




Metric and Spectral Insight into Bee-Pollen-to-Bee-Bread Transformation Process

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sampling of Bee Pollen and Bee Bread
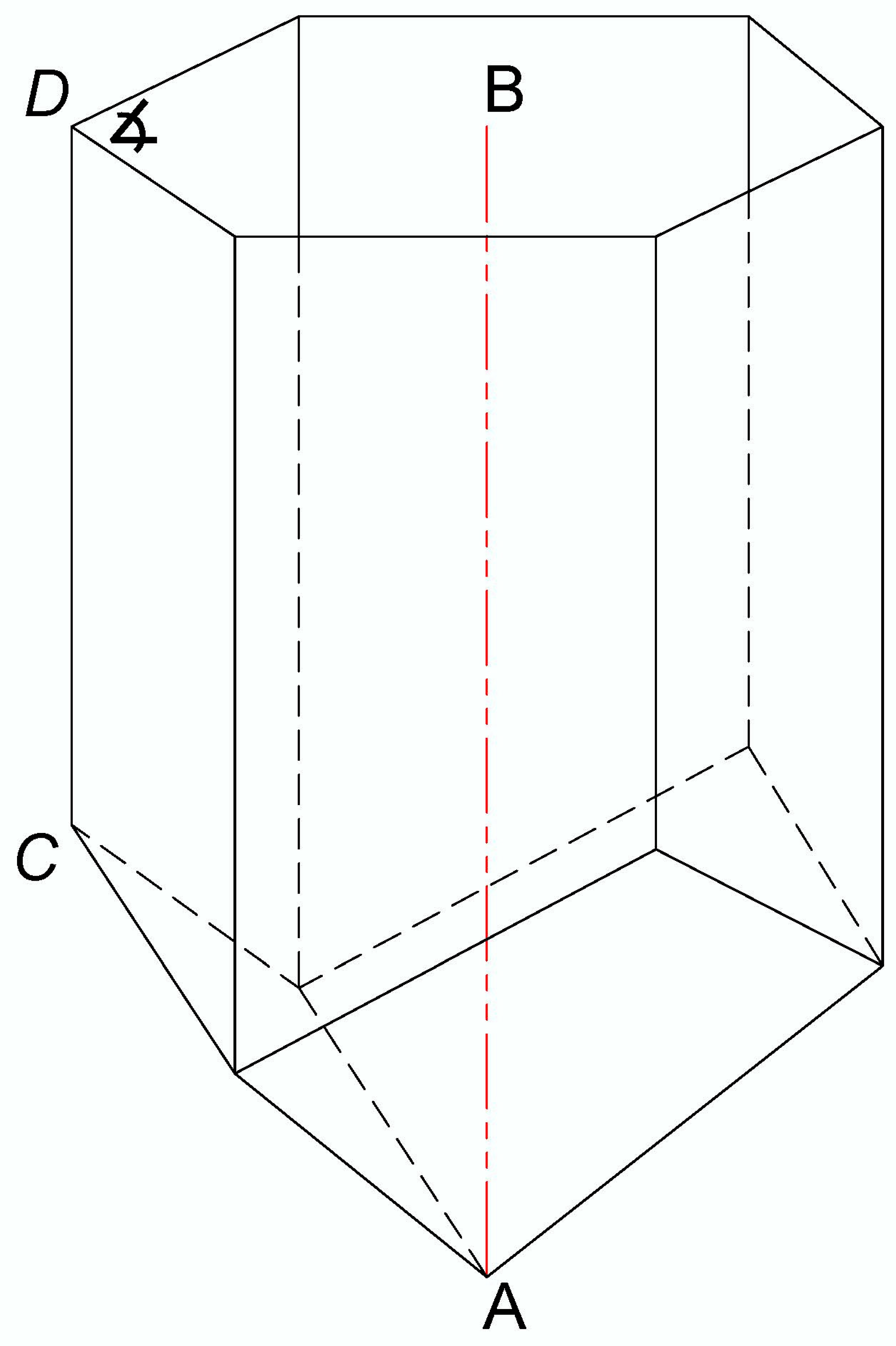
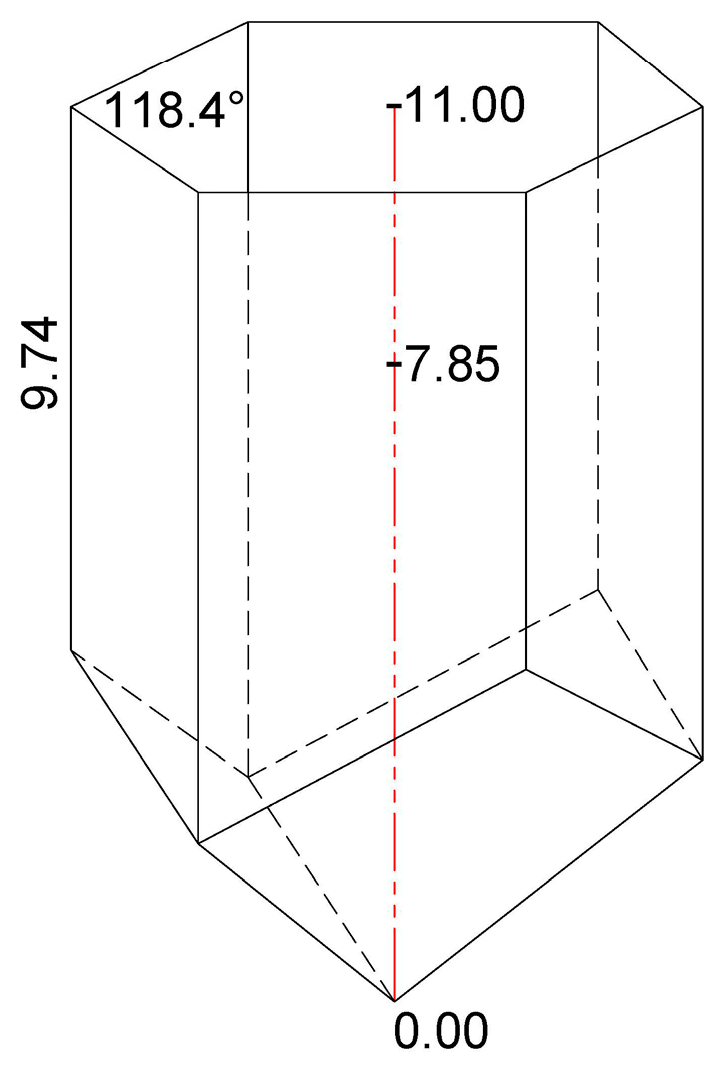
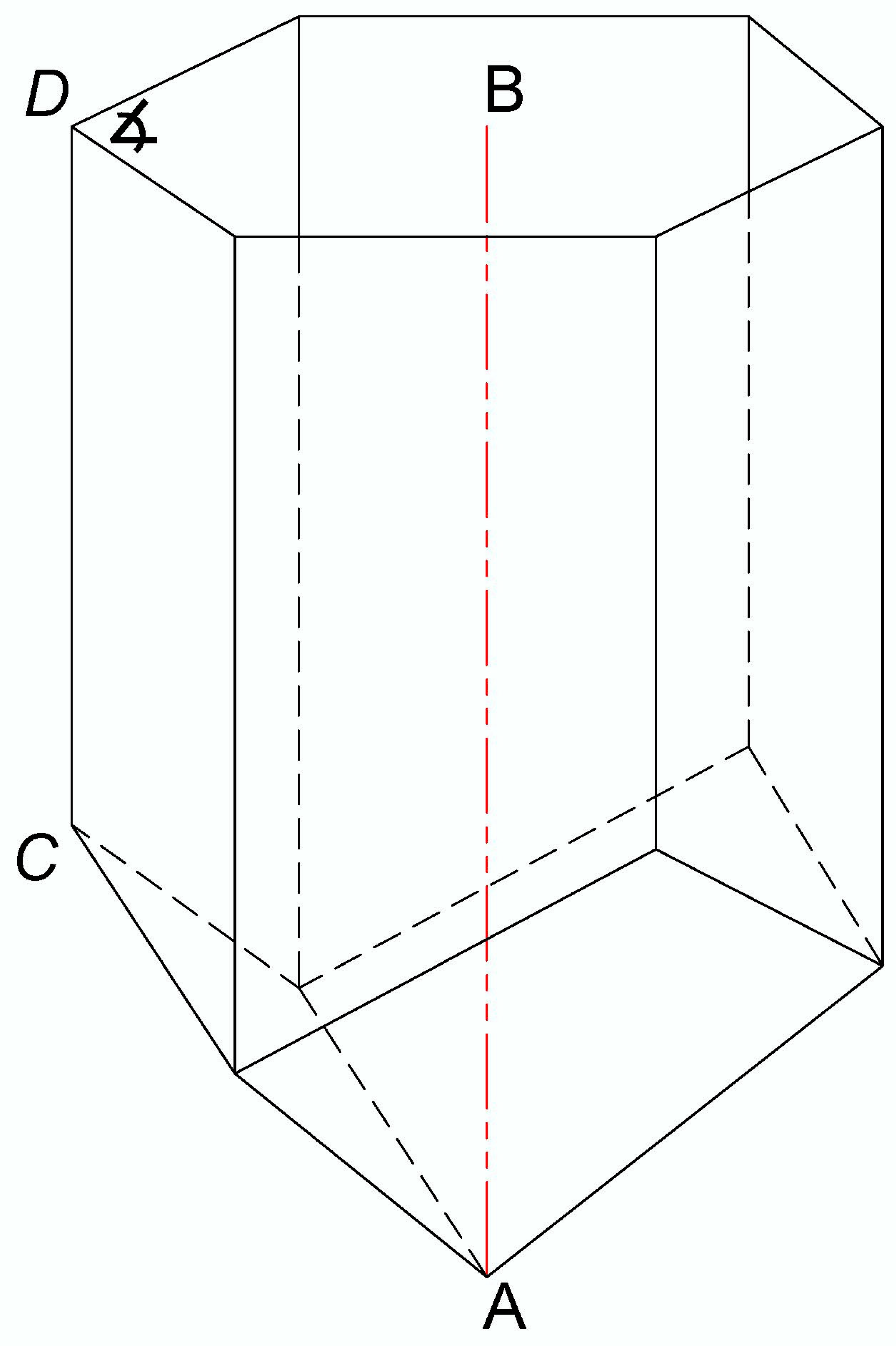
2.3. Determination of the Cell Depth of Bee Bread Storage and Complementary Cell Measurements
2.4. Statistical Analysis
2.5. FTIR-ATR Spectroscopy and Spectral Data Analysis
2.6. Freeze-Drying (Lyophilisation) of BP and BB Samples
3. Results and Discussion
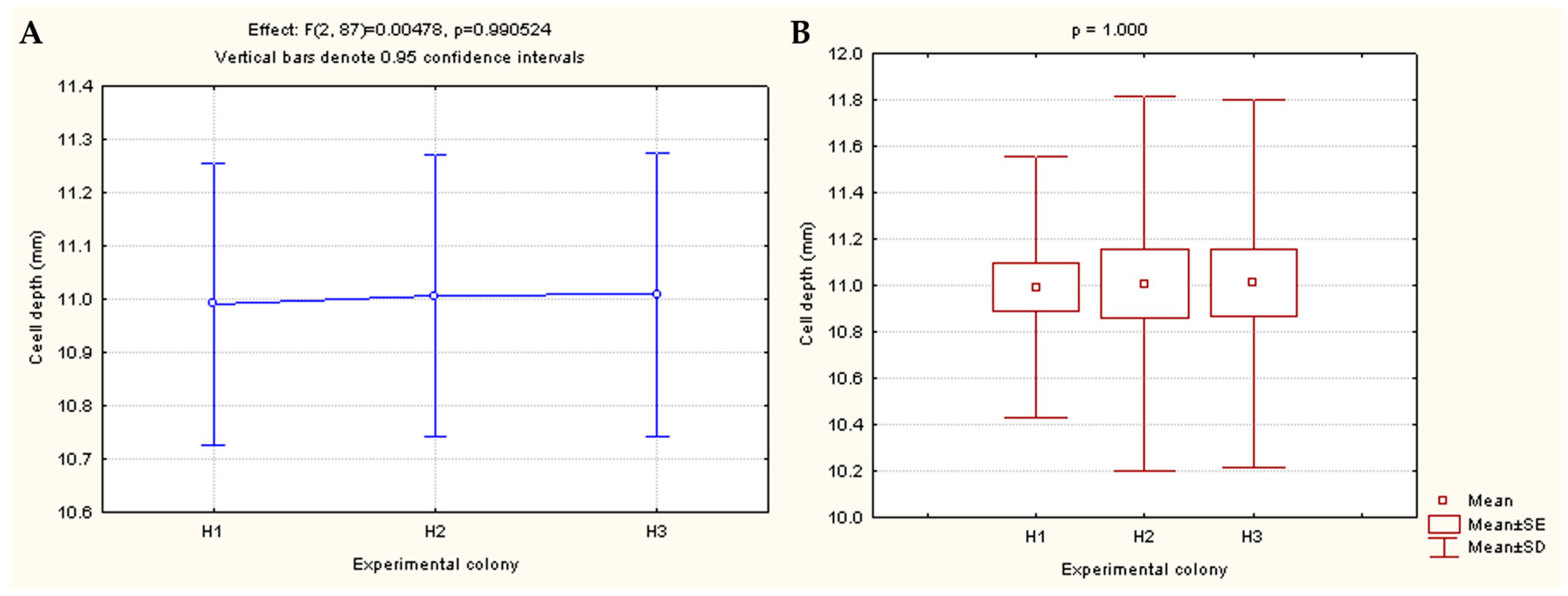
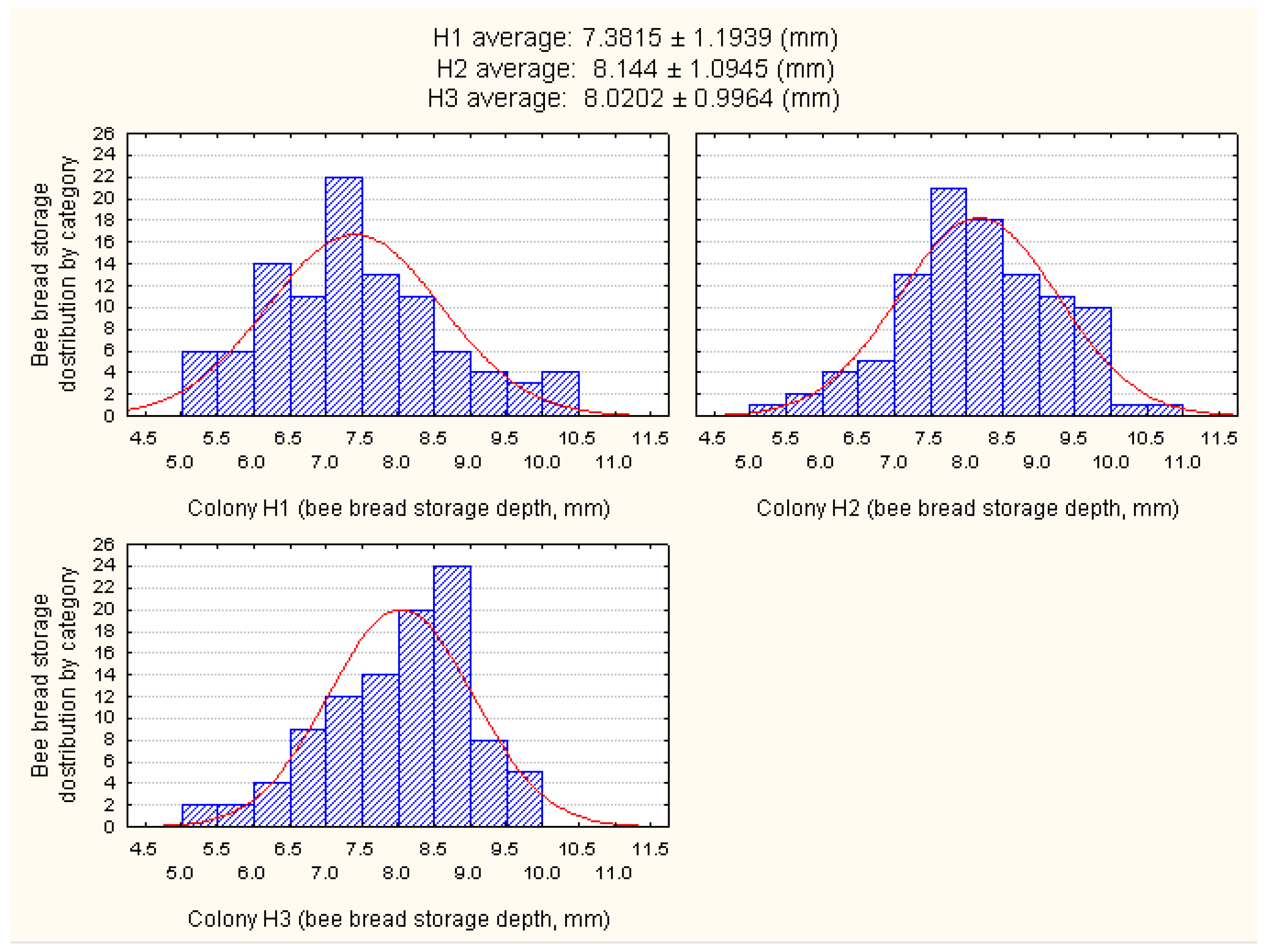
3.1. Determination of the Cell Depth of Bee Bread Storage
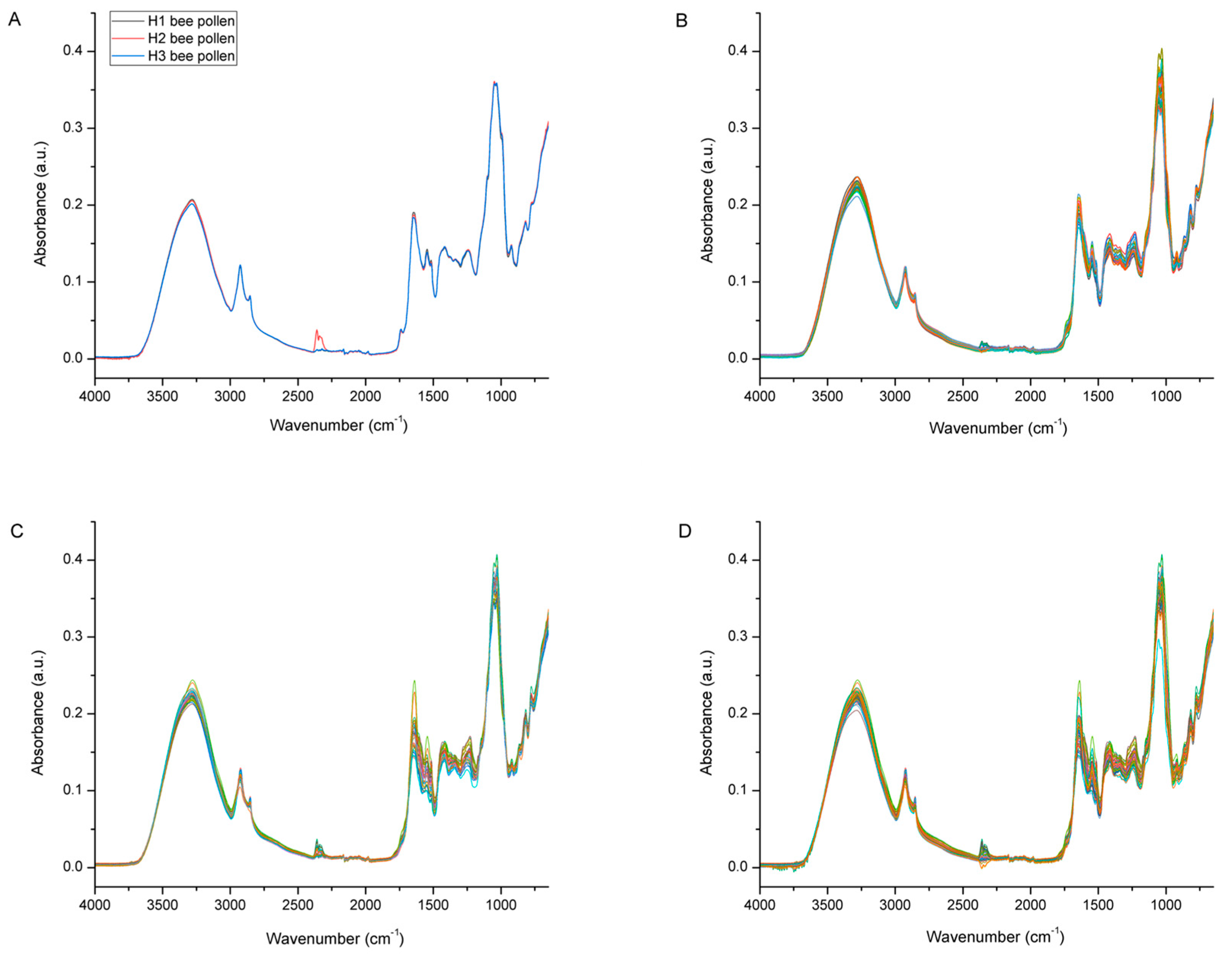
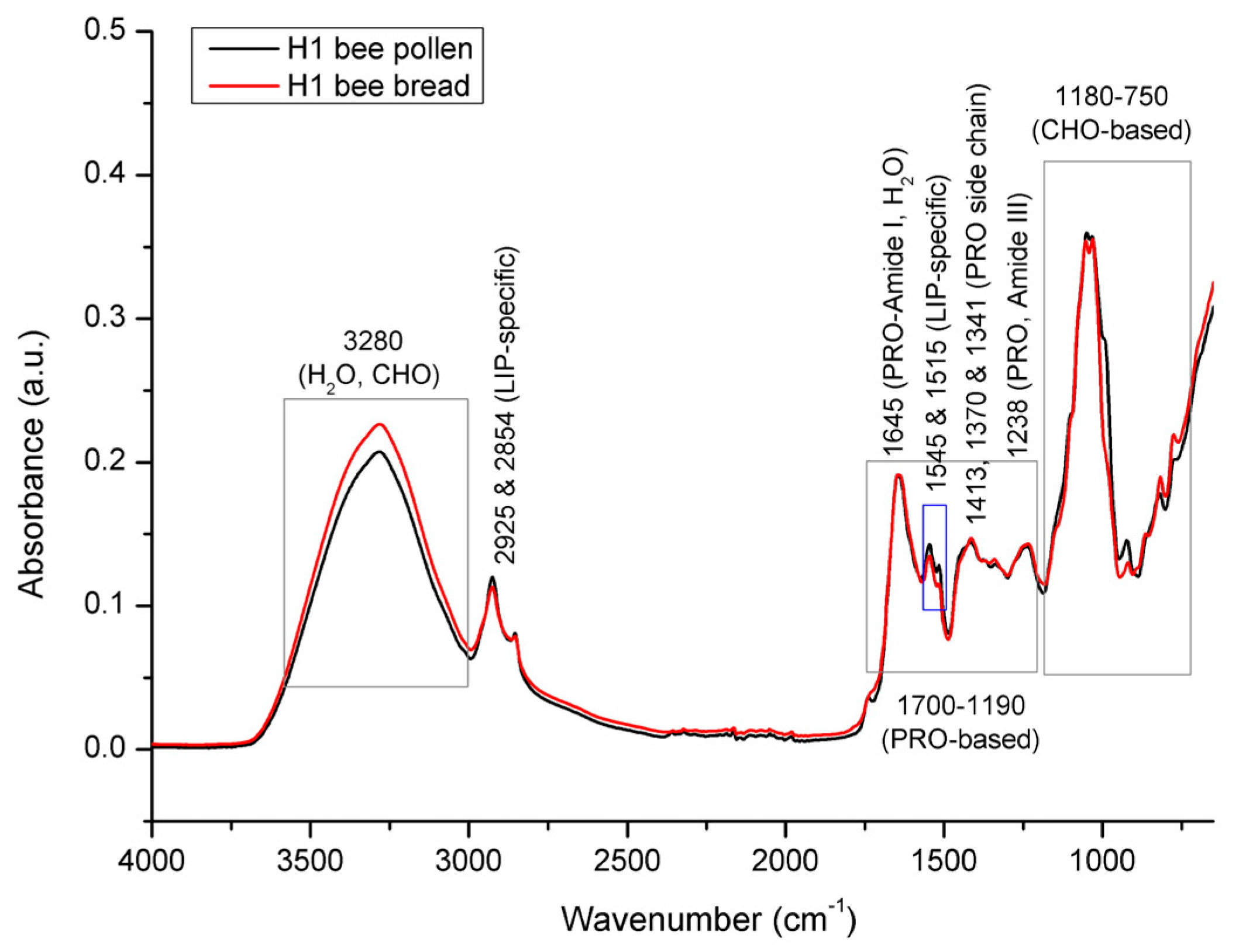
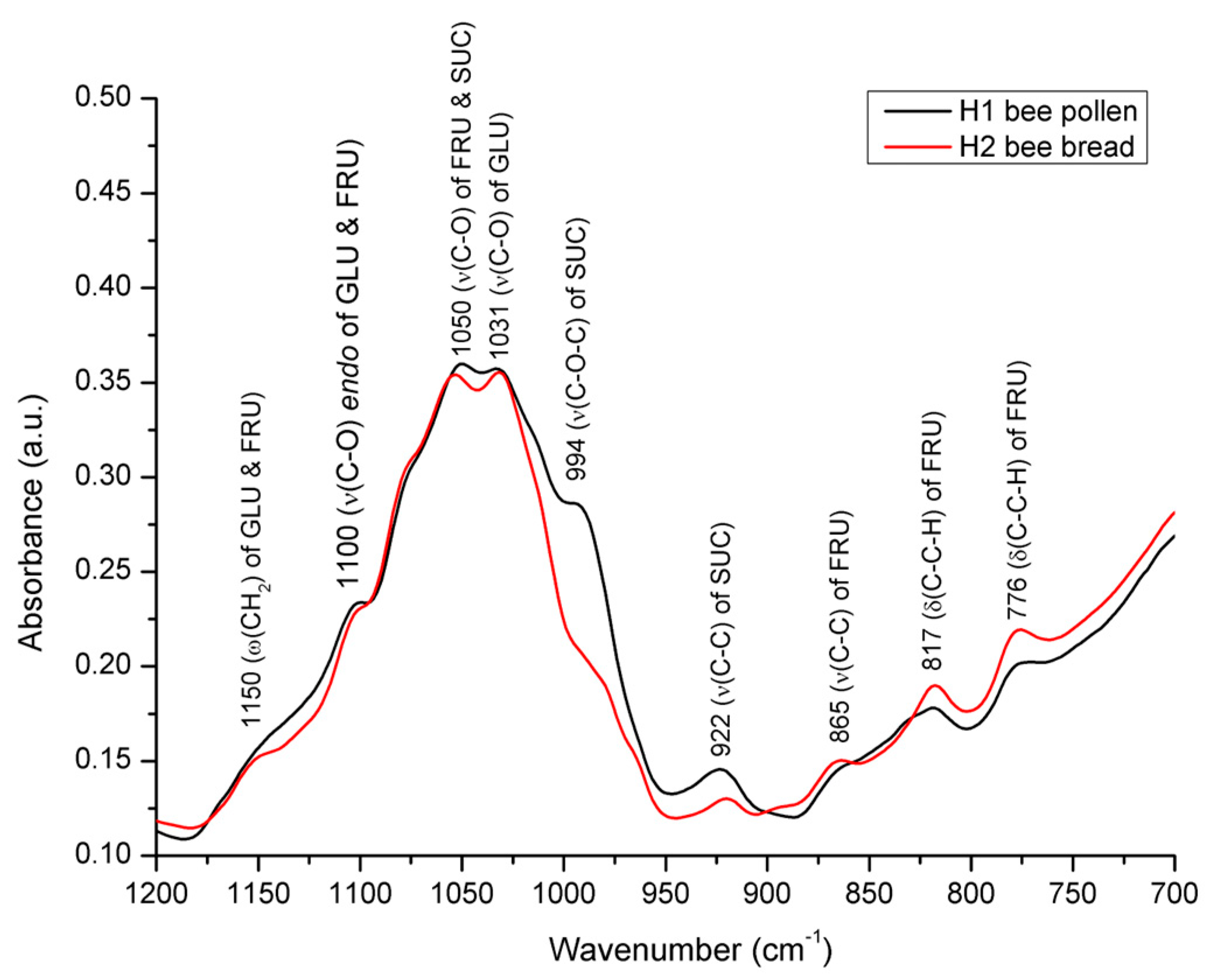
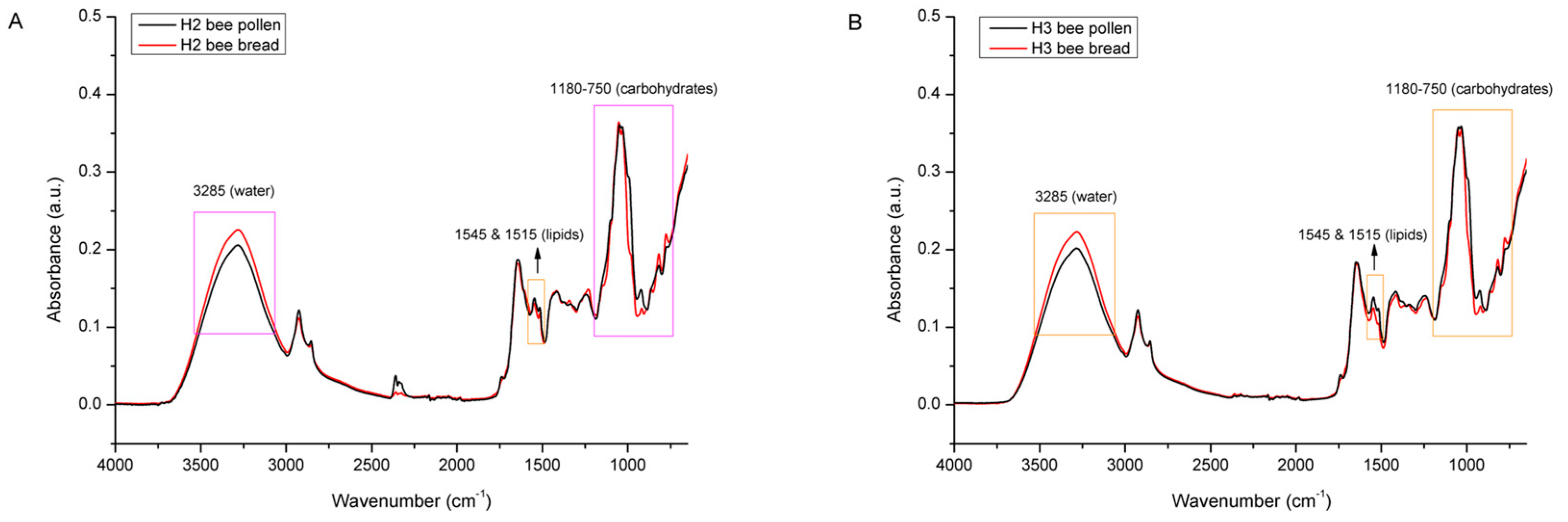
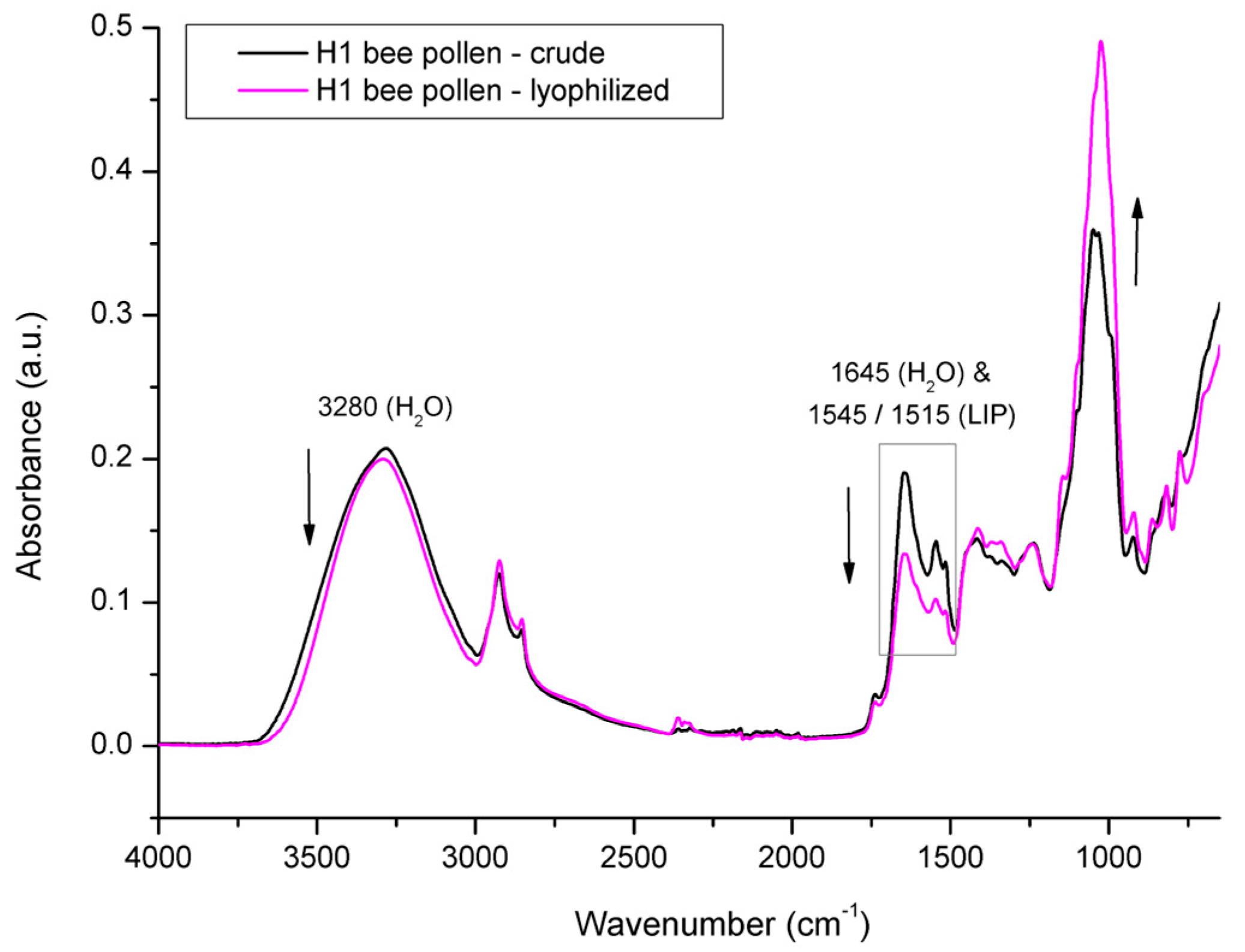
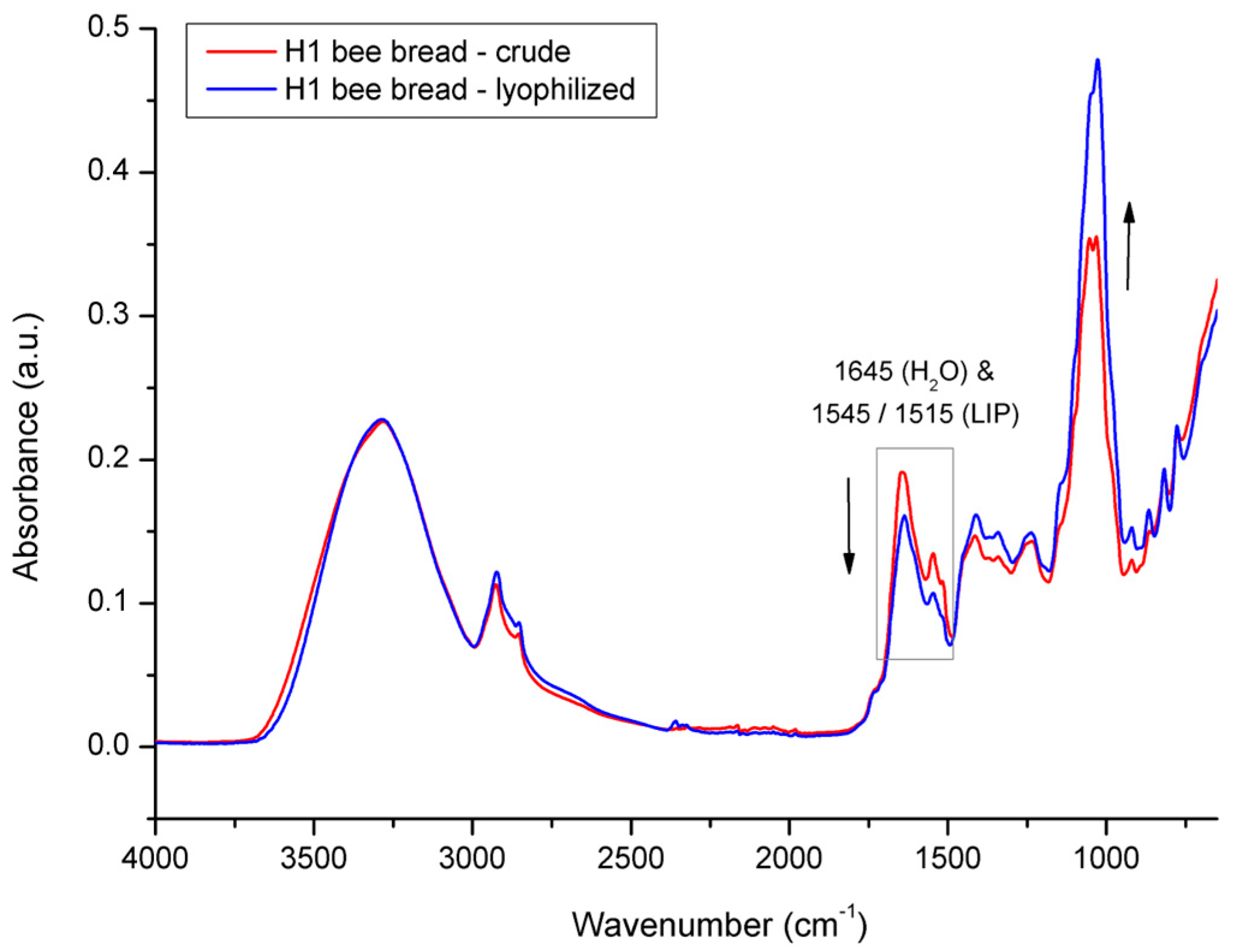
3.2. Spectral Analysis of Bee Pollen and Bee Bread
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Giampieri, F.; Quiles, J.L.; Cianciosi, D.; Forbes-Hernández, T.Y.; Orantes-Bermejo, F.J.; Alvarez-Suarez, J.M.; Battino, M. Bee Products: An Emblematic Example of Underutilized Sources of Bioactive Compounds. J. Agric. Food Chem. 2022, 70, 6833–6848. [Google Scholar] [CrossRef]
- Pełka, K.; Otłowska, O.; Worobo, R.W.; Szweda, P. Bee Bread Exhibits Higher Antimicrobial Potential Compared to Bee Pollen. Antibiotics 2021, 10, 125. [Google Scholar] [CrossRef]
- Khalifa, S.A.M.; Elashal, M.; Kieliszek, M.; Ghazala, N.E.; Farag, M.A.; Saeed, A.; Xiao, J.; Zou, X.; Khatib, A.; Göransson, U.; et al. Recent Insights into Chemical and Pharmacological Studies of Bee Bread. Trends Food Sci. Technol. 2020, 97, 300–316. [Google Scholar] [CrossRef]
- Baltrušaytyte, V.; Venskutonis, P.R.; Čeksteryte, V. Antibacterial Activity of Honey and Beebread of Different Origin against S. aureus and S. epidermidis. Food Technol. Biotechnol. 2007, 45, 201–208. [Google Scholar]
- Mărgăoan, R.; Cornea-Cipcigan, M.; Topal, E.; Kösoğlu, M. Impact of Fermentation Processes on the Bioactive Profile and Health-Promoting Properties of Bee Bread, Mead and Honey Vinegar. Processes 2020, 8, 1081. [Google Scholar] [CrossRef]
