
Effect of Red Light on the Expression of the Phytochrome Gene Family and the Accumulation of Glycoside Alkaloids in Potatoes




Abstract
:
1. Introduction
2. Materials and Methods
2.1. Preparation of Plant Materials
2.2. Light Irritation Treatment and Sampling
2.3. Extraction and Detection of GAs
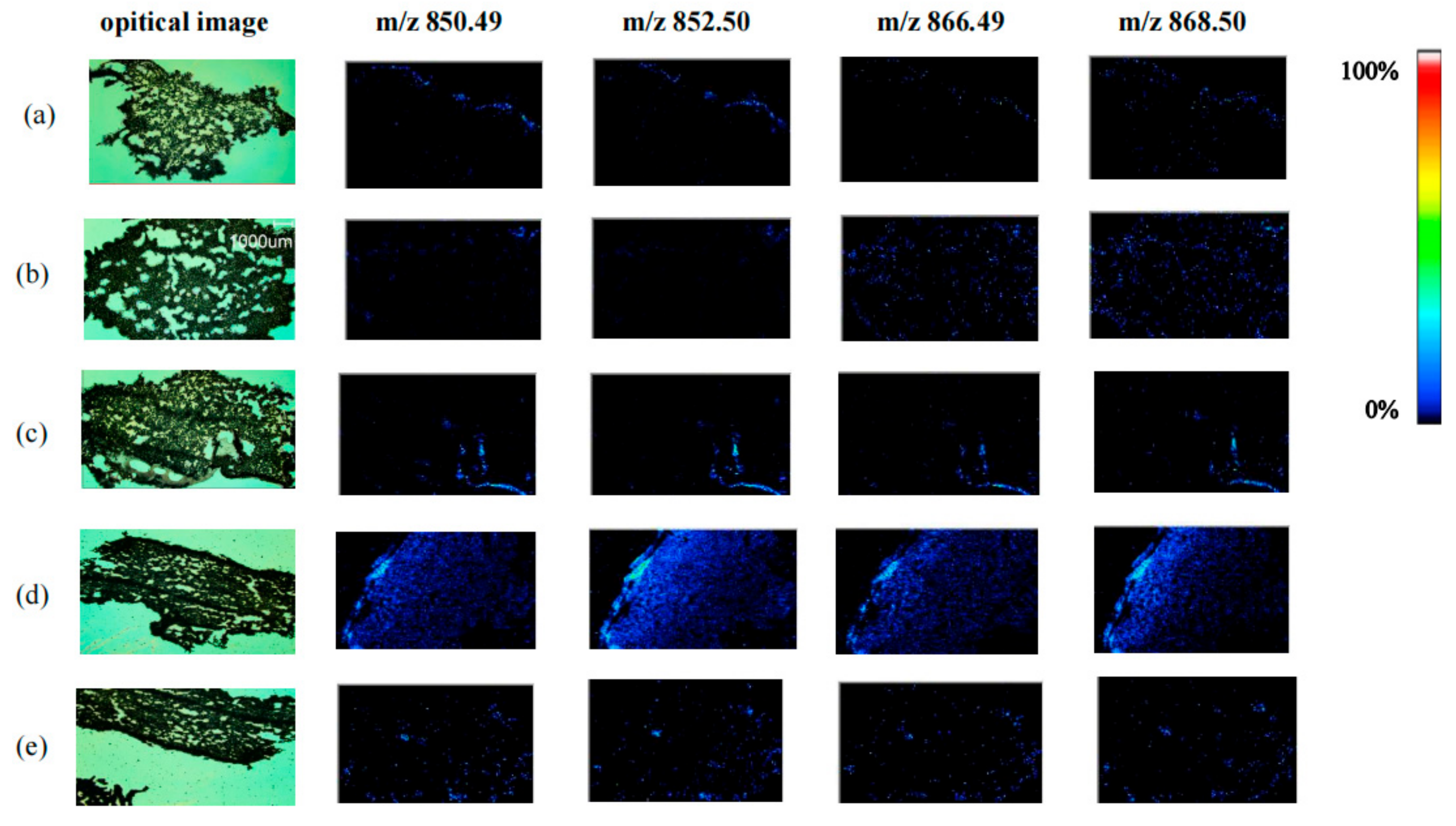
2.4. GA Observation via an Imaging Mass Microscope
2.5. Identification and Analysis of StPHY Genes
2.6. The Expression Pattern of StPHYs Genes
2.7. Statistical Analysis
3. Results
3.1. GA Analysis by Mass Spectrometry Imaging (MSI)
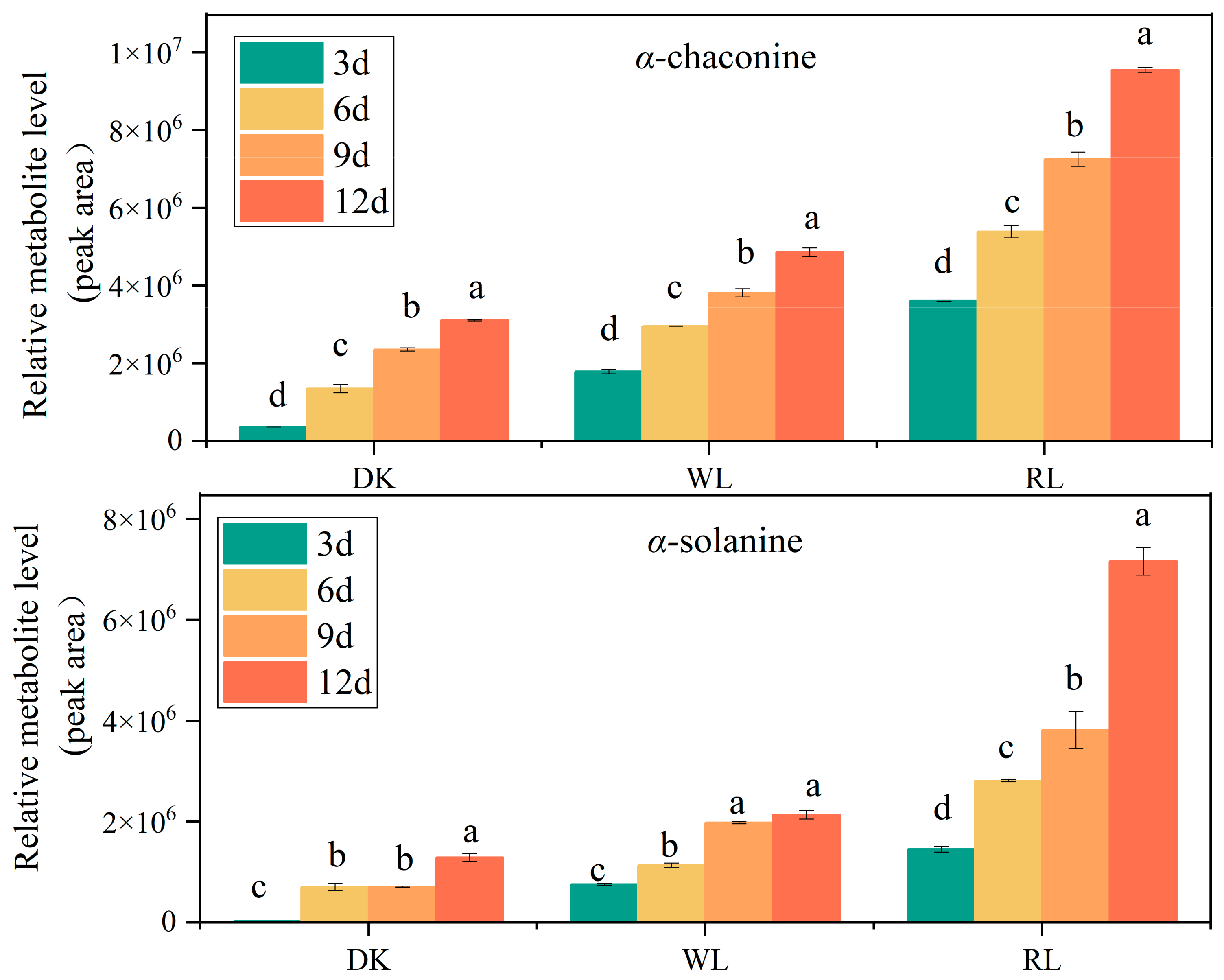
3.2. GA Detection in Potato Tubers via HPLC
3.3. Identification of Potato PHYs
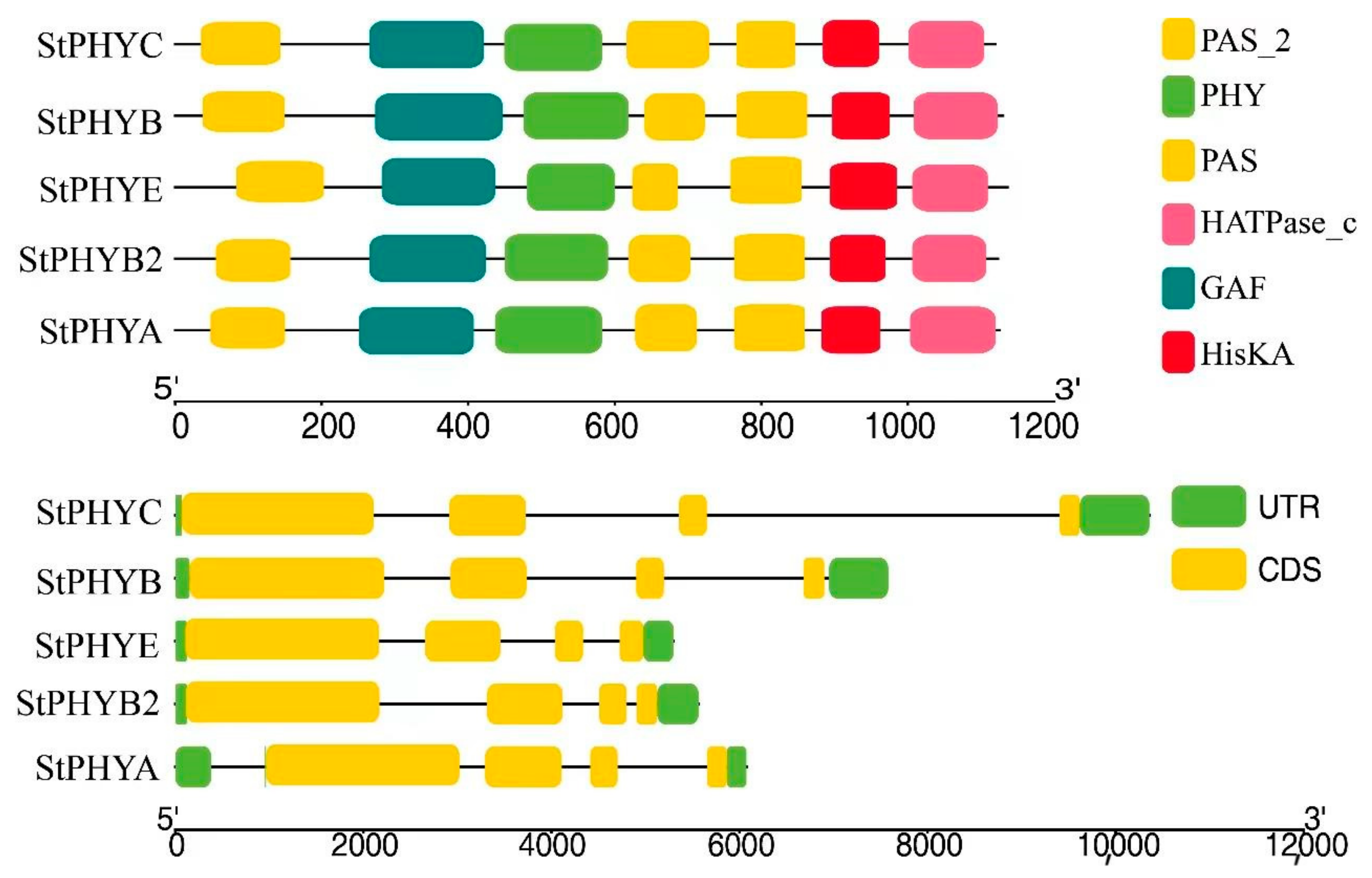
3.4. Analysis of StPHYs Gene Structure
3.5. Phylogenetic Analysis of StPHYs
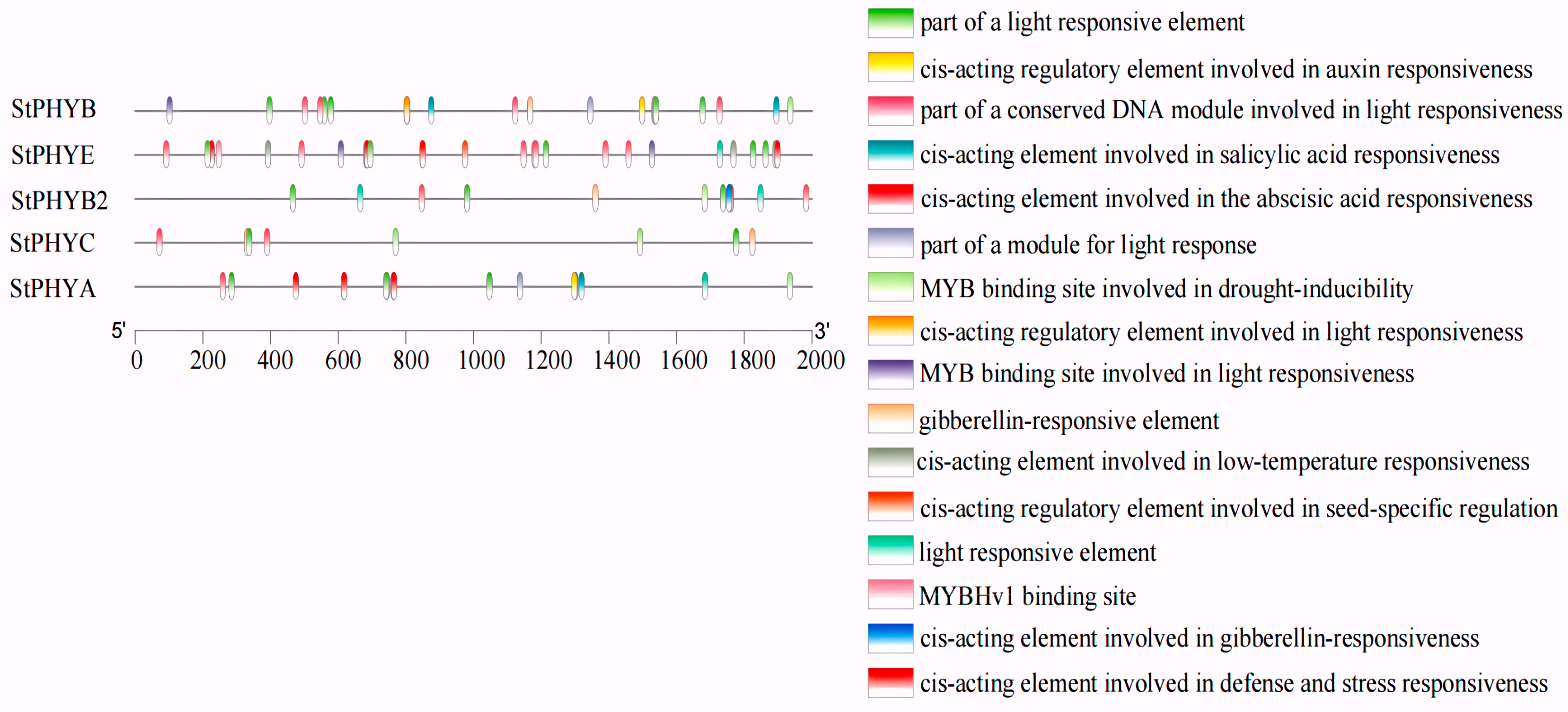
3.6. Promoter Cis-Acting Element Analysis of StPHYs
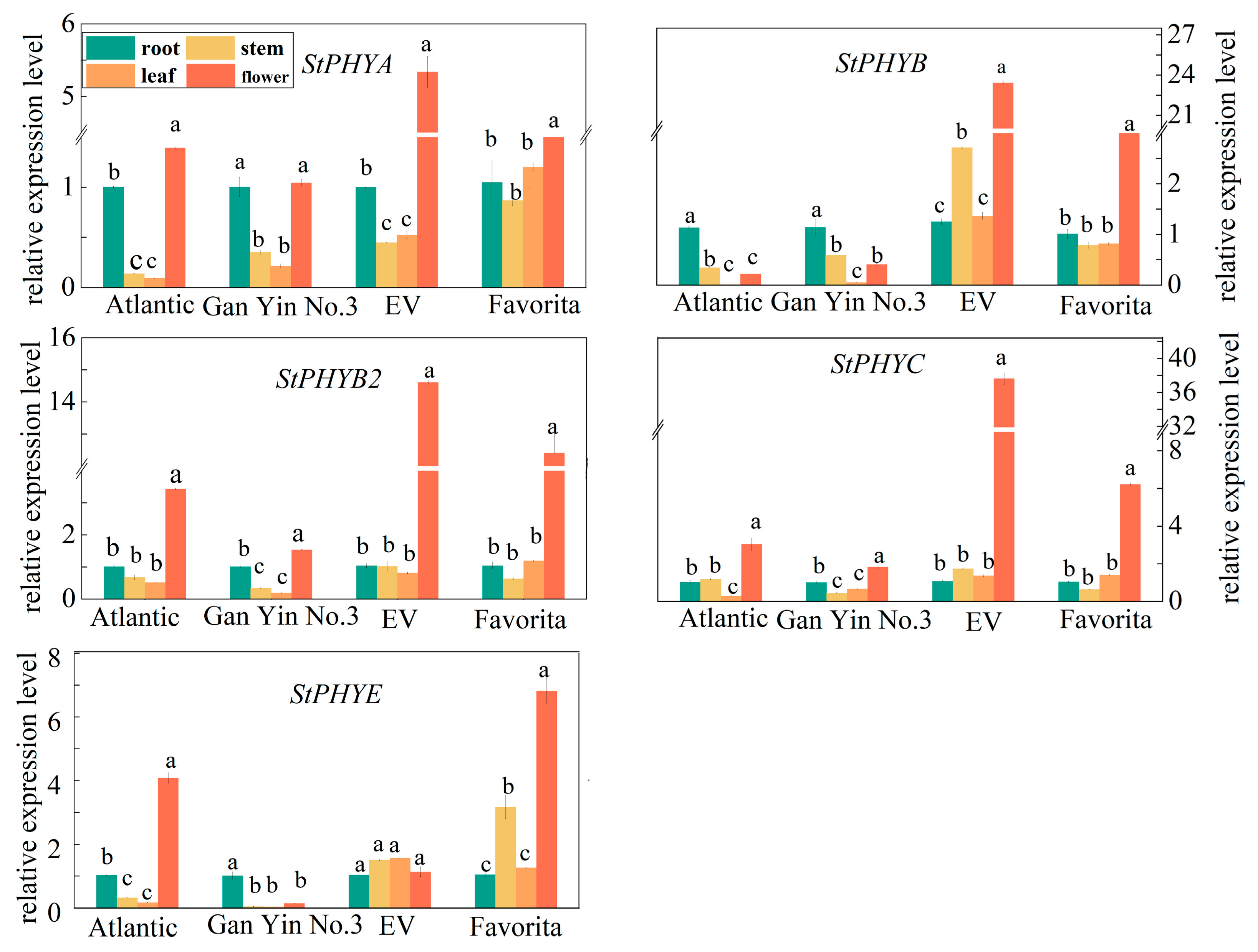
3.7. Expression Analysis of StPHYs Genes in Different Organs from Different Potato Varieties
3.8. Expression Analysis of StPHYs Genes under Different Lights
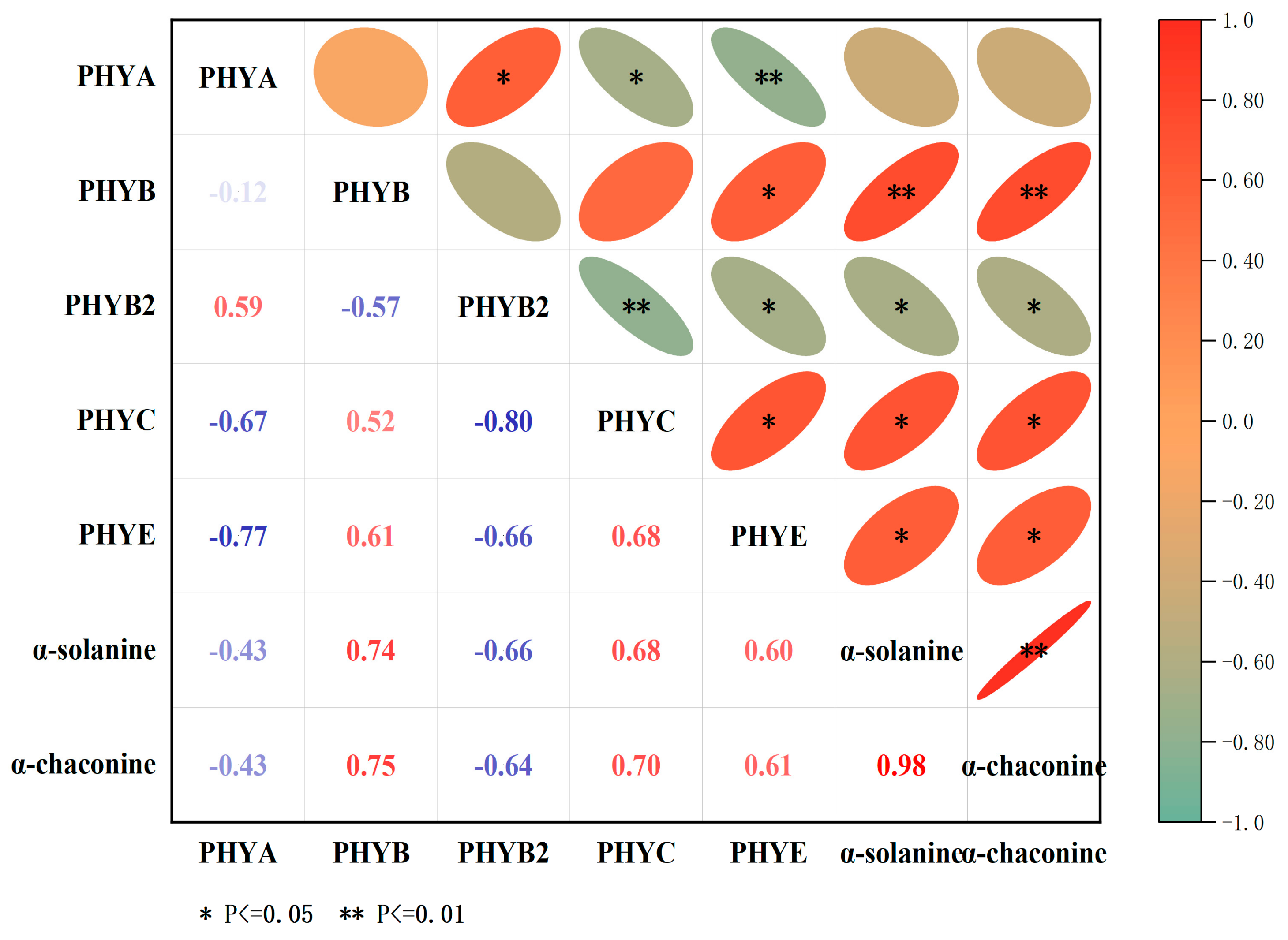
3.9. Correlation Analysis between StPHYs Expression and GA Contents
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mishra, S.; Monro, J.; Hedderley, D. Effect of processing on slowly digestible starch and resistant starch in potato. Starch-Stärke 2008, 60, 500–507. [Google Scholar] [CrossRef]
