Microbe 2015, 17, 548–552. [Google Scholar] [CrossRef]
- Shi, G.; Kong, J.; Wang, Y.; Xuan, Z.; Xu, F. Glycyrrhiza uralensis Fisch. alleviates dextran sulfate sodium-induced colitis in mice through inhibiting of NF-κB signaling pathways and modulating intestinal microbiota. J. Ethnopharmacol. 2022, 298, 115640. [Google Scholar] [CrossRef]
- Beukema, M.; Akkerman, R.; Jermendi, E.; Koster, T.; Laskewitz, A.; Kong, C.; Schols, H.A.; Faas, M.M.; de Vos, P. Pectins that Structurally Differ in the Distribution of Methyl-Esters Attenuate Citrobacter rodentium-Induced Colitis. Mol. Nutr. Food Res. 2021, 65, e2100346. [Google Scholar] [CrossRef]
- Hou, R.; Xu, T.; Li, Q.; Yang, F.; Wang, C.; Huang, T.; Hao, Z. Polysaccharide from Echinacea purpurea reduce the oxidant stress in vitro and in vivo. Int. J. Biol. Macromol. 2020, 149, 41–50. [Google Scholar] [CrossRef]
- Kiesler, P.; Fuss, I.J.; Strober, W. Experimental Models of Inflammatory Bowel Diseases. Cell Mol. Gastroenter. 2015, 1, 154–170. [Google Scholar] [CrossRef]
- Coleman, M.M.; Finlay, C.M.; Moran, B.; Keane, J.; Dunne, P.J.; Mills, K.H. The immunoregulatory role of CD4(+) FoxP3(+) CD25(-) regulatory T cells in lungs of mice infected with Bordetella pertussis. FEMS Immunol. Med. Microbiol. 2012, 64, 413–424. [Google Scholar] [CrossRef]
- Manel, N.; Unutmaz, D.; Littman, D.R. The differentiation of human T(H)-17 cells requires transforming growth factor-beta and induction of the nuclear receptor RORgammat. Nat. Immunol. 2008, 9, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Veldhoen, M.; Hocking, R.J.; Atkins, C.J.; Locksley, R.M.; Stockinger, B. TGFbeta in the context of an inflammatory cytokine milieu supports de novo differentiation of IL-17-producing T cells. Immunity 2006, 24, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Ivanov, I.I.; Spolski, R.; Min, R.; Shenderov, K.; Egawa, T.; E Levy, D.; Leonard, W.J.; Littman, D.R. IL-6 programs T(H)-17 cell differentiation by promoting sequential engagement of the IL-21 and IL-23 pathways. Nat. Immunol. 2007, 8, 967–974. [Google Scholar] [CrossRef] [PubMed]
- Lavelle, E.C.; Murphy, C.; O’Neill, L.A.; Creagh, E.M. The role of TLRs, NLRs, and RLRs in mucosal innate immunity and homeostasis. Mucosal. Immunol. 2010, 3, 17–28. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, L.A.; Golenbock, D.; Bowie, A.G. The history of Toll-like receptors-redefining innate immunity. Nat. Rev. Immunol. 2013, 13, 453–460. [Google Scholar] [CrossRef]
- Piccinini, A.M.; Zuliani-Alvarez, L.; Lim, J.M.; Midwood, K.S. Distinct microenvironmental cues stimulate divergent TLR4-mediated signaling pathways in macrophages. Sci. Signal. 2016, 9, a86. [Google Scholar] [CrossRef]
- Ren, W.; Ban, J.; Xia, Y.; Zhou, F.; Yuan, C.; Jia, H.; Huang, H.; Jiang, M.; Liang, M.; Li, Z.; et al. Echinacea purpurea-derived homogeneous polysaccharide exerts anti-tumor efficacy via facilitating M1 macrophage polarization. Innovation 2023, 4, 100391. [Google Scholar] [CrossRef]
- Liu, R.; Wang, H.; Zhang, Z.; Song, D.; Chen, J.; Ji, C. Advances in the study of the interaction between polysaccharides and intestinal flora. Food Sci. 2022, 43, 363–373. [Google Scholar] [CrossRef]