- Bakour, M.; Fernandes, Â.; Barros, L.; Sokovic, M.; Ferreira, I.C.F.R. Badiaa lyoussi Bee Bread as a Functional Product: Chemical Composition and Bioactive Properties. LWT-Food Sci. Technol. 2019, 109, 276–282. [Google Scholar] [CrossRef]
- Ivanišová, E.; Kačániová, M.; Frančáková, H.; Petrová, J.; Hutková, J.; Brovarskyi, V.; Velychko, S.; Adamchuk, L.; Schubertová, Z.; Musilová, J. Bee Bread—Perspective Source of Bioactive Compounds for Future. Potravinarstvo 2015, 9, 592–598. [Google Scholar] [CrossRef] [PubMed]
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Chlebowska-Śmigiel, A.; Wolska, I. Pollen and Bee Bread as New Health-Oriented Products: A Review. Trends Food Sci. Technol. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Eswaran, V.U.; Bhargava, H.R. Chemical Analysis and Anti-Microbial Activity of Karnataka Bee Bread of Apis Species. World Appl. Sci. J. 2014, 32, 379–385. [Google Scholar]
- Markiewicz-Żukowska, R.; Naliwajko, S.K.; Bortosiuk, E.; Moskwa, J.; Isidorov, V.; Soroczyńska, J.; Borawska, M.H. Chemical Composition and Antioxidant Activity of Beebread, and Its Influence on the Glioblastoma Cell Line (U87MG). J. Apic. Sci. 2013, 57, 147–157. [Google Scholar] [CrossRef]
- Abouda, Z.; Zerdani, I.; Kalalou, I.; Faid, M.; Ahami, M.T. The Antibacterial Activity of Moroccan Bee Bread and Bee-Pollen (Fresh and Dried) against Pathogenic Bacteria. Res. J. Microbiol. 2011, 6, 376–384. [Google Scholar] [CrossRef]
- Baltrušaityte, V.; Venskutonis, P.R.; Čeksteryte, V. Radical Scavenging Activity of Different Floral Origin Honey and Beebread Phenolic Extracts. Food Chem. 2007, 101, 502–514. [Google Scholar] [CrossRef]
- Herbert, E.W.; Shimanuki, H. Chemical Composition and Nutritive Value of Bee-Collected and Bee-Stored Pollen. Apidologie 1978, 9, 33–40. [Google Scholar] [CrossRef]
- Thakur, M.; Nanda, V. Composition and Functionality of Bee Pollen: A Review. Trends Food Sci. Technol. 2020, 98, 82–106. [Google Scholar] [CrossRef]
- Saavedra, K.I.; Rojas, C.; Delgado, G.E. Características Polínicas y Composición Química Del Polen Apícola Colectado En Cayaltí (Lambayeque-Perú). Rev. Chil. Nutr. 2013, 40, 71–78. [Google Scholar] [CrossRef]
- Prđun, S.; Svečnjak, L.; Valentić, M.; Marijanović, Z.; Jerković, I. Characterization of Bee Pollen: Physico-Chemical Properties, Headspace Composition and FTIR Spectral Profiles. Foods 2021, 10, 2103. [Google Scholar] [CrossRef]