- Ek, K.L.; Wang, S.; Copeland, L.; Brand-Miller, J.C. Discovery of a low-glycaemic index potato and relationship with starch digestion in vitro. Br. J. Nutr. 2014, 111, 699–705. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Zhou, Y.; Yuan, Y.; Fan, Z.; Wu, Y.; Liu, A.; Lu, X. Potato preload mitigated postprandial glycemic excursion in healthy subjects: An Acute Randomized Trial. Nutrients 2020, 12, 2759. [Google Scholar] [CrossRef]
- Wang, C.C.; Meng, L.H.; Gao, Y.; Grierson, D.; Fu, D.Q. Manipulation of light signal transduction factors as a means of modifying Steroidal Glycoalkaloids accumulation in tomato leaves. Front. Plant Sci. 2018, 9, 437. [Google Scholar] [CrossRef]
- Milner, S.E.; Brunton, N.P.; Jones, P.W.; O’Brien, N.M.; Collins, S.G.; Maguire, A.R. bioactivities of glycoalkaloids and their aglycones from Solanum species. J. Agric. Food Chem. 2011, 59, 3454–3484. [Google Scholar] [CrossRef]
- Keukens, E.A.; Vrije, T.; van den Boom, C.; de Waard, P.; Plasman, H.H.; Thiel, F.; Chupin, V.; Jongen, W.M.F.; de Kruijff, B. Molecular basis of glycoalkaloid induced membrane disruption. Biochim. Biophys. Acta 1995, 1240, 216–228. [Google Scholar] [CrossRef]