- Hasani, A.; Ebrahimzadeh, S.; Hemmati, F.; Khabbaz, A.; Hasani, A.; Gholizadeh, P. The role of Akkermansia muciniphila in obesity, diabetes and atherosclerosis. J. Med. Microbiol. 2021, 70, 001435. [Google Scholar] [CrossRef]
- Liu, Y.; Li, Y.; Yu, X.; Yu, L.; Tian, F.; Zhao, J.; Zhang, H.; Zhai, Q.; Chen, W. Physiological Characteristics of Lactobacillus casei Strains and Their Alleviation Effects against Inflammatory Bowel Disease. J. Microbiol. Biotechn. 2021, 31, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Dong, Y.; Shen, W.; Du, J.; Sun, Q.; Yang, Y.; Yin, D. Platycodon grandiflorus polysaccharide regulates colonic immunity through mesenteric lymphatic circulation to attenuate ulcerative colitis. Chin. J. Nat. Med. 2023, 21, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Aarland, R.C.; Bañuelos-Hernández, A.E.; Fragoso-Serrano, M.; Sierra-Palacios, E.d.C.; de León-Sánchez, F.D.; Pérez-Flores, L.J.; Rivera-Cabrera, F.; Mendoza-Espinoza, J.A. Studies on phytochemical, antioxidant, anti-inflammatory, hypoglycaemic and antiproliferative activities of Echinacea purpurea and Echinacea angustifolia extracts. Pharm. Biol. 2017, 55, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Garand, M.; Al Khodor, S. Integrating omics for a better understanding of Inflammatory Bowel Disease: A step towards personalized medicine. J. Transl. Med. 2019, 17, 419. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Score | Percentage of Weight Loss (%) | Fecal Viscosity | Fecal Occult Blood |
|---|---|---|---|
| 0 | <1 | Black, hard, shaped | / |
| 1 | 1~5 | Black, soft and formed | Indicator is lilac |
| 2 | 6~10 | Yellow, soft, half formed | Indicator purple |
| 3 | 11~20 | Yellow, soft, shapeless | Indicator dark purple |
| 4 | >20 | Yellow, watery stool | Blood fecal |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Lin, Y.; Zheng, X.; Zheng, X.; Yan, M.; Wang, H.; Liu, C. Echinacea purpurea (L.) Moench Polysaccharide Alleviates DSS-Induced Colitis in Rats by Restoring Th17/Treg Balance and Regulating Intestinal Flora. Foods 2023, 12, 4265. https://doi.org/10.3390/foods12234265
Li Y, Lin Y, Zheng X, Zheng X, Yan M, Wang H, Liu C. Echinacea purpurea (L.) Moench Polysaccharide Alleviates DSS-Induced Colitis in Rats by Restoring Th17/Treg Balance and Regulating Intestinal Flora. Foods. 2023; 12(23):4265. https://doi.org/10.3390/foods12234265
Chicago/Turabian StyleLi, Yaoxing, Yongshi Lin, Xirui Zheng, Xiaoman Zheng, Mingen Yan, Huiting Wang, and Cui Liu. 2023. "Echinacea purpurea (L.) Moench Polysaccharide Alleviates DSS-Induced Colitis in Rats by Restoring Th17/Treg Balance and Regulating Intestinal Flora" Foods 12, no. 23: 4265. https://doi.org/10.3390/foods12234265
APA StyleLi, Y., Lin, Y., Zheng, X., Zheng, X., Yan, M., Wang, H., & Liu, C. (2023). Echinacea purpurea (L.) Moench Polysaccharide Alleviates DSS-Induced Colitis in Rats by Restoring Th17/Treg Balance and Regulating Intestinal Flora. Foods, 12(23), 4265. https://doi.org/10.3390/foods12234265