- Castiglioni, S.; Astolfi, P.; Conti, C.; Monaci, E.; Stefano, M.; Carloni, P. Morphological, Physicochemical and FTIR Spectroscopic Properties of Bee Pollen Loads from Different Botanical Origin. Molecules 2019, 24, 3974. [Google Scholar] [CrossRef]
- Campos, M.G.R.; Bogdanov, S.; de Almeida-Muradian, L.B.; Szczesna, T.; Mancebo, Y.; Frigerio, C.; Ferreira, F. Pollen Composition and Standardisation of Analytical Methods. J. Apic. Res. 2008, 47, 154–161. [Google Scholar] [CrossRef]
- Sagona, S.; Pozzo, L.; Peiretti, P.G.; Biondi, C.; Giusti, M.; Gabriele, M.; Pucci, L.; Felicioli, A. Palynological Origin, Chemical Composition, Lipid Peroxidation and Fatty Acid Profile of Organic Tuscanian Bee-Pollen. J. Apic. Res. 2017, 56, 136–143. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Kaźmierczak, J.; Mencner, L.; Olczyk, K. Bee Pollen: Chemical Composition and Therapeutic Application. Evid.-Based Complement. Altern. Med. 2015, 2015, 297425. [Google Scholar] [CrossRef]
- Feás, X.; Vázquez-Tato, M.P.; Estevinho, L.; Seijas, J.A.; Iglesias, A. Organic Bee Pollen: Botanical Origin, Nutritional Value, Bioactive Compounds, Antioxidant Activity and Microbiological Quality. Molecules 2012, 17, 8359–8377. [Google Scholar] [CrossRef] [PubMed]
- Almeida-Muradian, L.B.; Pamplona, L.C.; Coimbra, S.; Barth, O.M. Chemical Composition and Botanical Evaluation of Dried Bee Pollen Pellets. J. Food Compos. Anal. 2005, 18, 105–111. [Google Scholar] [CrossRef]
- Mărgăoan, R.; Stranț, M.; Varadi, A.; Topal, E.; Yücel, B.; Cornea-Cipcigan, M.; Campos, M.G.; Vodnar, D.C. Bee Collected Pollen and Bee Bread: Bioactive Constituents and Health Benefits. Antioxidants 2019, 8, 568. [Google Scholar] [CrossRef]
- Zuluaga, C.M.; Serrato, J.C.; Quicazan, M.C. Chemical, Nutritional and Bioactive Characterization of Colombian Bee-Bread. Chem. Eng. Trans. 2015, 43, 175–180. [Google Scholar] [CrossRef]
- Smati, N. Bee Bread Preservation Methods: Physical, Chemical and Microbial Stability throughout Storage. Master’s Thesis, Instituto Politécnico de Bragança—Escola Superior Agrária, Bragança, Portugal, 2021. [Google Scholar]
- Disayathanoowat, T.; Li, H.; Supapimon, N.; Suwannarach, N.; Lumyong, S.; Chantawannakul, P.; Guo, J. Different Dynamics of Bacterial and Fungal Communities in Hive-Stored Bee Bread and Their Possible Roles: A Case Study from Two Commercial Honey Bees in China. Microorganisms 2020, 8, 264. [Google Scholar] [CrossRef]