- Huber, M.; Nieuwendijk, N.M.; Pantazopoulou, C.K.; Pierik, R. Light signalling shapes plant-plant interactions in dense canopies. Plant Cell Environ. 2021, 44, 1014–1029. [Google Scholar] [CrossRef]
- Zhu, Y.; Tepperman, J.M.; Fairchild, C.D.; Quail, P.H. Phytochrome B binds with greater apparent affinity than phytochrome A to the basic helix-loop-helix factor PIF3 in a reaction requiring the PAS domain of PIF3. Proc. Natl. Acad. Sci. USA 2000, 97, 13419–13424. [Google Scholar] [CrossRef]
- Shen, Y.; Khanna, R.; Carle, C.M.; Quail, P.H. Phytochrome induces rapid PIF5 phosphorylation and degradation in response to red-light activation. Plant Physiol. 2007, 145, 1043–1051. [Google Scholar] [CrossRef]
- Roig-Villanova, I.; Bou, J.; Sorin, C.; Devlin, P.F.; Martínez-García, J.F. Identification of primary target genes of phytochrome signaling. Early transcriptional control during shade avoidance responses in Arabidopsis. Plant Physiol. 2006, 141, 85–96. [Google Scholar] [CrossRef]
- Zhu, L.; Huq, E. Characterization of light-regulated protein-protein interactions by in vivo coimmunoprecipitation (Co-IP) assays in plants. Methods Mol. Biol. 2019, 2026, 29–39. [Google Scholar] [PubMed]
- Carlson, K.D.; Bhogale, S.; Anderson, D.; Tomanek, L.; Madlung, A. Phytochrome a regulates carbon flux in dark grown tomato seedlings. Front. Plant Sci. 2019, 10, 152. [Google Scholar] [CrossRef]