- Urcan, A.C.; Mărghitaş, L.A.; Dezmirean, D.S.; Bobiş, O.; Bonta, V.; Mureşan, C.I.; Mărgăoan, R. Chemical Composition and Biological Activities of Beebread—Review. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca Anim. Sci. Biotechnol. 2017, 74, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Mayda, N.; Özkök, A.; Bayram, N.E.; Gerçek, Y.C.; Sorkun, K. Bee Bread and Bee Pollen of Different Plant Sources: Determination of Phenolic Content, Antioxidant Activity, Fatty Acid and Element Profiles. J. Food Meas. Charact. 2020, 14, 1795–1809. [Google Scholar] [CrossRef]
- Bogdanov, S. Pollen: Nutrition, functional properties, health: A review. Bee Prod. Sci. 2011, 5, 20–350. [Google Scholar]
- DeGrandi-Hoffman, G.; Eckholm, B.J.; Huang, M.H. A Comparison of Bee Bread Made by Africanized and European Honey Bees (Apis mellifera) and Its Effects on Hemolymph Protein Titers. Apidologie 2013, 44, 52–63. [Google Scholar] [CrossRef]
- Bader, C.; Costa, J.; Lee, N.; Smith, R.; Ri, R.; Weaver, J.C.; Oxman, N. Computational Methods for the Characterization of Apis mellifera Comb Architecture. Commun. Biol. 2022, 5, 468. [Google Scholar] [CrossRef]
- Barene, I.; Daberte, I.; Siksna, S. Investigation of Bee Bread and Development of Its Dosage Forms. Med. Teor. Ir Prakt. 2015, 21, 16–22. [Google Scholar] [CrossRef]
- Quinby, M. Mysteries of Bee-Keeping Explained; Saxton, C.M., Ed.; Agricultural Book Publisher: New York, NY, USA, 2008. [Google Scholar]
- Delaplane, K.S.; Van Der Steen, J.; Guzman-Novoa, E. Standard Methods for Estimating Strength Parameters of Apis mellifera Colonies. J. Apic. Res. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Svečnjak, L.; Chesson, L.A.; Gallina, A.; Maia, M.; Martinello, M.; Mutinelli, F.; Muz, M.N.; Nunes, F.M.; Saucy, F.; Tipple, B.J.; et al. Standard Methods for Apis mellifera Beeswax Research. J. Apic. Res. 2019, 58, 1–108. [Google Scholar] [CrossRef]
- Statistica. Data Analysis Software System, Version 7; StatSoft Inc.: Tulsa, OK, USA, 2004. [Google Scholar]
- Resolutions Pro. FTIR Software, Version 5.3.0; Agilent Technologies: Palo Alto, CA, USA, 2015. [Google Scholar]
- Origin. Data Analysis and Graphing Software, Version 8.1; OriginLab Corporation: Northampton, MA, USA, 2009. [Google Scholar]
- Seeley, T.D.; Morse, R.A. The Nest of the Honey Bee (Apis mellifera L.). Insectes Soc. 1976, 23, 495–512. [Google Scholar] [CrossRef]