- Kozuka, T.; Sawada, Y.; Imai, H.; Kanai, M.; Hirai, M.Y.; Mano, S.; Uemura, M.; Nishimura, M.; Kusaba, M.; Nagatani, A. Regulation of sugar and storage oil metabolism by phytochrome during De-etiolation. Plant Physiol. 2020, 182, 1114–1129. [Google Scholar] [CrossRef] [PubMed]
- Bancos, S.; Szatmari, A.M.; Castle, J.; Kozma-Bognar, L.; Shibata, K.; Yokota, T. Diurnal regulation of the brassinosteroid-biosynthetic CPD gene in Arabidopsis. Plant Physiol. 2006, 141, 299–309. [Google Scholar] [CrossRef]
- Stephenson, P.G.; Terry, M.J. Light signalling pathways regulating the Mg-chelatase branchpoint of chlorophyll synthesis during de-etiolation in Arabidopsis thaliana. Photochem. Photobiol. 2008, 7, 1243–1252. [Google Scholar] [CrossRef] [PubMed]
- Baur, S.; Bellé, N.; Hausladen, H.; Wurzer, S.; Brehm, L.; Stark, T.D.; Hücklhoven, R. Quantitation of toxic steroidal glycoalkaloids and newly identified saponins in post-harvest light-stressed potato (Solanum tuberosum L.) varieties. J. Agric. Food Chem. 2022, 70, 8300–8308. [Google Scholar] [CrossRef] [PubMed]
- Zeng, F.; Zhou, T.; Kang, X. Determination of glycoalkaloids in potato tuber by HPLC. Chin. Potato J. 2015, 29, 263–268. [Google Scholar]
- Deng, Y.; He, M.; Feng, F.; Feng, X.; Zhang, Y.; Zhang, F. The distribution and changes of glycoalkaloids in potato tubers under different storage time based on MALDI-TOF mass spectrometry imaging. Talanta 2021, 221, 121453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.; Zheng, T.; Zhu, X.; Jiu, S.; Liu, Z.; Guan, L.; Jia, H.; Fang, J. Genome-wide identification of PIFs in grapes (Vitis vinifera L.) and their transcriptional analysis under lighting/shading conditions. Genes 2018, 9, 451. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dhalsamant, K.; Singh, C.B.; Lankapalli, R.A. Review on greening and glycoalkaloids in potato tubers: Potential solutions. J. Agric. Food Chem. 2022, 70, 13819–13831. [Google Scholar] [CrossRef]
- Mekapogu, M.; Sohn, H.B.; Kim, S.J.; Lee, Y.-Y.; Park, H.-M.; Jin, Y.-I.; Hong, S.-Y.; Suh, J.-T.; Kweon, K.; Jeong, J.-C.; et al. Effect of light quality on the expression of glycoalkaloid biosynthetic genes contributing to steroidal glycoalkaloid accumulation in potato. Am. J. Potato Res. 2016, 93, 264–277. [Google Scholar] [CrossRef]
- Okamoto, H.; Ducreux, L.J.M.; Allwood, J.W.; Hedley, P.E.; Wright, A.; Gururajan, V.; Terry, M.J.; Taylor, M.A. Light regulation of chlorophyll and glycoalkaloid biosynthesis during tuber greening of potato S. tuberosum. Front. Plant Sci. 2020, 11, 753. [Google Scholar] [CrossRef]
- Yu, W.W.; Liu, Y.; Song, L.; Jacobs, D.F.; Du, X.; Ying, Y.; Shao, Q.; Wu, J. Effect of differential light quality on morphology, photosynthesis, and antioxidant enzyme activity in camptotheca acuminata seedlings. J. Plant Growth Regul. 2017, 36, 148–160. [Google Scholar] [CrossRef]