- Al-Kahtani, S.N.; Taha, E.-K.A. Effect of Comb Age on Cell Measurements and Worker Body Size. PLoS ONE 2021, 16, e0260865. [Google Scholar] [CrossRef]
- McMullan, J.B.; Brown, M.J.F. The Influence of Small-Cell Brood Combs on the Morphometry of Honeybees (Apis mellifera). Apidologie 2006, 37, 665–672. [Google Scholar] [CrossRef]
- Yang, S.; Deng, S.; Kuang, H.; Zhou, D.; Gong, X.; Dong, K. Evaluating and Comparing the Natural Cell Structure and Dimensions of Honey Bee Comb Cells of Chinese Bee, Apis cerana cerana (Hymenoptera: Apidae) and Italian Bee, Apis mellifera ligustica (Hymenoptera: Apidae). J. Insect Sci. 2021, 21, 1. [Google Scholar] [CrossRef]
- Faji, M.; Abebe, A.; Mijena, D.; Begna, F.; Tarekegn, A. Determination of Bee Spacing and Comb Cell Dimensions for Apis mellifera scutellata Honeybee Race in Western Ethiopia. Int. J. Livest. Prod. 2018, 9, 206–210. [Google Scholar] [CrossRef]
- Carroll, M.J.; Brown, N.; Goodall, C.; Downs, A.M.; Sheenan, T.H.; Anderson, K.E. Correction: Honey Bees Preferentially Consume Freshly-Stored Pollen. PLoS ONE 2021, 16, e0249458. [Google Scholar] [CrossRef]
- Anjos, O.; Santos, A.J.A.; Dias, T.; Estevinho, L.M. Application of FTIR-ATR Spectroscopy on the Bee Pollen Characterization. J. Apic. Res. 2017, 56, 210–218. [Google Scholar] [CrossRef]
- Isopescu, R.D.; Spulber, R.; Josceanu, A.M.; Mihaiescu, D.E.; Popa, O. Romanian Bee Pollen Classification and Property Modelling. J. Apic. Res. 2020, 59, 443–451. [Google Scholar] [CrossRef]
- Kasprzyk, I.; Depciuch, J.; Grabek-Lejko, D.; Parlinska-Wojtan, M. FTIR-ATR Spectroscopy of Pollen and Honey as a Tool for Unifloral Honey Authentication. The Case Study of Rape Honey. Food Control 2018, 84, 33–40. [Google Scholar] [CrossRef]
- Dranca, F.; Ursachi, F.; Oroian, M. Bee Bread: Physicochemical Characterization and Phenolic Content Extraction Optimization. Foods 2020, 9, 1358. [Google Scholar] [CrossRef]
- Max, J.-J.; Chapados, C. Glucose and Fructose Hydrates in Aqueous Solution by IR Spectroscopy. J. Phys. Chem. A 2007, 111, 2679–2689. [Google Scholar] [CrossRef] [PubMed]
- Max, J.J.; Chapados, C. Sucrose Hydrates in Aqueous Solution by IR Spectroscopy. J. Phys. Chem. A 2001, 105, 10681–10688. [Google Scholar] [CrossRef]