- Rymuza, K.; Gugała, M.; Zarzecka, K.; Sikorska, A.; Findura, P.; Malaga-Toboła, U.; Kapela, K.; Radzka, E. The effect of light exposures on the content of harmful substances in edible potato tuber. Agriculture 2020, 10, 139. [Google Scholar] [CrossRef]
- Nitithamyong, A.; Vonelbe, J.H.; Wheeler, R.M.; Tibbitts, T.W. Glycoalkaloids in potato tubers grown under controlled environments. Am. J. Potato Res. 1999, 76, 337–343. [Google Scholar] [CrossRef]
- Xiong, Y.; Liu, X.; You, Q.; Han, L.; Shi, J.; Yang, J.; Cui, W.; Zhang, H.; Chao, Q.; Zhu, Y.; et al. Analysis of DNA methylation in potato tuber in response to light exposure during storage. Plant Physiol. Biochem. 2022, 170, 218–224. [Google Scholar] [CrossRef]
- Friedman, M. Potato glycoalkaloids and metabolites: Roles in the plant and in the diet. J. Agric. Food Chem. 2006, 54, 8655–8681. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, J.W.; Wang, D.; Tao, S.-H.; Ji, Y.-L.; Wu, B. Relation between light qualities and accumulation of steroidal glycoalkaloids as well as signal molecule in cell in potato tubers. Acta Agron. Sin. 2010, 36, 629–635. [Google Scholar] [CrossRef]