- Socrates, G. Infrared and Raman Characteristic Group Frequencies. Tables and Charts, 3rd ed.; John Wiley and Sons, Ltd.: New York, NY, USA, 2004. [Google Scholar]
- Kong, J.; Yu, S. Fourier Transform Infrared Spectroscopic Analysis of Protein Secondary Structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [CrossRef]
- Conte, G.; Benelli, G.; Serra, A.; Signorini, F.; Bientinesi, M.; Nicolella, C.; Mele, M.; Canale, A. Lipid Characterization of Chestnut and Willow Honeybee-Collected Pollen: Impact of Freeze-Drying and Microwave-Assisted Drying. J. Food Compos. Anal. 2017, 55, 12–19. [Google Scholar] [CrossRef]
- Laurens, L.M.L.; Wolfrum, E.J. Feasibility of Spectroscopic Characterization of Algal Lipids: Chemometric Correlation of NIR and FTIR Spectra with Exogenous Lipids in Algal Biomass. Bioenergy Res. 2011, 4, 22–35. [Google Scholar] [CrossRef]
- Pielichowska, K.; Głowinkowski, S.; Lekki, J.; Biniaś, D.; Pielichowski, K.; Jenczyk, J. PEO/Fatty Acid Blends for Thermal Energy Storage Materials. Structural/Morphological Features and Hydrogen Interactions. Eur. Polym. J. 2008, 44, 3344–3360. [Google Scholar] [CrossRef]
- Chandra, P.; Enespa; Singh, R.; Arora, P.K. Microbial Lipases and Their Industrial Applications: A Comprehensive Review. Microb. Cell Factories 2020, 19, 169. [Google Scholar] [CrossRef]
- Ciafardini, G.; Zullo, B.A.; Iride, A. Lipase Production by Yeasts from Extra Virgin Olive Oil. Food Microbiol. 2006, 23, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.B.; Katz, M.B.; González, S.; Oliver, G. Determination of Esterolytic and Lipolytic Activities of Lactic Acid Bacteria. In Public Health Microbiology. Methods in Molecular Biology; Spencer, J.F.T., Ragout de Spencer, A.L., Eds.; Humana Press: Totowa, NJ, USA, 2004; Volume 268, pp. 465–470. [Google Scholar]
- Wang, J.; Kliks, M.M.; Jun, S.; Jackson, M.; Li, Q.X. Rapid Analysis of Glucose, Fructose, Sucrose, and Maltose in Honeys from Different Geographic Regions Using Fourier Transform Infrared Spectroscopy and Multivariate Analysis. J. Food Sci. 2010, 75, C208–C214. [Google Scholar] [CrossRef]
- Kačuráková, M.; Mathlouthi, M. FTIR and Laser-Raman Spectra of Oligosaccharides in Water: Characterization of the Glycosidic Bond. Carbohydr. Res. 1996, 284, 145–157. [Google Scholar] [CrossRef] [PubMed]