- Sharrock, R.A.; Quail, P.H. Novel phytochrome sequences in Arabidopsis thaliana: Structure, evolution, and differential expression of a plant regulatory photoreceptor family. Genes Dev. 1989, 3, 1745–1757. [Google Scholar] [CrossRef]
- Wahlgren, W.Y.; Claesson, E.; Tuure, I. Structural mechanism of signal transduction in a phytochrome histidine kinase. Nat. Commun. 2022, 13, 7673. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Melkonian, M.; Rothfels, C.J.; Villarreal, J.C.; Stevenson, D.W.; Graham, S.W.; Wong, G.K.-S.; Pryer, K.M.; Mathews, S. Phytochrome diversity in green plants and the origin of canonical plant phytochromes. Nat. Commun. 2015, 6, 7852. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Dai, S. Advances in photoreceptor-mediated signaling transduction in flowering time regulation. Chin. Bull. Bot. 2019, 54, 9–22. [Google Scholar]
- Jiang, M.; Bin, J.; Pan, R. Effect of light quality on the primary nitrogen assimination of rice (Oryza sativa L.) seedlings. J. Integr. Plant Biol. 2000, 42, 234–238. [Google Scholar]
- Kami, C.; Lorrain, S.; Hornitschek, P.; Fankhauser, C. Chapter two—Light-regulated plant growth and development. Curr. Top. Dev. Biol. 2010, 91, 29–66. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence ID | Number of Amino Acid | Molecular Weight | pI | Instability Index | GRAVY | Subcellular Localization |
|---|---|---|---|---|---|---|