- Svečnjak, L.; Prđun, S.; Rogina, J.; Bubalo, D.; Jerković, I. Characterization of Satsuma Mandarin (Citrus unshiu Marc.) Nectar-to-Honey Transformation Pathway Using FTIR-ATR Spectroscopy. Food Chem. 2017, 232, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Viesser, J.A.; de Melo Pereira, G.V.; de Carvalho Neto, D.P.; Rogez, H.; Góes-Neto, A.; Azevedo, V.; Brenig, B.; Aburjaile, F.; Soccol, C.R. Co-Culturing Fructophilic Lactic Acid Bacteria and Yeast Enhanced Sugar Metabolism and Aroma Formation during Cocoa Beans Fermentation. Int. J. Food Microbiol. 2021, 339, 109015. [Google Scholar] [CrossRef] [PubMed]
- Castagna, A.; Benelli, G.; Conte, G.; Sgherri, C.; Signorini, F.; Nicolella, C.; Ranieri, A.; Canale, A. Drying Techniques and Storage: Do They Affect the Nutritional Value of Bee-Collected Pollen? Molecules 2020, 25, 4925. [Google Scholar] [CrossRef]
- Keskin, M.; Özkök, A. Effects of Drying Techniques on Chemical Composition and Volatile Constituents of Bee Pollen. Czech J. Food Sci. 2020, 38, 203–208. [Google Scholar] [CrossRef]
- de Arruda, V.A.S.; Pereira, A.A.S.; de Freitas, A.S.; Barth, O.M.; de Almeida-Muradian, L.B. Dried Bee Pollen: B Complex Vitamins, Physicochemical and Botanical Composition. J. Food Compos. Anal. 2013, 29, 100–105. [Google Scholar] [CrossRef]
- Hazarika, U.; Kovács, Z.; Bodor, Z.; Gosztola, B. Phytochemicals and Organoleptic Properties of French Tarragon (Artemisia dracunculus L.) Influenced by Different Preservation Methods. LWT-Food Sci. Technol. 2022, 169, 114006. [Google Scholar] [CrossRef]
- Pena-Rodríguez, E.; Mata-Ventosa, A.; Garcia-Vega, L.; Pérez-Torras, S.; Fernández-Campos, F. The Physicochemical, Biopharmaceutical, and In Vitro Efficacy Properties of Freeze-Dried Dexamethasone-Loaded Lipomers. Pharmaceutics 2021, 13, 1322. [Google Scholar] [CrossRef]
- Franzé, S.; Selmin, F.; Samaritani, E.; Minghetti, P.; Cilurzo, F. Lyophilization of Liposomal Formulations: Still Necessary, Still Challenging. Pharmaceutics 2018, 10, 139. [Google Scholar] [CrossRef] [PubMed]
- Ryckebosch, E.; Muylaert, K.; Eeckhout, M.; Ruyssen, T.; Foubert, I. Influence of Drying and Storage on Lipid and Carotenoid Stability of the Microalga Phaeodactylum tricornutum. J. Agric. Food Chem. 2011, 59, 11063–11069. [Google Scholar] [CrossRef] [PubMed]
- Silva, S.D.; Rosa, N.F.; Ferreira, A.E.; Boas, L.V.; Bronze, M.R. Rapid Determination of α-Tocopherol in Vegetable Oils by Fourier Transform Infrared Spectroscopy. Food Anal. Methods 2009, 2, 120–127. [Google Scholar] [CrossRef]