| PHYC | 1120 | 124,731.52 | 5.89 | 47.76 | −0.111 | Nucleus |
| PHYB | 1130 | 125,364.58 | 5.68 | 43.98 | −0.141 | Nucleus |
| PHYE | 704 | 77,442.38 | 5.84 | 53.61 | −0.171 | Nucleus |
| PHYB2 | 1125 | 125,782.06 | 5.75 | 46.74 | −0.145 | Nucleus |
| PHYA | 1123 | 124,671.98 | 5.87 | 42.05 | −0.147 | Nucleus |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Jiang, H.; Liu, W.; Wang, Y.; Zeng, F. Effect of Red Light on the Expression of the Phytochrome Gene Family and the Accumulation of Glycoside Alkaloids in Potatoes. Foods 2023, 12, 4194. https://doi.org/10.3390/foods12234194
Zhang X, Jiang H, Liu W, Wang Y, Zeng F. Effect of Red Light on the Expression of the Phytochrome Gene Family and the Accumulation of Glycoside Alkaloids in Potatoes. Foods. 2023; 12(23):4194. https://doi.org/10.3390/foods12234194
Chicago/Turabian StyleZhang, Xiaolu, Hong Jiang, Weigang Liu, Ya Wang, and Fankui Zeng. 2023. "Effect of Red Light on the Expression of the Phytochrome Gene Family and the Accumulation of Glycoside Alkaloids in Potatoes" Foods 12, no. 23: 4194. https://doi.org/10.3390/foods12234194
APA StyleZhang, X., Jiang, H., Liu, W., Wang, Y., & Zeng, F. (2023). Effect of Red Light on the Expression of the Phytochrome Gene Family and the Accumulation of Glycoside Alkaloids in Potatoes. Foods, 12(23), 4194. https://doi.org/10.3390/foods12234194