- Dahl, Å. Pollen Lipids Can Play a Role in Allergic Airway Inflammation. Front. Immunol. 2018, 9, 2816. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Depth of Honeycomb Cell (mm) | Experimental Colony | |||
|---|---|---|---|---|
| H1 | H2 | H3 | Total | |
| Average | 10.99 | 11.01 | 11.01 | 11.00 |
| Minimum | 9.71 | 8.97 | 9.37 | 9.35 |
| Maximum | 12.05 | 11.98 | 12.36 | 12.13 |
| Standard deviation | 0.57 | 0.81 | 0.79 | 0.72 |
| Depth of the Cell in Which Bees Store Bee Bread (mm) | Experimental Colony | |||
|---|---|---|---|---|
| H1 | H2 | H3 | Total | |
| Average | 7.38 | 8.14 | 8.02 | 7.85 |
| Minimum | 5.19 | 5.25 | 5.20 | 5.21 |
| Maximum | 10.26 | 10.71 | 9.72 | 10.23 |
| Standard deviation | 1.19 | 1.09 | 1.00 | 1.09 |
| Side Length of the Honeycomb Cell (mm) | Experimental Colony | |||
|---|---|---|---|---|
| H1 | H2 | H3 | Total | |
| Average | 9.73 | 9.63 | 9.87 | 9.74 |
| Minimum | 9.12 | 9.12 | 9.11 | 9.12 |
| Maximum | 10.47 | 10.66 | 10.76 | 10.63 |
| Standard deviation | 0.39 | 0.35 | 0.48 | 0.41 |
| Angle (∡) of Hexagonal Honeycomb Cell (°) | Experimental Colony | |||
|---|---|---|---|---|
| H1 | H2 | H3 | Total | |
| Average | 118.5 | 118.4 | 118.4 | 118.4 |
| Minimum | 117.8 | 117.8 | 117.7 | 117.8 |
| Maximum | 119.1 | 119.2 | 119.4 | 119.2 |
| Standard deviation | 0.37 | 0.39 | 0.41 | 0.39 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Svečnjak, L.; Bošković, K.; Prđun, S.; Mrkonjić Fuka, M.; Tanuwidjaja, I. Metric and Spectral Insight into Bee-Pollen-to-Bee-Bread Transformation Process. Foods 2023, 12, 4149. https://doi.org/10.3390/foods12224149
Svečnjak L, Bošković K, Prđun S, Mrkonjić Fuka M, Tanuwidjaja I. Metric and Spectral Insight into Bee-Pollen-to-Bee-Bread Transformation Process. Foods. 2023; 12(22):4149. https://doi.org/10.3390/foods12224149
Chicago/Turabian StyleSvečnjak, Lidija, Kristian Bošković, Saša Prđun, Mirna Mrkonjić Fuka, and Irina Tanuwidjaja. 2023. "Metric and Spectral Insight into Bee-Pollen-to-Bee-Bread Transformation Process" Foods 12, no. 22: 4149. https://doi.org/10.3390/foods12224149
APA StyleSvečnjak, L., Bošković, K., Prđun, S., Mrkonjić Fuka, M., & Tanuwidjaja, I. (2023). Metric and Spectral Insight into Bee-Pollen-to-Bee-Bread Transformation Process. Foods, 12(22), 4149. https://doi.org/10.3390/foods12224149